






-
白及Bletilla striata为兰科Orchidaceae白及属Bletilla多年生草本植物,以其干燥块茎入药,具有止血[1]、促进伤口愈合[2]、抗菌[3]、抗病毒[4]、抗肿瘤[5]等药理作用。此外,白及块茎含较多的胶质多糖,具有抗衰老、美白和抗氧化[2]等功能,因此也被广泛应用于化妆品行业。白及的花色鲜艳美丽,具有较高的观赏价值[6]。由于白及种子无胚乳,自然萌发率低,白及的需求量与野生资源的缺乏产生了巨大的冲突[7],因此开展白及资源的保护及种质资源的研究工作刻不容缓。随着生物技术的发展,除了形态标记[8]、细胞标记[9]、生理生化标记[10]等传统的标记技术外,遗传多样性研究出现了新的基于基因组DNA水平的标记技术——分子标记。分子标记是在群体中易于跟踪和量化的基因位点,并且可能与特定基因或性状相关[11]。与其他标记相比,分子标记具有多态性好、数量多以及高共显性、简单稳定、易于追溯等优点[12]。简单重复序列间扩增(ISSR)和相关序列扩增多态性(SRAP)分子标记是针对非特异性序列进行扩增的,因此对于白及这类基因组序列信息较为匮乏的物种而言是一种快速鉴定的方法[13]。邢佳鑫等[14]应用简单重复序列(SSR)和ISSR标记对外观较相似的白及样本进行遗传多样性分析,探索了快速鉴定白及组培苗真伪的新方法;洪建聪等[15]利用筛选出的15对SRAP引物组合将50份白及样本分成6个大类;陈锦等[16]利用ISSR和随机扩增多态性(RAPD)标记研究白及种质资源的遗传多样性,结果显示:白及种源间的遗传差异与地理距离相关,还发现ISSR不受时间和地点的限制,比RAPD有更高的多态性比率,结果相对更稳定可靠;孙宇龙等[17]通过SRAP标记将12个白及野生种源聚类成3支系,并发现遗传距离和地理距离间有显著正相关性,为白及野生资源的就地保护和迁地保护提供了理论依据;WANG等[18]使用ISSR标记分析了白及再生植株的克隆保真度,结果发现:再生植株与母株的相似度接近100.0%,表明通过愈伤组织和原球茎培养的再生植株在遗传上是稳定的。
目前,同时应用2种分子标记技术在分子水平上对白及种质资源进行的相关研究较少。本研究采用ISSR和SRAP分子标记技术,对从浙江省、云南省、贵州省和四川省搜集的32个白及种源进行亲缘关系及遗传多样性的初步分析,揭示白及个体间与种源间的遗传分化,为白及资源的保护和利用提供参考。
-
2020年6—7月在浙江、云南、贵州、四川等22个地区采集32个白及种源,并将采集到的白及叶片洗净擦干,经液氮速冻后放置−80 ℃超低温冰箱备用。种源基本信息见表1。
编号 种源地 编号 种源地 编号 种源地 编号 种源地 1 浙江温州泰顺 9 云南丽江永胜 17 四川乐山沐川(2) 25 四川汶川水磨(2) 2 浙江台州天台 10 云南普洱思茅 18 四川雅安石棉 26 四川汶川水磨(3) 3 浙江衢州江山 11 云南曲靖会泽 19 四川内江东兴(1) 27 四川都江堰虹口(1) 4 浙江金华磐安 12 贵州遵义正安 20 四川内江东兴(2) 28 四川都江堰虹口(2) 5 浙江宁波象山 13 贵州黔西南安龙 21 四川内江东兴(3) 29 四川成都大邑 6 浙江杭州临安 14 贵州遵义赤水 22 四川内江东兴(4) 30 四川成都彭州(1) 7 云南红河蒙自(1) 15 贵州毕节大方 23 四川内江东兴(5) 31 四川成都彭州(2) 8 云南红河蒙自(2) 16 四川乐山沐川(1) 24 四川汶川水磨(1) 32 四川甘孜康定 Table 1. Source information of B. striata samples
-
主要仪器:JXFSTPRP-24L组织快速研磨仪(中国上海净信实业发展有限公司)、TU-100恒温金属浴(中国上海一恒科技有限公司)、Colibri超微量分光光度计(德国Titertek Berthold公司)、Centrifuge 5415 R高速冷冻离心机(德国Eppendorf公司)、Mini-P25微孔板离心机(中国杭州奥盛仪器有限公司)、T100 Thermal Cycler PCR仪、PowerPac HC高电流电泳仪[伯乐生命医学产品(中国上海)有限公司]、JY04S-3C凝胶成像系统(中国北京君意东方电泳设备有限公司)、B2042BluEye TM蓝光切胶仪(中国苏州宇恒生物科技有限公司)。
主要试剂:植物基因组DNA快速抽提试剂盒(中国上海生工生物股份有限公司)、2×Flash PCR MasterMix(中国北京康为世纪生物科技有限公司)、10 × loading buffer、DL 15000 DNA Marker、DL 2000 DNA Marker(日本TAKARA公司)。β-巯基乙醇、聚乙烯吡咯烷酮(PVP)、乙醇、氯仿、异戊醇等均为中国上海国药集团化学试剂有限公司生产;ISSR引物和SRAP引物由北京擎科生物科技有限公司合成。
-
取洁净干燥的白及嫩叶,采用植物基因组DNA快速抽提试剂盒提取样本DNA,并用质量分数为1%的琼脂糖凝胶电泳法检测完整性,选择条带清晰明亮且无拖尾与杂带的DNA,用超微量分光光度计检测DNA的浓度和纯度,结合DNA浓度及电泳的结果,将较优的DNA样品液用双蒸水(ddH2O)稀释至20 mg·L−1,放置冰箱−20 ℃保存备用。
-
本研究采用的ISSR引物为哥伦比亚大学公布的第9套100个ISSR通用引物序列;SRAP引物为参考前人[19-20]设计的238对SRAP引物组合,共包含14条上游引物(Me1~Me14)和17条下游引物(Em1~Em17)。引物的筛选经过初筛和复筛2个步骤:从32个供试材料中选取4个表型性状差异较明显的材料作为样本DNA,对引物进行初筛,选出扩增产物主带明显、条带清晰的引物。经过引物的初筛后,选择所有样本DNA作为模板,针对初筛后的引物进行复筛,选择多态性高、扩增条带清晰、重复性好的引物作为本研究的引物。根据每条引物的退火温度(Tm) ± 5 ℃的范围,设置8个温度梯度进行聚合酶链式反应(PCR);根据条带得出每个引物的最适退火温度。
-
ISSR-PCR扩增体系(20 μL):1 μL DNA模板、1 μL引物(20 mg·L−1)、10 μL 2×Flash PCR MasterMix、8 μL ddH2O;ISSR-PCR扩增程序:98 ℃预变性30 s;94 ℃变性10 s;退火(每个引物的退火温度不同)15 s,72 ℃延伸15 s;共35个循环,72 ℃延伸1 min,4 ℃保存。
SRAP-PCR扩增体系(20 μL):1 μL DNA模板、1 μL引物(20 mg·L−1)、10 μL 2×Flash PCR MasterMix、7 μL ddH2O;SRAP-PCR扩增程序:98 ℃预变性30 s;94 ℃变性10 s;退火(每个引物的退火温度不同)15 s,72 ℃延伸15 s;共38个循环,72 ℃延伸1 min,4 ℃保存。
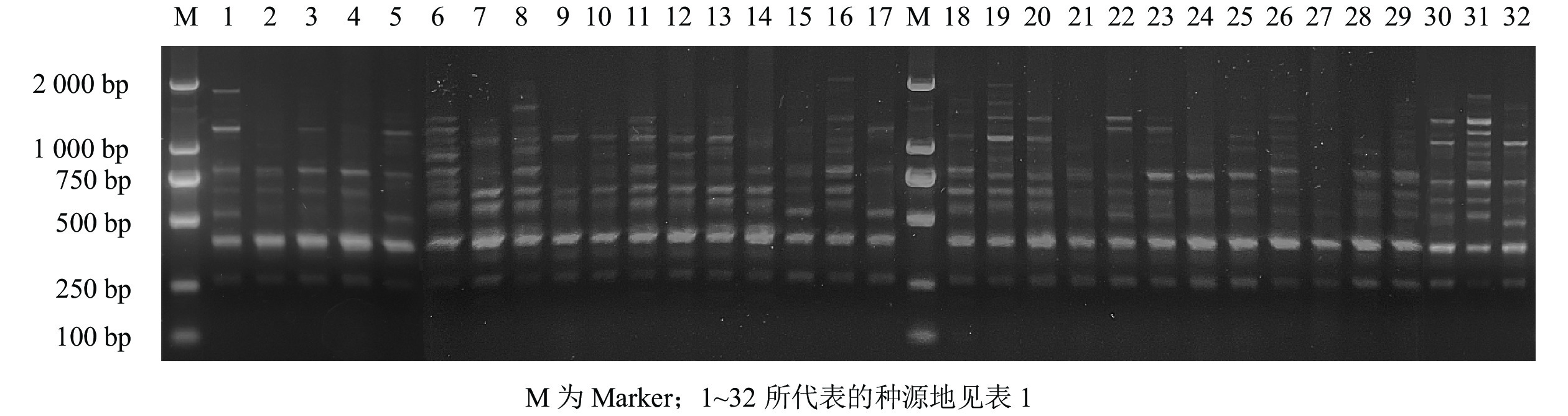
PCR扩增产物在含有Gelred核酸染料的质量分数为1.5%的琼脂糖凝胶1×TAE缓冲液中,130 V,电泳50 min后,经凝胶成像系统(JY04S-3C)拍照保存。
-
人工读取电泳结果数据,并进行统计处理。将条带中清晰可见以及可重复的标记为“1”,同一位置上条带缺失或者弱带标记为“0”,形成“0、1”数据矩阵,分别得到ISSR、SRAP及综合两者(SSR+SRAP)的数据。通过Excel表格,统计每个引物或每对引物组合扩增的条带以及多态性条带数,计算多态位点百分率(PPB),利用Popgene 32.0软件计算等位基因数(Na)、有效等位基因数(Ne)、Nei’s基因多样性指数(H)、Shannon’s多态性信息指数(I)、种源总基因多样性(Ht)、种源内基因多样性(Hs)、基因分化系数(Gst = 1−Hs/Ht)和基因流(Nm)等遗传多样性参数,并计算种源间的遗传距离和遗传一致性,使用OmicStudio工具(
https://www.omicstudio.cn/tool )对遗传距离进行主坐标(PCoA)分析,利用NTSYS-pc 2.10e软件对种源进行聚类分析,并绘制非加权组平均法(UPGMA)树状图。 -
提取白及基因组DNA后,经琼脂糖凝胶电泳检测DNA质量,并通过超微量分光光度计检测其浓度、纯度,均符合要求,可进行下一步试验。
-
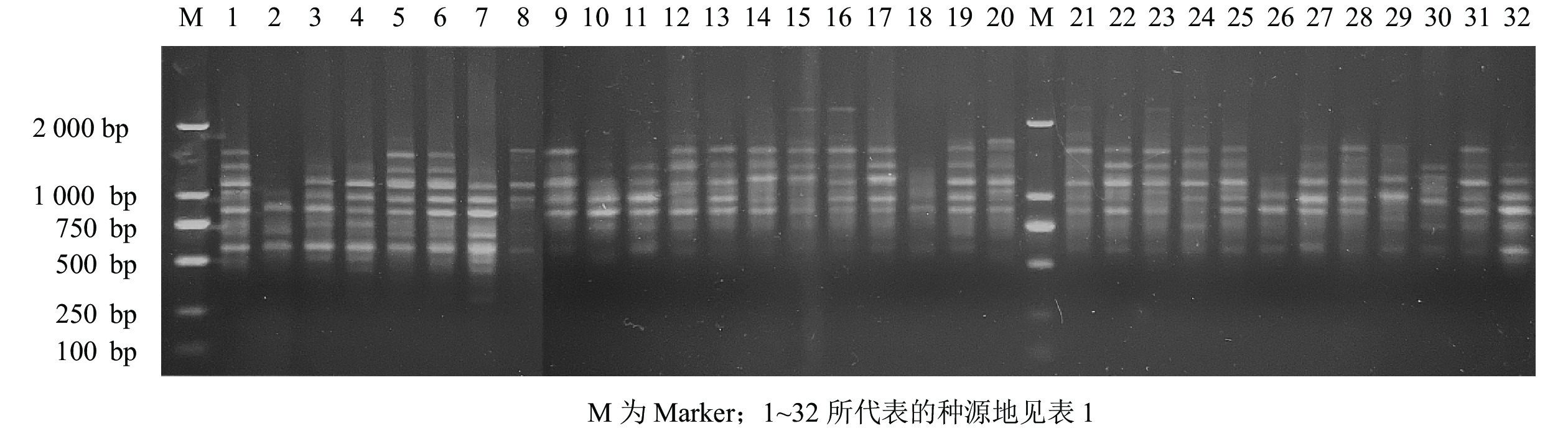
经过2次筛选,共获得11条条带清晰、多态性高且重复性好的引物(表2)。11条引物共扩增出188条条带,多态性条带有174条,PPB为92.20%。11条引物平均扩增条带数为17.09条,平均多态性条带数为15.82条。扩增位点数最多的引物是UBC876,扩出了25条条带;其次是引物UBC868和UBC881,均扩增出20条条带;扩增条带最少的引物是UBC855,只扩增出9条条带,PPB为64.71%~100.00%,其中,引物UBC827、UBC842、UBC855、UBC876、UBC879和UBC881的PPB为100%,均表现出极高的多态性,占总引物的54.55%;UBC880的PPB最低,为64.71%。以引物UBC876为例,它对所采集白及样本的ISSR-PCR的扩增结果见图1。
引物编号 ISSR引物 序列
(5′→3′)扩增条
带数/条多态性条
带数/条PPB/% 引物编号 ISSR引物 序列
(5′→3′)扩增条
带数/条多态性条
带数/条PPB/% 1 UBC823 (TC)8C 15 12 80.00 8 UBC876 (GATA)2(GACA)2 25 25 100.00 2 UBC826 (AC)8C 18 17 94.44 9 UBC879 C(TTCAC)3 15 15 100.00 3 UBC827 (AC)8G 16 16 100.00 10 UBC880 (GGAGA)3 17 11 64.71 4 UBC842 (GA)8(C/T)G 18 18 100.00 11 UBC881 (GGGTG)3 20 20 100.00 5 UBC855 (AC)8(C/T)T 9 9 100.00 合计 188 174 6 UBC866 (CTC)6 15 12 80.00 平均 17.09 15.82 92.20 7 UBC868 (GAA)6 20 19 95.00 Table 2. ISSR primer information and amplification results
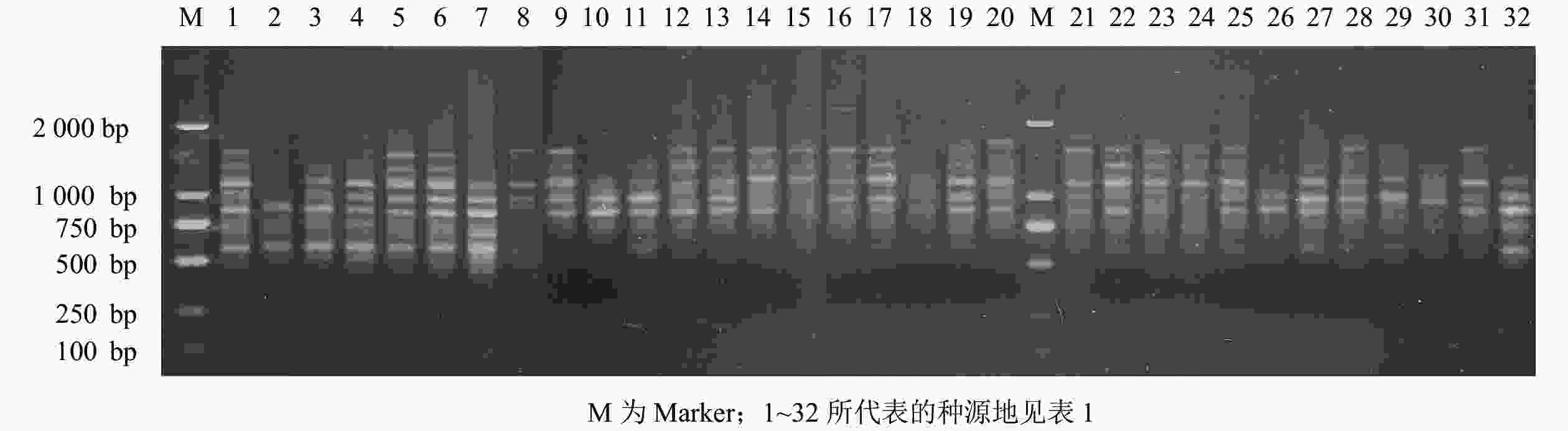
Figure 1. Amplification results of B. striata with primer UBC876 of ISSR
-
经过2次筛选,共获得11对条带清晰、多态性高且重复性好的引物组合(表3)。11对引物组合共扩增出216条条带,多态性条带有202条,多态性比率占93.52%,11对引物平均扩增条带数为19.64条,平均多态性条带数为18.36条。引物组合Me13-Em9扩增位点数最多,扩出28条条带;其次为引物组合Me3-Em2和Me11-Em10,均扩增出22条条带;扩增条带最少的引物组合为Me12-Em6,仅扩增出16条条带。引物的PPB为83.33%~100.00%,其中,引物组合Me9-Em13、Me11-Em10和Me12-Em6的PPB最高,均为100.00%,占总引物的25.93%;引物组合Me11-Em12的PPB最低,为83.33%。以引物组合Me9-Em13为例,它对部分白及基因组DNA的ISSR-PCR的扩增结果见图2。
引物
编号SRAP引物 正向引
物(5′→3′)反向引
物(5′→3′)扩增条
带数/条多态性条
带数/条PPB/% 引物
编号SRAP引物 正向引
物(5′→3′)反向引
物(5′→3′)扩增条
带数/条多态性条
带数/条PPB/% 1 Me1-Em12 BATA DCAT 19 17 89.47 8 Me11-Em12 BACG DCAT 18 15 83.33 2 Me3-Em2 BAAT DTGC 22 20 90.91 9 Me12-Em6 BAAA DGCA 16 16 100.00 3 Me5-Em5 BAAG DAAC 18 17 94.44 10 Me12-Em7 BAAA DCAA 17 16 94.12 4 Me6-Em2 BACT DTGC 19 16 84.21 11 Me13-Em9 BAAC DCGA 28 27 96.43 5 Me9-Em13 BACA DCTA 18 18 100.00 合计 216 202 6 Me9-Em14 BACA DCTC 19 18 94.74 平均 19.64 18.36 93.52 7 Me11-Em10 BACG DCAG 22 22 100.00 说明:B表示TGAGTCCAAACCGG;D表示GACTGCGTACGAATT Table 3. SRAP primer information and amplification results
Figure 2. Amplification results of B. striata with primer combination Me9-Em13 of SRAP
-
利用Popgene 32.0软件对32个白及种源的遗传距离和遗传一致度进行计算,表明在ISSR研究中,遗传一致度为0.5745~0.8989,遗传距离为0.1065~0.5543。遗传一致度最高的是21号四川内江东兴(3)和22号四川内江东兴(4)种源,为0.8989;它们的遗传距离最小,为0.1065,说明其亲缘关系较近。遗传一致度最低的是13号贵州黔西南安龙和25号四川汶川水磨(2)种源,它们的遗传一致度均为0.5745;它们的遗传距离最大,为0.5543,说明其亲缘关系较远。在SRAP研究中,遗传一致度为0.5231~0.8102,遗传距离为0.2105~0.5231。遗传距离最小的是2号浙江台州天台和3号浙江衢州江山种源;遗传距离最大的是4号浙江金华磐安和30号四川成都彭州(1)种源。综合ISSR和SRAP数据,遗传一致度为0.5668~0.8119,遗传距离为0.2084~0.5677;遗传距离最小的是25号四川汶川水磨(2)和32号四川甘孜康定种源,遗传距离最大的是4号浙江金华磐安和11号云南曲靖会泽种源。
-
将32个种源按地区划分为4个群体(浙江、云南、贵州和四川),使用Popgene 32.0软件分析种源的遗传多样性指数(表4)。在ISSR研究中,32个种源在物种水平上,Na为1.9255,Ne为1.4619,H为0.2819,I为0.4327,PPB为92.55%。在群体水平上,Na为1.5000~1.7926;Ne为1.3176~1.4393;H为0.1824~0.2607;I为0.2717~0.3940;PPB为50.00%~79.26%。可见,相对于物种水平而言,群体间的遗传多样性水平较低。在SRAP研究中,变异水平也是物种大于群体,与ISSR研究结果相似。综合ISSR和SRAP数据,32个种源在物种水平上,Na为1.9307,Ne为1.4988,H为0.2971,I为0.4509,PPB为93.07%。在群体水平上,Na为1.5000~1.8342,平均为1.6349;Ne为1.3281~1.4660,平均为1.3888;H为0.1877~0.2756,平均为0.2256;I为0.2780~0.4163,平均为0.3369;PPB为50.00%~83.42%,平均为63.49%,也是群体间的遗传多样性水平更低。其中,从ISSR、SRAP及综合ISSR和SRAP的研究结果来看,Na、Ne、H、I及PPB数值最小的均是贵州省种源,浙江省种源次之,各数值最大的均是四川种源,说明四川白及种源具有较高的遗传多样性,贵州白及种源遗传多样性较低。
研究方法 种源地 Na Ne H I PPB/% 浙江 1.5000 1.3215 0.1841 0.2729 50.00 ISSR 云南 1.6489 1.4042 0.2328 0.3473 64.89 贵州 1.5000 1.3176 0.1824 0.2717 50.00 四川 1.7926 1.4393 0.2607 0.3940 79.26 群体水平 1.6104 1.3707 0.2150 0.3215 61.04 物种水平 1.9255 1.4619 0.2819 0.4327 92.55 SRAP 浙江 1.5833 1.3753 0.2153 0.3191 58.33 云南 1.6713 1.4163 0.2432 0.3631 67.13 贵州 1.5000 1.3373 0.1923 0.2835 50.00 四川 1.8704 1.4891 0.2886 0.4357 87.04 群体水平 1.6563 1.4045 0.2349 0.3504 65.63 物种水平 1.9352 1.5309 0.3103 0.4667 93.52 ISSR+SRAP 浙江 1.5446 1.3503 0.2007 0.2976 54.46 云南 1.6609 1.4107 0.2384 0.3558 66.09 贵州 1.5000 1.3281 0.1877 0.2780 50.00 四川 1.8342 1.4660 0.2756 0.4163 83.42 群体水平 1.6349 1.3888 0.2256 0.3369 63.49 物种水平 1.9307 1.4988 0.2971 0.4509 93.07 Table 4. Genetic diversity of B. striata
-
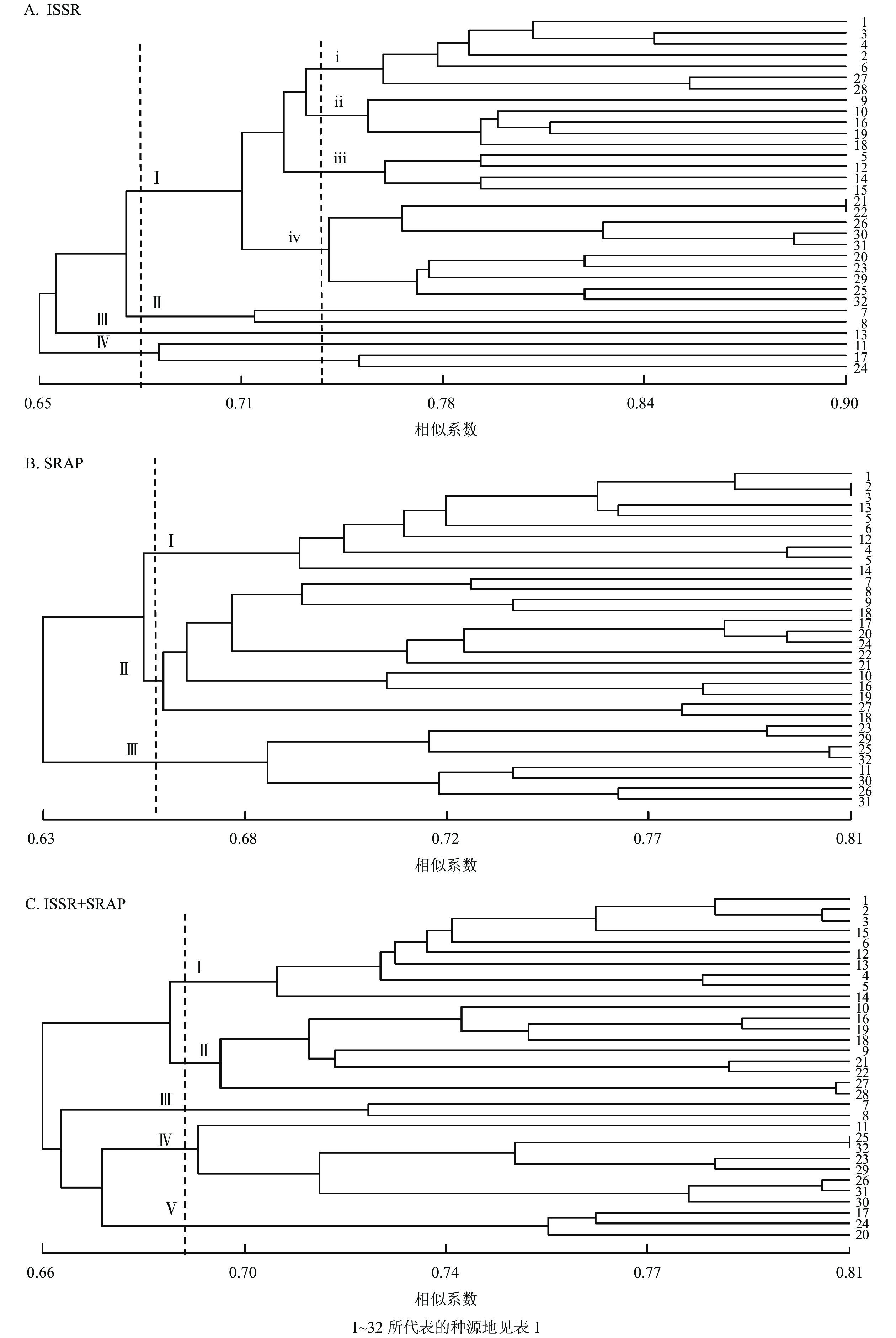
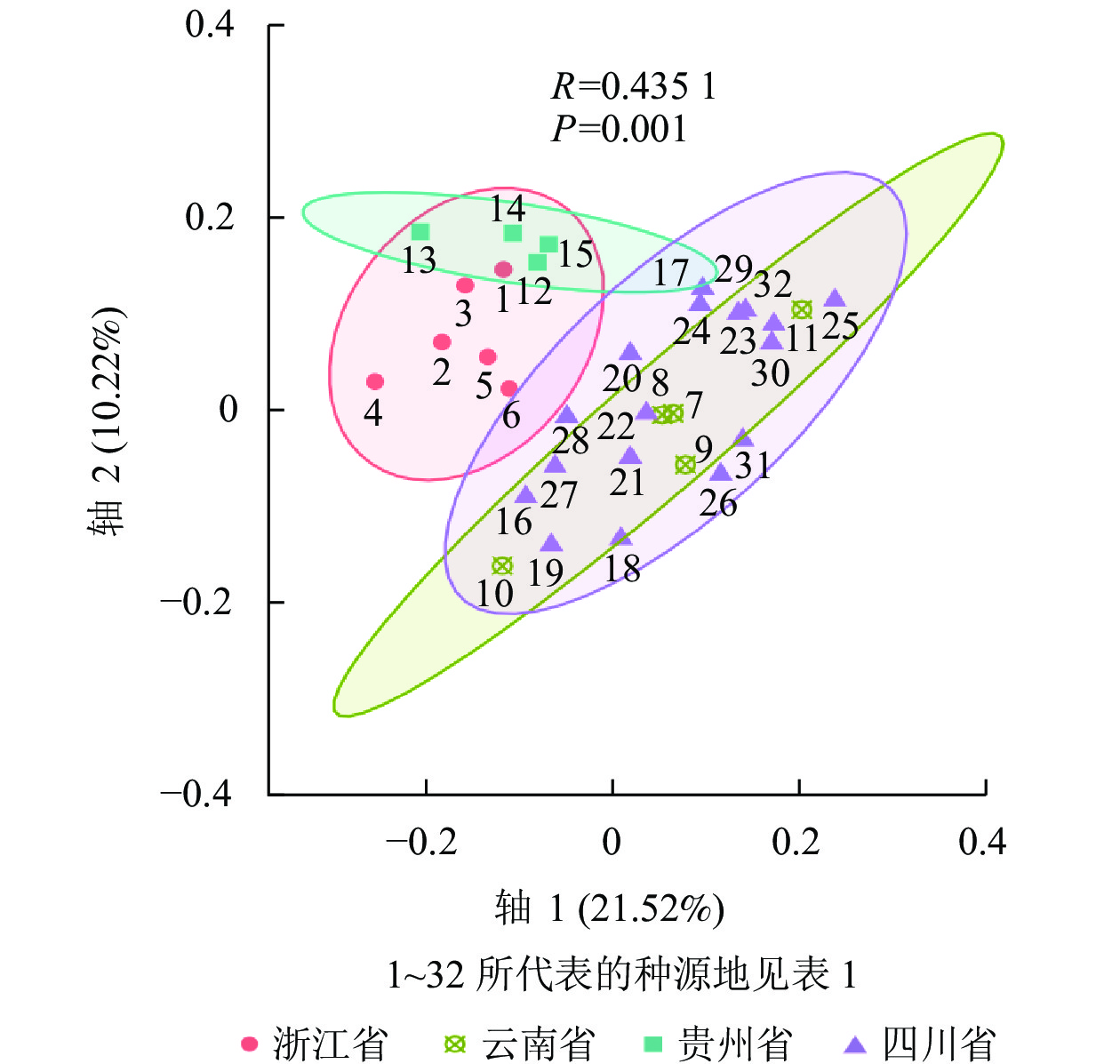
利用NTSYS-pc 2.10e软件将32个种源的标记结果进行聚类分析,得到相应的UPGMA树状图(图3)。ISSR的UPGMA树状图显示:32个种源的种质资源遗传相似系数为0.650~0.900,变幅为0.250。其中,在遗传相似系数为0.680时,可将32个白及种源分为4个类群:Ⅰ类群包括26个种源,其中浙江种源6个,云南种源2个、贵州种源3个,四川种源15个;Ⅱ类群包括2个云南红河蒙自种源;Ⅲ类群仅包括13号贵州黔西南安龙种源,Ⅳ类群包括11号云南曲靖会泽种源、17号四川乐山沐川(2)种源和24号四川汶川水磨(1)种源共3个。在遗传相似系数为0.738时,Ⅰ类群又可分为4个类群(ⅰ、ⅱ、ⅲ、ⅳ类群) (图3A)。SRAP的UPGMA树状图显示:遗传相似系数为0.630~0.810,变幅为0.180。其中,在遗传相似系数为0.658时,可将样本分为3个类群:Ⅰ类群包括浙江种源6个和贵州种源4个;Ⅱ类群包括云南种源4个和四川种源10个;Ⅲ类群包括云南曲靖会泽种源和四川种源7个(图3B)。ISSR+SRAP的UPGMA树状图显示:遗传相似系数为0.660~0.810,变幅为0.150。其中,在遗传相似系数为0.688时,可将样本分为5个类群:Ⅰ类群包括浙江种源6个和贵州种源4个;Ⅱ类群包括云南种源2个和四川种源7个;Ⅲ类群包括2个云南红河蒙自种源;Ⅳ类群包括云南曲靖会泽种源和四川种源7个;Ⅴ类群包括四川种源3个(图3C)。使用OmicStudio工具将ISSR+SRAP的标记结果进行PCoA分析(图4),结果与UPGMA聚类结果基本一致。
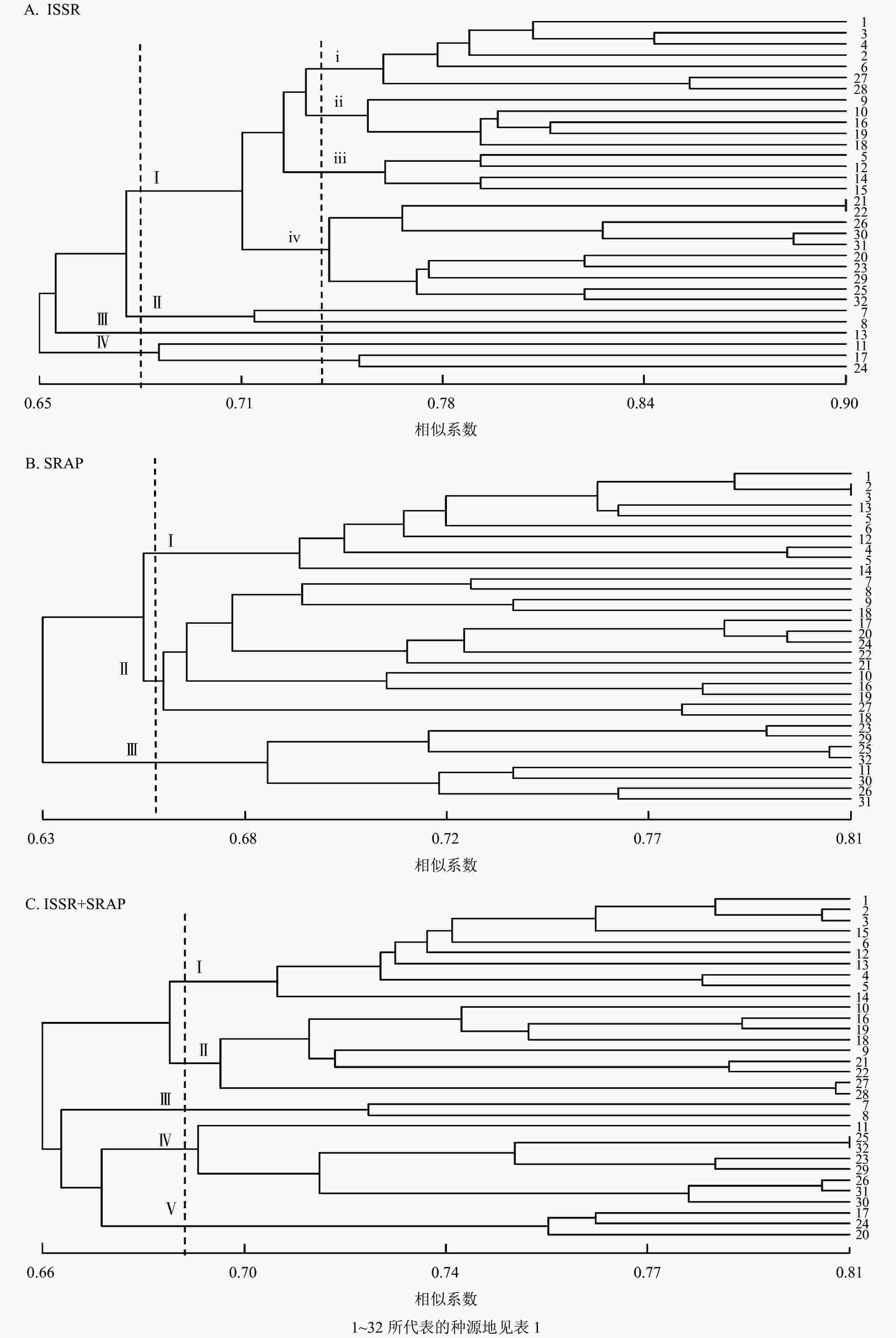
Figure 3. UPGMA cluster map of 32 provenance of B. striata
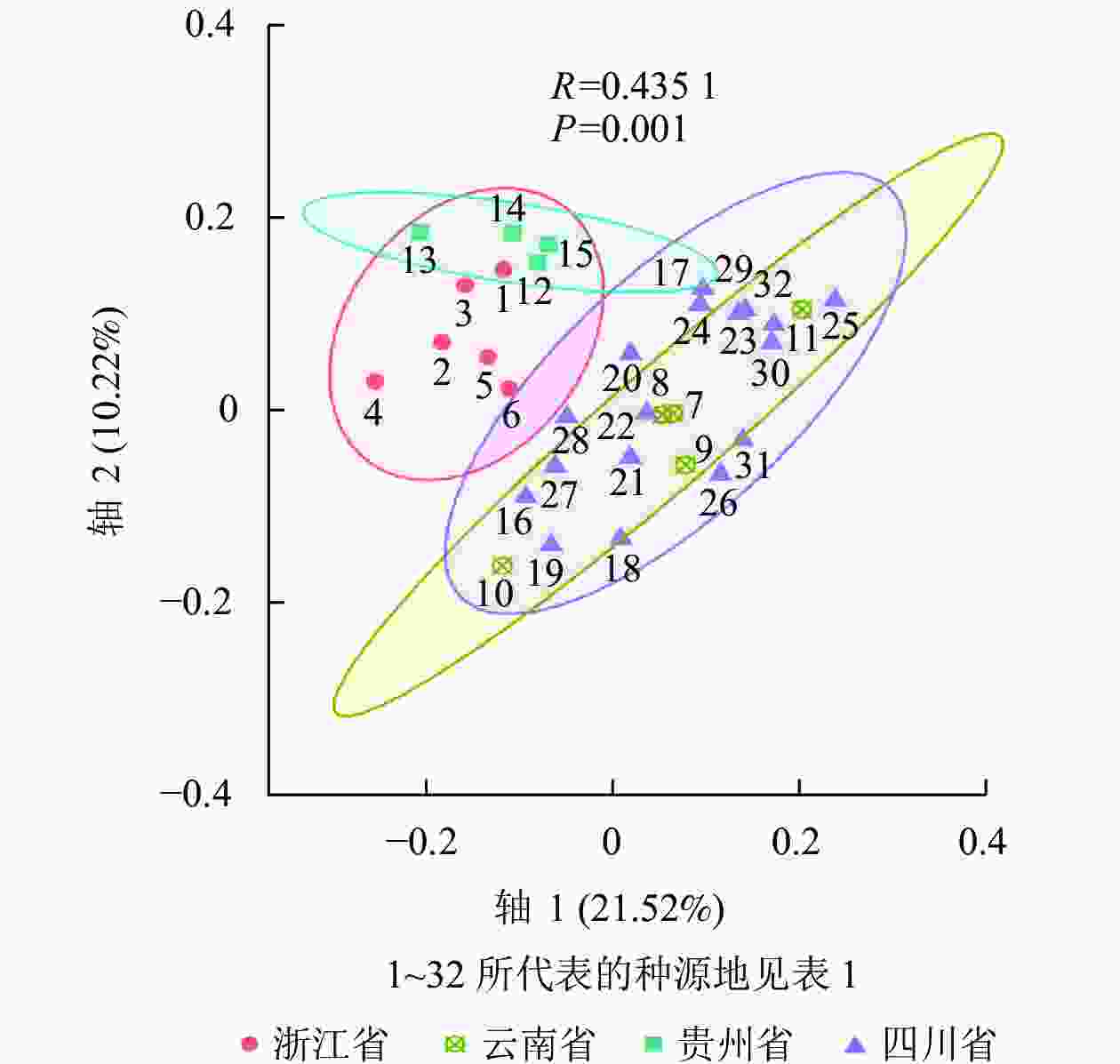
Figure 4. PCoA analysis of ISSR and SRAP markers data
-
陈锦等[16]、周天华等[21]、和志娇等[22]、吴劲松[23]、孙宇龙等[17]分别应用RAPD、SSR、ISSR、扩增片段长度多态性(AFLP)和SRAP等单一分子标记技术研究了白及种质资源的遗传多样性,其中每个引物最少扩增得到1条多态性条带,最多的有19条,PPB最低为46.10%,最高则达90.48%。本研究综合了ISSR和SRAP等2种标记方法对32个白及种源进行了分析,筛选出的11条ISSR引物,共扩增出188条条带,多态性条带174条,平均每条引物15.82条,最多的达25条(UBC876),PPB为92.20%;筛选出的11对SRAP引物组合,共扩增出216条条带,多态性条带有202条,平均每对引物19.64条,最多达27条(Me13-Em9),PPB为93.52%。结果充分体现了供试白及种源在分子水平上具有丰富的遗传多样性,同时也验证了ISSR和SRAP标记技术在白及种源多态性检测方面的高效率,且SRAP的检测效率略高于ISSR。综合ISSR和SRAP数据分析白及遗传多样性指数发现:四川省白及种源的Na、Ne、H、I及PPB最大,云南种源次之,贵州种源最小,说明四川白及种源的遗传多样性水平最高,适应环境能力较强。
和志娇等[22]采用ISSR标记对不同种源的野生白及进行遗传多样性评价,发现不同地理位置和不同海拔采集到的白及个体的遗传距离有差异;孙宇龙等[17]也发现:遗传距离和地理距离间有显著正相关性;洪建聪等[15]和GUO等[24]则发现:白及个体的遗传距离与地理距离无直接关联。本研究综合了ISSR和SRAP标记数据发现:21号四川内江东兴(3)种源与22号四川内江东兴(4)种源、2号浙江台州天台种源与3号浙江衢州江山种源、25号四川汶川水磨(2)种源与32号四川甘孜康定种源的遗传距离较小,亲缘关系较近;13号贵州黔西南安龙种源与25号四川汶川水磨(2)种源、4号浙江金华磐安种源与11号云南曲靖会泽种源和30号四川成都彭州(1)种源遗传距离较大,亲缘关系较远。总体上同省的样品间遗传距离相对较小,这些在UPGMA聚类图和PCoA分析图上也有比较直观的体现。UPGMA聚类显示:聚为一类的白及种源大多来自同一省份,而在PCoA分析图中,云南种源与四川种源的亲缘关系较近,浙江种源与贵州种源的亲缘关系较近。整体上各个种源间的遗传距离与地理位置呈现出一定的相关性,但又不完全按照地理位置的远近聚类,有些甚至有所交叉。
物种的遗传特性主要受到生态环境、生物学特征、繁殖方式以及各种人为活动的影响[25],野生白及由于长期的异花授粉和自然选择,其遗传背景复杂[24];人工栽培白及则多是从山上采挖假鳞茎进行种植,品种混杂,品质参差不齐。又由于白及对生境的依赖性较高,因此生境的破坏和片段化都会影响其遗传多样性,所以在保护物种多样性时,首先应保护生态环境。本研究的某些白及种源可能是来自不同地区的引种,在土壤、养分、温度和水分等环境条件的作用下,导致地理距离较近的种源遗传性状产生了相似的变异,从而使不同种源间的遗传距离与地理距离存在一定的重合性。总的来说,本研究32个白及种源的遗传多样性水平较高。
-
本研究综合利用ISSR和SRAP等2种分子标记技术,对浙江、云南、贵州、四川32个白及种源进行遗传多样性分析。结果显示:种源间具有较高的遗传多样性,且ISSR和SRAP技术均可有效揭示白及的遗传多样性和亲缘关系,运用2种分子标记技术也使得研究的可信度更高。
Genetic diversity analysis of Bletilla striata germplasm by ISSR and SRAP markers
doi: 10.11833/j.issn.2095-0756.20210690
- Received Date: 2021-10-09
- Accepted Date: 2022-04-07
- Rev Recd Date: 2022-03-31
- Available Online: 2023-04-03
- Publish Date: 2023-04-20
-
Key words:
- Bletilla striata /
- genetic diversity /
- ISSR marker /
- SRAP marker
Abstract:
| Citation: | WANG Jie, ZHU Xipeng, WANG Tengfei, et al. Genetic diversity analysis of Bletilla striata germplasm by ISSR and SRAP markers[J]. Journal of Zhejiang A&F University, 2023, 40(2): 321-329. DOI: 10.11833/j.issn.2095-0756.20210690

|
 DownLoad:
DownLoad: