







-
林分生物量是森林生态系统对有机质所积累的质量,是森林生态系统最基本的特征数据,对研究森林生态系统物质循环、能量流动具有重要的作用[1-3]。林分生物量的动态变化不仅能够反映物质与能量在生态系统中变化的复杂关系,同时在一定程度上能够反映生态系统的健康水平[4-5]。生物量的积累表现为林分生长。立地条件与林分生长密切相关,好的立地条件为林分生长提供充足的养分,加快林分生长[6]。林分结构和树种多样性也会影响林分有机质的积累速度。岩溶地区高温多雨且分布不均,基岩裸露率高、土壤瘠薄、季节性岩溶干旱明显[7-8]。岩溶区的适生植被不仅需具有石生性、喜钙性、耐干旱和耐贫瘠的特点,而且需具有发达的根系。植被的恢复和重建是岩溶退化区的重点和难点[9],生物量作为生态系统生产力的基础和功能的表现形式,是退化区植被恢复成功与否的评价指标[10]。目前对岩溶生态系统植被生物量的研究集中在年龄序列、演替阶段等单一因素对生物量的影响[11-13]。然而,生态系统中的影响是一个复杂的过程,由很多相互的作用组成[14-15],因此综合探讨林分生物量与多种影响因素之间的关系具有必要性和现实性。传统的路径分析及主成分分析不能同时处理多个因变量与自变量的联系,不能提供完整的信息[16]。结构方程模型是一种多变量的统计方法,能够同时研究自然系统中多个因子间的相互关系[6, 17]。相较于传统的相关性分析,结构方程模型是基于研究者的先验知识预先设定系统内因子间的依赖关系,能够判别各因子间的关系强度,同时还能作出整体拟合和判断,反映自变量与因变量之间的直接和间接关系,帮助研究者更全面掌握自然系统中的真实机制[6, 18]。近年来结构方程模型在生态学和林学上得到广泛应用[19-20]。河南省淅川县是南北过渡带和岩溶石漠化集连片区的最北端,岩溶面积达348.82 km2,又是南水北调中线工程核心水源地、主要淹没区及渠首所在地,地理位置特殊[21]。调查发现:岩溶区林分质量明显低于非岩溶区,生态资源环境问题严重,影响着南水北调水中线安全。提高该区域生态环境,保障南水北调中线安全势在必行,林分质量提升已经迫在眉睫。本研究以淅川县岩溶区天然次生林为研究对象,利用结构方程模型,探讨岩溶区建群种生物量、林分总生物量与基岩裸露度、土壤厚度以及林分密度和物种多样性之间的耦合关系,明确各关系之间影响强度,以期为河南省淅川县岩溶区植被的经营和管理提供理论依据,以期改善淅川县岩溶区生态环境。
HTML
-
淅川县位于河南省西南部,豫、鄂、陕三省结合处,32°55′~33°23′N, 110°58′~111°53′E,地处伏牛山余脉,拥有亚洲最大的淡水湖,属于南水北调中线工程水源地所在地。淅川县气候适宜,属于北亚热带向暖温带过渡的季风性气候区,四季分明。年平均降水量为804.3 mm,6−9月降水占全年降水量的58.0%以上,最高达79.8%。年平均气温为15.8 ℃,无霜期约228 d。岩溶地貌在淅川分布广泛,主要集中在丹江口库区周边和荆关、西簧、金河、大石桥等12个乡镇,岩溶区现有植被以次生林为主。其中主要的乔木树种为栓皮栎Quercus variabilis、黄连木Pistacia chinensis、化香Platycarya strobilacea、侧柏Platycladus orientalis、女贞Ligustrum lucidum等;主要的灌木树种为小果蔷薇Rosa cymosa、菝葜Smilax china、黄荆Vitex negundo、荚蒾Viburnum dilatatum、连翘Hypericum ascyron等;主要的草本为碎米莎草Cyperus iria、绶草Spiranthes sinensis、天门冬Asparagus cochinchinensis、荩草Arthraxon hispidus等。
-
2018年7−9月分别在淅川县金河镇、西簧乡、荆关镇岩溶区选择6个样点18个20 m×30 m样地,在每个样地中划分6个10 m×10 m小样方,共108个样方进行数据采集。在每块样地进行每木检尺,确定乔木层树种组成,测定树高、胸径、枝下高和冠幅。同时测量每块样地的坡度、坡向、海拔、基岩裸露度、土壤厚度,测定土壤含水率和土壤容重等。
采用中国森林生态系统碳储量—生物量方程[22]计算林分生物量和建群种生物量。
采用Shannon-Wiener指数计算各样地树种多样性[23]:
式(1)中:H为树种多样性;B为林分胸径面积总和;Bi为树种i的胸径断面积总和;N为树种数。结果见表1。
样地
编号纬度(N) 经度(E) 坡度/
(°)海拔/
m土壤厚度/cm 基岩裸露度/% 含水率/% 平均胸径/cm 平均树高/m 优势木生物量/
(t·hm−2)林分总生物量/
(t·hm−2)主要树种 种群密度/
(株·hm−2)P1 33°15′57.94″ 111°14′35.31″ 21 333 35.0 70 13.7 11.8 9.8 97.71 119.18 栓皮栎、马尾松、
山槐1 750 P2 33°15′57.80″ 111°14′34.67″ 21 255 15.0 55 13.6 8.0 8.6 61.50 79.35 栓皮栎、马尾松、
蒙桑1 550 P3 33°15′57.01″ 111°14′34.39″ 29 249 15.0 50 20.1 9.9 10.7 106.82 124.15 栓皮栎、山胡椒 1 567 P4 33°07′26.76″ 111°26′01.23″ 24 311 22.0 75 18.9 13.0 13.0 144.02 158.23 栓皮栎、黄连木、
侧柏1 933 P5 33°07′26.44″ 111°26′01.60″ 24 322 18.0 70 17.0 11.9 13.0 130.15 147.98 栓皮栎、黄连木、
侧柏2 000 P6 33°07′26.73″ 111°26′02.10″ 27 345 27.0 75 19.2 11.0 12.5 166.94 191.28 栓皮栎、黄连木、
侧柏1 883 P7 33°07′49.72″ 111°25′22.30″ 24 219 20.0 65 16.9 12.9 15.4 112.21 143.61 栓皮栎、黄连木、
女贞1 767 P8 33°07′50.85″ 111°25′22.60″ 24 239 28.0 50 17.0 14.3 17.5 224.42 239.15 栓皮栎、黄连木、
蒙桑1 950 P9 33°07′51.15″ 111°25′22.76″ 21 250 22.0 52 15.7 14.0 14.6 148.92 169.33 栓皮栎、黄连木、
鸡仔木1 400 P10 33°07′26.65″ 111°25′57.23″ 20 279 28.0 80 17.6 10.5 13.0 120.29 139.66 栓皮栎、黄连木、
侧柏1 833 P11 33°07′26.86″ 111°25′57.00″ 21 297 25.0 78 16.4 12.0 10.4 117.67 139.44 栓皮栎、黄连木 1 667 P12 33°07′27.10″ 111°25′56.75″ 20 308 26.0 80 18.3 13.3 16.1 172.21 182.27 栓皮栎、黄连木 2 700 P13 33°07′49.36″ 111°26′11.57″ 35 470 17.0 10 10.1 11.7 11.7 165.95 176.74 栓皮栎、化香、
黄连木1 983 P14 33°07′50.35″ 111°26′13.63″ 30 472 18.0 12 6.0 11.0 11.8 138.31 154.57 栓皮栎、黄连木、
侧柏1 650 P15 33°07′49.01″ 111°25′22.02″ 30 450 15.0 20 4.3 11.2 12.0 142.95 147.96 栓皮栎、黄连木、
侧柏2 483 P16 33°16′37.23″ 111°03′06.28″ 35 651 12.4 20 10.6 10.8 12.7 296.72 312.55 栓皮栎、飞蛾槭 2 867 P17 33°16′35.60″ 111°03′05.38″ 35 627 18.8 10 12.9 14.0 14.8 280.25 288.55 栓皮栎、侧柏 1 800 P18 33°16′34.65″ 111°03′18.91″ 32 574 22.0 40 14.0 21.0 19.7 190.55 234.44 栓皮栎、槲栎 767 说明:栓皮栎Quercus variabilis、马尾松Pinus massoniana、山槐Maackia hupehensis、蒙桑Morus mongolica、山胡椒Lindera glauca、 侧柏Platycladus orientalis、女贞Ligustrum lucidum、鸡仔木Sinoadina racemosa、飞蛾槭Acer oblongum、槲栎Quercus aliena Table 1. Sample information
-
结构方程模型是一种建立、估计和检验因果关系的模型,结合了因素分析和路径分析2种统计方法,并且同时检验模型中的显性变量、潜在变量、干扰或误差变量间的关系。观测变量是直接测得的变量、潜在变量是不可直接测得的变量,但是可由观测变量表示。如基岩裸露度、土壤厚度、海拔等为可以直接测量的指标,这些指标能够反映不能够直接测量的指标立地条件(潜在变量),本研究中采用能够直接测量的指标构建结构方程模型。
结构方程模型由测量模型和结构模型两部分组成。测量模型是潜在变量与显性变量之间关系的模型。
式(2)~(3)中:X为外生显性变量向量;Y为内生显性变量向量;Λx和Λy为指标变量(X, Y)的因素负荷量;δ、ε为外生显性变量和内生显性变量的测量误差;ξ为外生潜在变量;η为内生潜在变量[24]。
立地条件决定林分的生长状况,直接影响林分的结构和树木的多样性以及林分的生物量,立地条件包含地形因子、土壤因子以及气候因子[18],森林的功能与林分结构密切相关,林分结构表征了林木的分布与生长状况。林分多样性影响树种间的差异,并且影响林分的生长[25]。由于岩溶区基岩裸露度高,土壤瘠薄且不连续,对岩溶区植被有着最直接的影响。故本研究中将基岩裸露度(x1)、土壤厚度(x2)、树种多样性(x3)、林分密度(x4)等观测变量作为探讨与林分生物量关系的显变量,对应误差为δ1、δ2、δ3、δ4。建群种生物量(η1)和林分总生物量(η2)作为与内生潜在变量(生物量η)有关的观测变量,ε1、ε2为对应的误差,构建各显变量与内生潜变量(生物量η)结构方程模型。运用SPSS 18.0软件对数据进行处理,运用Amos 23.0软件构建结构方程模型进行林分生长与影响因子间关系耦合。
1.1. 研究地区概况
1.2. 调查方法与数据处理
1.3. 模型构建原理
-
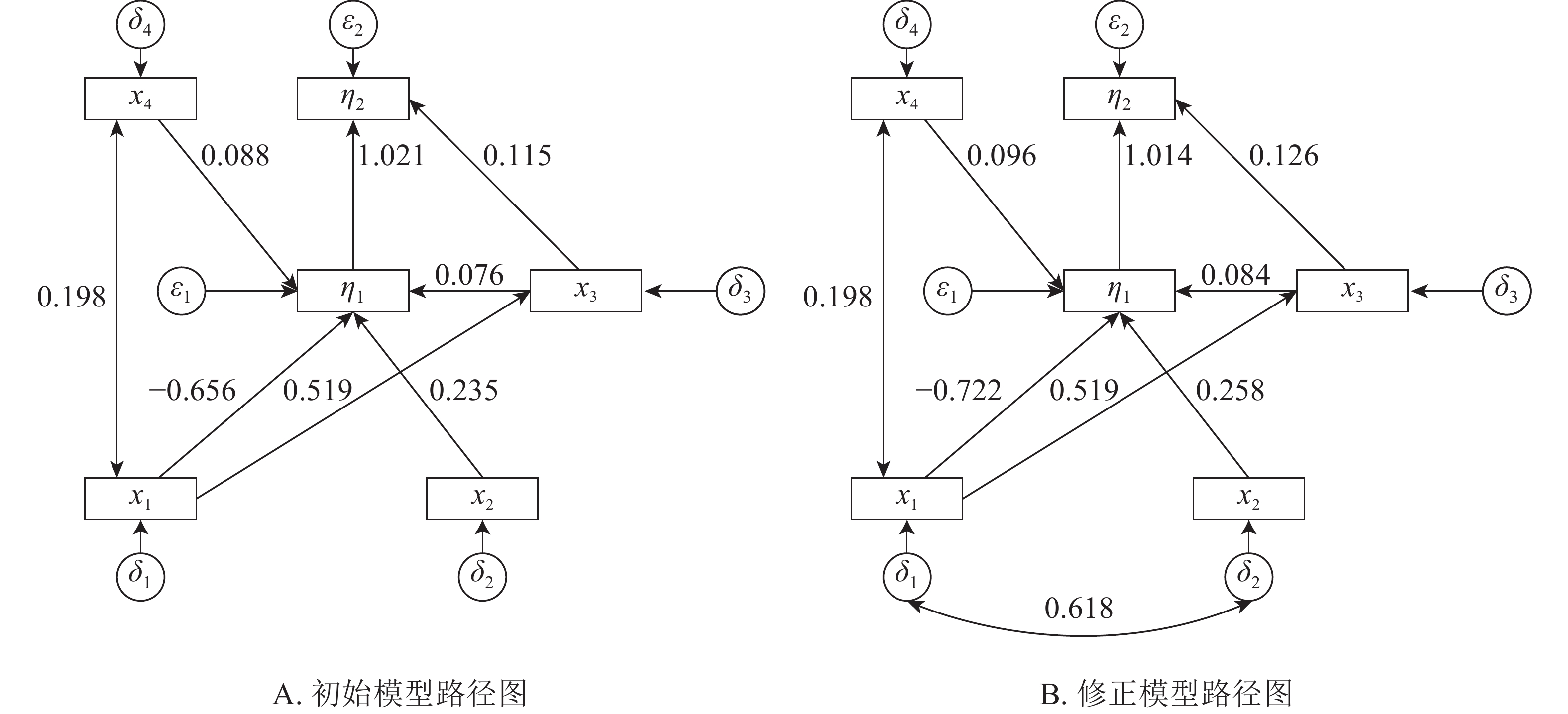
本研究建立基岩裸露度(x1)、土壤厚度(x2)、树种多样性(x3)、林分密度(x4)等观测变量与建群种生物量和林分生物量耦合关系的结构方程模型。根据拟合结果,逐步的调整路径关系,删除关系不显著(P>0.05)的路径,得到初始结构方程模型1(图1A)。初始模型χ2=13.236,df=7,χ2/df=1.891<3,显著性概率值P=0.067>0.05,接受虚无假设。但模型适配度指数IGF=0.829<0.90,模型规准适配指数(INF)、相对适配指数(IRF)、非规准适配指数(ITL)均较小(表2),模型拟合结果需进一步进行模型修正。修正后得到最优模型2(图1B)。
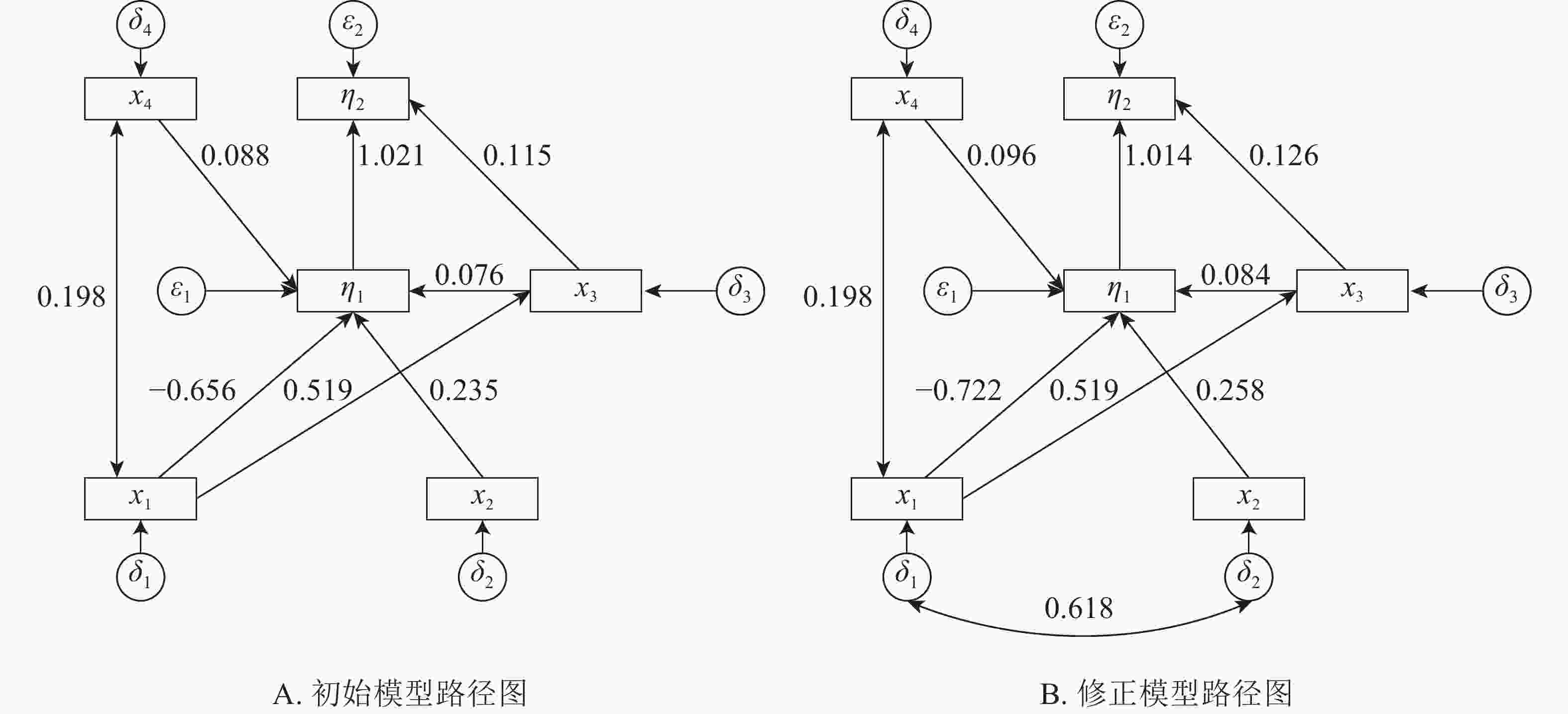
Figure 1. Initial and correction model path map
统计量 参数名称 参数含义 评价标准 模型1 模型2 绝对适配统计量 c2/df 卡方自由度比 <3 1.890 0.842 IGF 适配度指数 >0.90 0.829 0.914 增值适配度统计量 INF 规准适配指数 介于0~1之间(可以>1),越接近1,表示模型适配度越好 0.882 0.955 IRF 相对适配指数 0.747 0.887 IIF 增值适配指数 0.941 1.009 ITL 非规准适配指数 0.862 1.024 IGF 比较适配指数 0.936 1.000 简约适配统计量 PNC 非中心参数 越小越好 6.236 0 Table 2. Fitting parameters of structure equation model for the growth of forest
最优模型χ2=5.051,df=6, χ2/df=0.842<3,显著性概率值P=0.537>0.05,接受虚无假设,观测数据所导出的方差协方差矩阵与假设模型导出的方差协方差矩阵适配的假设获得支持,假设模型与观测数据适配。模型中各项检验指标均达到标准,如表2。同时渐进残差均方和平方根RMESA=0<0.08、以及赤池信息量AIC(预设模型)=35.051<42.000(饱和模型)<123.965(独立模型),假设模型与数据适配良好。
-
由表3所示:基岩裸露度对林分密度、树种多样性有正影响,对建群种生物量有负影响,其路径系数分别为0.198、0.519、−0.722,即每当基岩裸露度变化1时,其林分密度、树种多样性和建群种生物量分别变化0.198、0.519、−0.722。基岩裸露度对树种多样性的的正影响较大,对建群种生物量的负影响最大。建群种物量受到土壤厚度、林分密度和物种多样性的正影响,受到基岩裸露度的负影响,其路径系数分别为0.258、0.096、0.084、−0.722,即当土壤厚度、林分密度、物种多样性和基岩裸露度分别变化1时,建群种生物量分别变化0.258、0.096、0.084、−0.722。土壤厚度对建群种生物量有最大的正影响。林分总生物量受到树种多样性和建群种生物量的正影响,其路径系数分别为0.126、1.014。林分总生物量的变化几乎完全受林分建群种生物量的影响。
影响因素 标准化直接影响 标准化间接影响 标准化总影响 基岩
裸露度土壤
厚度树种
多样性林分
密度建群种
生物量基岩
裸露度土壤
厚度树种
多样性林分
密度建群种
生物量基岩
裸露度土壤
厚度树种
多样性林分
密度建群种
生物量物种多样性 0.519 0 0 0 0 0 0 0 0 0 0.519 0 0 0 0 林分密度 0.198 0 0 0 0 0 0 0 0 0 0.198 0 0 0 0 建群种生物量 −0.722 0.258 0.084 0.096 0 0.063 0 0 0 0 −0.659 0.258 0.084 0.096 0 林分总生物量 0 0 0.126 0 1.014 −0.604 0.262 0.085 0.098 0 −0.604 0.262 0.211 0.098 1.014 Table 3. Standard impact coefficient of SEM
河南省淅川县岩溶区建群种生物量和林分总生物量主要受到基岩裸露度、土壤厚度、林分密度和物种多样性的直接或间接影响。基岩裸露度对建群种生物量的总影响系数为−0.659,直接影响系数为−0.722,间接影响系数为0.063;对林分总生物量的总影响系数为−0.604,间接影响系数为−0.604,直接影响系数为0。因为基岩裸露度对林分总生物量直接影响系数可忽略,在结构方程模型中将该路径删除。当基岩裸露度变化1时,建群种生物量和总生物量分别变化−0.659、−0.604。即随着基岩裸露度增加,建群种生物量和林分总生物量下降。土壤厚度对建群种生物量的总影响系数或直接影响系数为0.258;对林分总生物量的总影响系数或间接影响系数0.262。随着土壤厚度的增加,林分建群种生物量和总生物量增加。树种多样性对林分建群种生物量的总影响系数或直接影响系数为0.084,对林分总生物量影响系数为0.211,直接影响系数为0.126,间接影响系数为0.085。随着树种多样性的增加建群种生物量和总生物量缓慢增加。林分密度对林分建群种生物量的总影响系数或直接影响系数为0.096,对林分总生物量的总影响系数或间接影响系数为0.098。建群种生物量对林分总生物量的影响系数为1.014,林分总生物量几乎完全受建群种生物量的决定。岩溶区土壤瘠薄、土壤水肥条件较差、基岩裸露度较大,立地条件差,对植被的适应性要求较高,建群种在立地竞争中占据绝对优势。岩溶区建群种生物量占林分总生物量的77.50%以上(表1),提升建群种的生长条件,能够直接提升林分的总生物量。
基岩裸露度对树种多样性的总影响系数或直接影响系数为0.519,对林分密度的总影响系数或直接影响系数为0.198,即随着基岩裸露度的增加,物种的多样性和林分密度增加。岩溶区基岩的裸露不仅仅造成地被的不连续,而且造成明显的小生境,高度异质的岩溶区物种多样性随着基岩裸露度增加而增加。
-
河南省淅川县岩溶区天然次生林基岩裸露度、土壤厚度、树种多样性、林分密度与建群种生物量和总生物量存在着复杂的耦合关系。基岩裸露度与林分密度为正相关关系,与树种多性为极显著正相关关系(P<0.01),与建群种生物量为极显著负相关关系(P<0.01);土壤厚度与建群种生物量显著正相关关系(P<0.05),树种多样性、林分密度与建群种生物量为正相关关系;建群种生物量与林分总生物量为显著正相关关系(P<0.01)。根据模型拟合结果表明淅川县岩溶区主要影响林分生物量的观测变量基岩裸露度和土壤厚度。基岩裸露度增加,造成地被破碎,土壤不连续,植物生存空间变少、生长的立地条件变差,树种多样性增加,林分密度增加,但树木的胸径树高减少,林分生物量反而下降。土壤厚度是土壤肥力存在和植物生长的重要物质基础,是植被重建最基本的条件[26-27]。土壤厚度增加能够促进植物的生长,增加林分生物量。
基岩裸露度和土壤厚度等作为立地质量的评价指标,能反映立地条件的优劣。一般随立地条件的改善,林分的物种多样性越来越高,结构功能越来越好[28]。同时生态学理论认为群落的稳定性与植物的多样性关系密切,群落的物种多样性是衡量生态系统稳定性的指标[28-29]。淅川县岩溶区立地条件提升时树种多样性会降低,可能是立地质量改善促进了建群种个体的生长,在竞争中占据优势地位,导致其他物种减少,林分复杂程度降低,群落的稳定性降低。岩溶区植被受到强烈的特殊地质环境胁迫,植物群落简单,多样性低,并且岩溶区生长植物既要有石生性、喜钙性以及耐干旱的特点,又必须具有发达、强壮的根系[30],所以岩溶区的适生种、优势种具有明显的特殊性[31-32]。立地条件提升能显著促进建群种的生长,使得林分复杂程度越来越低,多样性降低。
生态系统的变化是一个复杂的过程,并不是简单的一一对应关系[15]。研究系统复杂的生态过程用传统的方法有一定局限性,传统的生长模型大多为直接模型,无法反映隐含的间接关系[33-34]。本研究用结构方程模型来探讨淅川县岩溶区基岩裸露度、土壤厚度、林分密度、树种多样性和建群种生物量、林分总生物量之间的复杂关系。根据模型计算结果可知:因子间既有直接的影响关系,也有间接的关系。基岩裸露度对建群种生物量有直接作用,同时通过影响树种多样性和建群种生物量对林分生物量产生间接作用。土壤厚度、物种多样性、林分密度对建群种生物量有直接作用,并且通过影响建群种生物量对林分总生物量产生间接作用。保护岩溶区生态环境,防止水土流失,保持土壤厚度以及稳定或降低基岩裸露度能够提高岩溶区次生林的生物量,增加干物质积累。
本研究对象是淅川县岩溶区天然次生林,由于岩溶区特殊的地质水文结构,林分的树种多样性低,稳定性较差。同时所选取的样地数量相对较少,模型的最终适配结果虽然存在一定的不足,但是也能反映出岩溶区林分生长与影响因子间的关系。合理改善立地条件,降低基岩裸露度和增加土壤厚度,及时抚育提升树种多样性,增加林分密度,有利于增加岩溶区生物量的积累,促进林分生长,逐步提高岩溶区林分的生态功能。
-
基岩裸露度与林分生物量关系复杂。基岩裸露度能直接降低林分生物量,但又能通过增加树种多样性和林分密度降低间接提升林分生物量。土壤厚度、物种多样性和林分密度直接影响林分的生物量。河南省淅川县岩溶区林分生长与影响因子间的关系耦合结果表明:岩溶区建群种生物量和林分总生物量受到基岩裸露度的极显著负影响(P<0.01),其影响系数分别为−0.659、−0.604;受到土壤厚度的显著正影响(P<0.05),影响系数分别为0.258、0.262;受到树种多样性和林分密度正影响,影响系数分别为0.084、0.211,0.096、0.098。在岩溶区防治水土流失,稳定土壤厚度及基岩裸露度,能改善林分结构等立地条件,增加建群种和林分总生物量,提升岩溶区的生态环境质量。
 DownLoad:
DownLoad: