






-
紫茎Stewartia sinensis属于山茶科Theaceae紫茎属Stewartia的落叶小乔木或乔木树种,为第三纪孑遗植物,分布广泛,在浙江、安徽、福建等12个省均有生长,垂直生长于海拔550~2 200 m的常绿或混交阔叶林[1-2]。紫茎高可达20 m,枝叶繁茂,树形优美,树干片状剥落呈多种色彩,白花黄蕊清秀淡雅,极具观赏性,宜栽植于庭院。紫茎属植物间断分布于东亚至北美大陆[2],对于研究东亚至北美植物区系及古植物地理具有重要的科学意义[3]。由于自然更新能力差和生长缓慢,加之生境破坏、人类活动等影响,野生种群数量不断减少[4],紫茎在种间竞争中处于劣势,已被列入国家三级珍稀濒危植物[5]和浙江省重点保护野生植物[3]。幼苗是植物生活史最为脆弱的环节之一,木本植物的天然更新以及幼苗幼树的建成,与森林群落内部至群落外缘光强的异质性以及土壤肥沃与否存在密切的生态关系[6-7]。通过对野外紫茎分布地段调查发现:群落内紫茎的幼苗和幼树数量极为稀少,但在群落外缘的路旁生长定居一定数量的幼苗幼树。推测春夏期间,常绿落叶阔叶森林群落高度郁闭状况必然影响紫茎幼苗的生长光强,从而影响其定居过程。光合作用是植物幼苗其他生命活动的基础,光合作用与氮素水平存在着直接或间接的关系[8-9]。森林群落植被密度高,濒危植物自身的生态适应性相较于其他植物弱,有限的氮素成为限制濒危植物生长的因素之一[10]。森林群落内的紫茎幼苗因光照和养分的竞争限制作用,无法占据适宜的生态位而生长不良,甚至出现幼苗枯死现象。已有紫茎的研究成果主要涉及地理区系分布[1-2]、人工繁殖技术[11-12]、群落特征[13-14]、药用价值[15]等方面,有关幼苗生长适应性与环境的生态关系尚未展开研究,因此迫切需要研究光照强度和氮素水平的耦合效应对紫茎幼苗光合特性的影响,以期在濒危植物紫茎的保护和扩繁过程,为紫茎幼苗阶段的光照和氮素调控提供理论依据。为此,本研究选择紫茎幼苗为材料,设置光强和氮素双因素控制试验,模拟常绿落叶阔叶林中到林缘光环境和土壤肥力的梯度变化,测定不同处理组合紫茎幼苗的光响应、光合色素以及叶绿素荧光,筛选适宜紫茎幼苗生长的光强氮素组合处理。
-
2021年3月从清凉峰国家级自然保护区苗圃(30°09′28″N, 118°86′37″E)选取长势基本一致的1年生紫茎实生幼苗[平均苗高(32.41±4.26) cm,地径(0.49±0.06) cm]移至浙江农林大学东湖校区试验地。试验地海拔为42 m,全年降水量为1 628.6 mm,年平均气温为16.4 ℃,全年无霜期约237 d。幼苗栽植于口径24 cm、高20 cm、底径14 cm的花盆内,每盆1株。盆栽基质为当地黄壤[全氮(1.53±0.01) g·kg−1,碱解氮(185.0±10.5) mg·kg−1,有效磷(8.7±0.1) mg·kg−1,速效钾(278.0±14.9) mg·kg−1,有机质(31.6±0.1)%,pH (土水质量比为1.0∶2.5) 5.07±0.01]。盆栽苗放置于2层遮光大棚内,缓苗1个月后,将60盆紫茎随机分成12组,每组5盆。依据紫茎所在常绿或混交阔叶林的郁闭度调查结果,为量化空旷地、疏林、中等郁闭和密林的光照强度,设置3个遮光处理和1个全光对照处理。遮光大棚采用黑色尼龙遮光网覆盖并适当开洞模拟林窗,大棚长30.0 m×宽11.5 m×高3.2 m,通风状况良好,透光率分别为轻度遮光(44.79±0.51)%(L1)、中度遮光(19.60±0.23)%(L2)、高度遮光(7.25±0.10)%(L3)和全光100.00%(L0),透光率为在3个晴天正午所测的光量子通量密度(PPFD),取均值。参考尹丽等[10]对麻疯树Jatropha curcas幼苗的施氮量,在每个遮光处理下设置3个氮素处理,分别为低氮0.2 g· kg−1 (N1)、高氮0.6 g·kg−1 (N2)和不施氮对照 (N0),所用氮肥为硫酸铵[(NH4)2SO4],施氮以盆中干土计,即1 kg土分别施用约1 g (N1)和3 g (N2)硫酸铵,采用土表施氮法用适量水溶解稀释后分3次施用,每隔10 d施肥1次,施氮后3 d内不浇水以防流失。处理90 d后测定各项指标。
-
使用Li-6800光合仪,于2021年5月25日至6月4日(测定期间为晴天) 8:00—11:30,使用Li-6800光合仪测定紫茎的光合参数。对待测功能叶片用1200 μmol·m−2·s−1的光强进行20 min光诱导,设置光合有效辐射(PAR)梯度为2 000、1 500、1 200、1 000、800、500、300、100、50、30、10和0 μmol·m−2·s−1。叶室控制温度为(25.0±0.5) ℃,相对湿度为(40±5)%,叶室二氧化碳(CO2)摩尔分数为400 μmol·mol−1,气体流速为500 μmol·s−1。每个处理随机取3株,测定净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)和胞间CO2摩尔分数(Ci)等参数;用直角双曲线修正模型[16]拟合光响应模型,根据模型计算最大净光合速率(Pmax)、光饱和点(LSP)、光补偿点(LCP)、表观量子效率(AQY)和暗呼吸速率(Rd)。
-
采用乙醇浸提法测定光合色素质量分数[17]。选取生长良好且成熟的紫茎叶片,剪成2 mm宽的条状碎片,称取0.2 g置于试管中,加入体积分数为95%的乙醇10 mL,置于黑暗处浸提至叶组织完全变白,将提取液倒入光径1 cm的比色杯内,用UV-2700型紫外分光光度计在665、649和470 nm的波长下读取吸光度,并计算总叶绿素(Chl)、叶绿素a(Chl a)、叶绿素b(Chl b)和类胡萝卜素(Car)的质量分数。
-
于2021年5月20—23日,每个处理选3株紫茎,每株选3片叶片,对待测叶片进行40 min的暗适应处理,使用YZQ-500动态荧光仪(翼鬃麒科技公司)测定叶片荧光参数初始荧光(Fo)和最大荧光(Fm),并采用李鹏民等[18]的方法计算暗适应下光系统Ⅱ(PSⅡ)潜在活性(Fv/Fo)、PSⅡ最大光能转化效率(Fv/Fm)和以吸收光能为基础的性能指数(PIABS)。
-
用WPS 2019对原始数据进行整理,SPSS 26进行单因素方差分析(one-way ANOVA)和最小显著差异分析(LSD),Origin 2018制作光响应进程变化图,双因素方差分析法(two-way AVOVA)检验光强与氮素对紫茎幼苗光响应参数、光合色素质量分数和叶绿素荧光参数的交互影响效应。
-
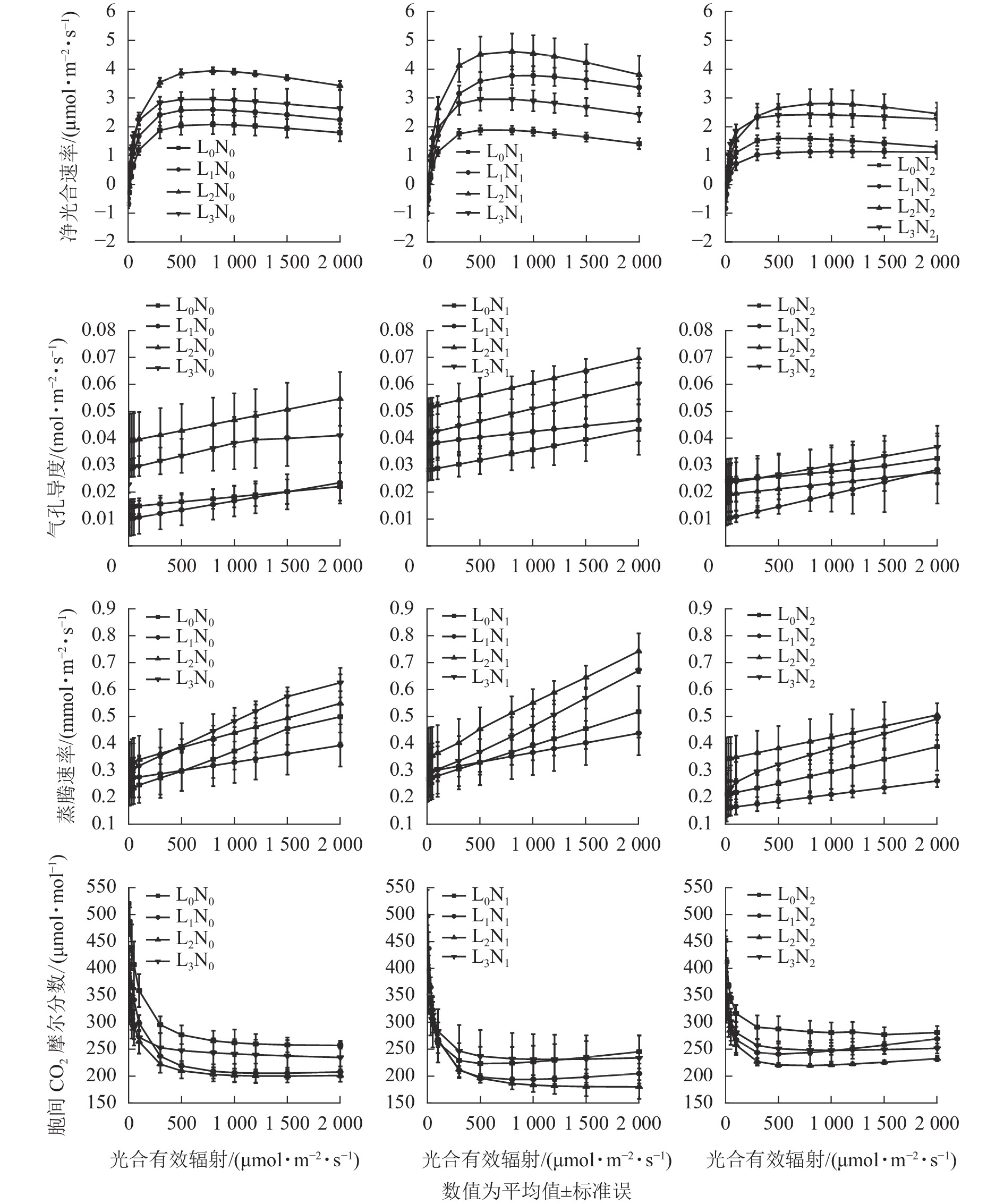
随着PAR增加,紫茎幼苗Pn总体呈先升高后趋于平缓的变化特点,当PAR<100 μmol·m−2·s−1时,不同处理下紫茎幼苗的Pn较为接近(图1)。未施氮条件下,L2的紫茎幼苗Pn高于其他3种遮光处理;N1氮素水平下,不同遮光处理的紫茎幼苗Pn从大到小依次为L2、L1、L3、L0,适度施氮明显提高了L1和L2紫茎幼苗的Pn;N2氮素水平下,各遮光处理的紫茎幼苗Pn均有所降低。在L0光强下,各氮素水平紫茎幼苗的Pn差异不明显,施氮组的Pn略低于未施氮组;在L1和L2光强下,各氮素水平下紫茎幼苗Pn差异明显,从大到小依次为N1、N0、N2;在L3光强下,各氮素水平紫茎幼苗Pn差异不明显。
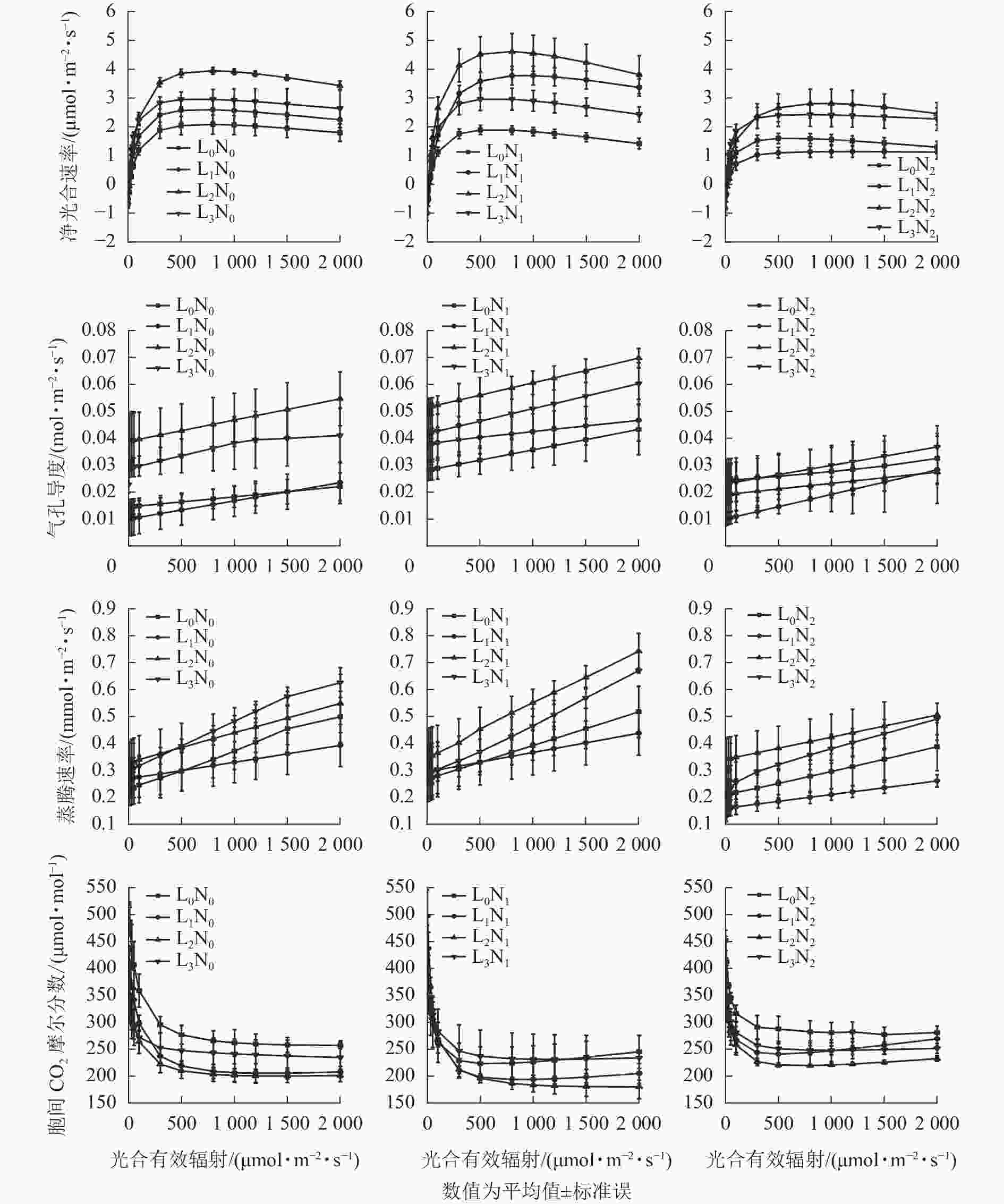
Figure 1. Photoresponse curve of S. sinensis seedlings under different light intensity and nitrogen treatments
由图1可知:随着PAR增加,紫茎幼苗Gs和Tr总体呈升高趋势。未施氮条件下,不同遮光的紫茎幼苗Gs从大到小依次为L2、L3、L0、L1,Tr从大到小依次为L3、L2、L0、L1;N1氮素水平下,紫茎幼苗Gs从大到小依次为L2、L3、L1、L0,Tr从大到小依次为L2、L3、L0、L1;N2氮素水平下,紫茎幼苗Gs均比N1低且差异较小,Tr大小顺序与N1相同。在L0光强下,各氮素水平紫茎幼苗Gs从大到小依次为N1、N0、N2,Tr从大到小依次为N0、N1、N2;在L1和L2光强下,紫茎幼苗Gs和Tr从大到小均依次为N1、N0、N2;在L3光强下,Gs从大到小依次为N1、N0、N2,N2处理的紫茎幼苗Tr明显低于N0和N1。
随着PAR增加,紫茎幼苗Ci总体呈先下降后趋于平缓的变化特点。当PAR<100 μmol·m−2·s−1时,不同光氮处理下紫茎幼苗的Ci较为接近。未施氮条件下,不同遮光的紫茎幼苗Ci从大到小依次为L2、L1、L3、L0;N1氮素水平下,L2的紫茎幼苗Ci最低,L0和L3的紫茎幼苗Ci较高;N2氮素水平下,各遮光处理的紫茎幼苗Ci均有所升高。在L0光强下,N1氮素水平的紫茎幼苗Ci最低;在L1和L2光强下,各氮素水平紫茎幼苗Ci从大到小依次为N1、N0、N2;L3光强下,各氮素水平紫茎幼苗Ci差异较小。
-
分析表1可知:紫茎幼苗的LCP变化范围为4.8~26.0 μmol·m−2·s−1,LSP变化范围为571.3~931.4 μmol·m−2·s−1,L1N1处理的紫茎幼苗光强生态幅最大。在L0光强下,不同氮素同水平紫茎幼苗的Rd、Pmax、LSP和LCP从大到小依次为N0、N1、N2,均无显著差异(P>0.05),N2处理的LCP相比N0降低了52.50%,L0处理的AQY在各组处理中较小,最小值出现在N0。在L1光强下,紫茎幼苗的Rd和LSP从大到小依次为N1、N2、N0,N1处理的Rd和LSP在所有处理中达最大,Pmax和AQY从大到小依次为N1、N0、N2,N1处理的Pmax相比N0增加了45.21%,N0处理的LCP最低且显著低于N2 (P<0.05)。在L2光强下,N0处理的紫茎幼苗Rd最高,不同氮素水平紫茎幼苗的Pmax从大到小依次为N1、N0、N2,N1处理的Pmax在所有遮光处理中达最大,LSP和AQY从大到小依次为N0、N1、N2,N2处理的LCP最小。在L3光强下,紫茎幼苗的Rd和AQY从大到小均为N2、N1、N0,N1处理的Pmax最大,LSP从大到小依次为N0、N2、N1,N0处理的LCP各组处理中最小。
处理 Rd/(μmol·m−2·s−1) Pmax/(μmol·m−2·s−1) LSP/(μmol·m−2·s−1) LCP/(μmol·m−2·s−1) AQY/(μmol·mol−1) L0N0 0.69±0.08 ab 2.09±0.03 def 791.4±79.6 ab 20.0±0.7 abc 0.027±0.003 b L0N1 0.55±0.09 ab 1.92±0.15 def 717.7±103.1 ab 19.1±4.4 abc 0.031±0.006 b L0N2 0.34±0.12 b 1.54±0.17 ef 638.2±63.0 ab 9.5±2.2 bc 0.032±0.007 b L1N0 0.51±.0.09 b 2.61±0.42 cde 741.2±78.9 ab 10.5±1.3 bc 0.036±0.007 ab L1N1 1.00±.0.27 a 3.79±0.32 abc 931.4±66.3 a 23.6±5.6 ab 0.047±0.017 ab L1N2 0.72±0.35 ab 1.12±0.06 f 877.2±92.0 ab 26.0±17.3 a 0.034±0.014 ab L2N0 0.62±0.21 ab 3.95±0.12 ab 797.1±70.8 ab 11.9±4.4 abc 0.043±0.007 ab L2N1 0.52±0.13 ab 4.63±0.65 a 758.7±101.5 ab 8.3±2.0 bc 0.041±0.002 ab L2N2 0.52±0.11 ab 2.81±0.43 bcd 571.3±49.6 b 11.3±2.7 abc 0.032±0.002 b L3N0 0.30±0.08 b 2.93±0.26 bcd 778.0±185.8 ab 4.8±1.3 c 0.043±0.002 ab L3N1 0.49±0.15 b 3.01±0.18 bcd 699.4±98.1 ab 9.6±4.2 bc 0.048±0.005 ab L3N2 0.55±0.05 ab 2.56±0.48 cde 733.6±101.4 ab 8.3±1.3 bc 0.061±0.019 a 说明:数值为平均值±标准误,同一参数不同小写字母表示不同处理间差异显著(P<0.05) Table 1. Comparison of photosynthetic parameters of S. sinensis seedlings under different light intensity and nitrogen treatments
-
由表2可知:光强对紫茎幼苗Pmax的影响极显著(P<0.001),对LCP和AQY的影响显著(P<0.05),对Rd和LSP的影响不显著(P>0.05);光强对紫茎幼苗光合参数的影响均高于氮素;氮素对紫茎幼苗Pmax的影响极显著(P<0.001);光强和氮素的交互作用对紫茎幼苗光合参数的影响均不显著(P>0.05)。
参数 光强 氮素 光强×氮素 Rd 2.028 ns 0.759 ns 1.783 ns Pmax 13.462 *** 11.209 *** 1.948 ns LSP 1.362 ns 0.642 ns 0.985 ns LCP 4.105 * 0.559 ns 1.943 ns AQY 3.133 * 0.283 ns 0.596 ns 说明:*、**、***、ns分别表示影响效应的显著性为P< 0.05、P<0.01、P<0.001、P>0.05 Table 2. Effect of light intensity, nitrogen and their interaction on photosynthetic parameters of S. sinensis seedlings
-
由表3可知:未施氮条件下,紫茎幼苗Chl a、Chl b、Chl和Car的质量分数均随着光强减弱而增加。施氮明显引起紫茎幼苗的Chl a/b和Car/Chl下降。在L0光强下,紫茎幼苗Chl a、Chl b和Chl从大到小依次为N1、N2、N0,Car的质量分数随着施氮增加而增大,N1和N2处理的Car的质量分数差异不显著(P>0.05),N1处理的Chl a/b和Car/Chl最小。在L1光强下,紫茎幼苗Chl a、Chl b和Chl的质量分数随着氮素增加而增加,且差异显著(P<0.05);Car的质量分数随着氮素增加先增多后减少,N0处理的Car的质量分数显著低于N1 (P<0.05),N2处理的Chl a/b和Car/Chl最小。在L2光强下,紫茎幼苗Chl a、Chl b和Chl的质量分数随着氮素增加而增加,Car的质量分数从大到小依次为N1、N0、N2,N2的Chl a/b和Car/Chl最小。在L3光强下,紫茎幼苗Chl a和Car的质量分数从大到小依次为N0、N1、N2,Chl b和Chl的质量分数随着氮素增加而增加,N2处理的Chl a/b和Car/Chl最小。
处理 Chl a/(mg·g−1) Chl b/(mg·g−1) Chl/(mg·g−1) Car/(mg·g−1) Chl a/b Car/Chl L0N0 0.59±0.04 f 0.26±0.02 g 0.85±0.05 i 0.17±0.01 f 2.28±0.13 b 0.20±0.00 a L0N1 1.88±0.04 b 1.15±0.13 c 3.03±0.15 d 0.46±0.05 a 1.72±0.10 d 0.16±0.02 b L0N2 1.74±0.03 c 0.93±0.04 d 2.66±0.06 e 0.47±0.01 a 1.89±0.06 c 0.18±0.00 b L1N0 1.07±0.04 e 0.43±0.02 f 1.50±0.06 h 0.26±0.01 e 2.50±0.04 a 0.17±0.00 b L1N1 1.32±0.06 d 0.70±0.05 e 2.03±0.11 g 0.36±0.03 bcd 1.90±0.04 c 0.18±0.00 b L1N2 2.09±0.01 a 1.95±0.03 b 4.03±0.02 b 0.32±0.04 cde 1.07±0.02 e 0.08±0.01 d L2N0 1.73±0.02 c 0.68±0.01 e 2.42±0.03 f 0.37±0.00 bcd 2.52±0.01 a 0.15±0.00 b L2N1 2.09±0.01 a 1.25±0.02 c 3.34±0.03 c 0.43±0.03 ab 1.68±0.02 d 0.13±0.01 c L2N2 2.12±0.02 a 1.92±0.05 b 4.03±0.04 b 0.29±0.02 de 1.22±0.03 e 0.07±0.01 d L3N0 2.17±0.02 a 1.11±0.06 c 3.28±0.08 c 0.40±0.01 abc 2.00±0.08 c 0.12±0.01 c L3N1 2.10±0.01 a 2.00±0.07 b 4.10±0.06 b 0.25±0.04 e 1.06±0.04 e 0.06±0.01 d L3N2 1.77±0.02 c 3.19±0.07 a 4.95±0.05 a 0.05±0.01 g 0.56±0.02 f 0.01±0.00 e 说明:数值为平均值±标准误,同一参数不同小写字母表示不同处理间差异显著(P<0.05) Table 3. Content and proportion of photosynthetic pigment of S.sinensis seedlings under different light intensity and nitrogen treatments
-
由表4可知:光强、氮素及其交互作用对紫茎幼苗的Chl a、Chl b、Chl和Car的质量分数和Chl a/b、Car/Chl的影响均达到极显著水平(P<0.001),氮素对紫茎幼苗Chl b、Chl和Chl a/b的影响均大于光强。
参数 光强 氮素 光强×氮素 Chl a 316.423*** 346.477*** 184.931*** Chl b 314.398*** 605.842*** 48.029*** Chl 434.234*** 726.104*** 54.300*** Car 15.993*** 12.818*** 26.129*** Chl a/b 89.690*** 373.330*** 21.619*** Car/Chl 113.570*** 104.247*** 12.472*** 说明:***表示影响效应的显著性为P<0.001 Table 4. Effect of light intensity, nitrogen and their interaction on photosynthetic pigment of S. sinensis seedlings
-
由表5可知:无论施氮与否,紫茎幼苗的Fo和Fm均随着光强减弱而升高,Fv/Fo、Fv/Fm和PIABS随着光强减弱呈先升高后降低的变化特征,全光下紫茎幼苗Fv/Fm较低且变化范围为0.526~0.633,PIABS也较小且变化范围为0.436~0.735,由此表明全光下紫茎光抑制出现了现象较为严重。施氮时紫茎幼苗Fo较低,说明施氮使PSⅡ受体侧接受电子的能力增强。L0光强下紫茎幼苗叶片的Fo、Fm、Fv/Fo和Fv/Fm从大到小依次为N0、N2、N1,PIABS从大到小依次为N2、N0、N1。L1光强下,紫茎幼苗的Fo在N2最低,Fm在N0最高,Fv/Fo、Fv/Fm和PIABS从大到小依次为N2、N0、N1,N2处理的PIABS为所有处理中最大。L2光强下,紫茎幼苗的Fo在N1最小,Fm在N0最大,Fv/Fo和Fv/Fm从大到小依次为N0、N2、N1,且均无显著差异(P>0.05),N0处理的Fv/Fo和Fv/Fm均为所有处理中最大,Fv/Fm为0.720,说明紫茎幼苗受到的胁迫最弱,PIABS在N1处理中出现最大值。L3光强下,紫茎幼苗的Fo在N2处理达最小值,Fm在N1处理达最大值,Fv/Fo、Fv/Fm和PIABS从大到小依次为N2、N1、N0。
处理 Fo Fm Fv/Fo Fv/Fm PIABS L0N0 396.0±14.1 d 1081.7±29.7 fg 1.75±0.10 d 0.633±0.014 cd 0.544±0.103 f L0N1 378.0±5.9 d 816.9±50.2 h 1.15±0.11 e 0.526±0.024 e 0.436±0.077 f L0N2 389.3±9.4 d 1013.9±92.0 g 1.58±0.20 d 0.595±0.031 d 0.735±0.182 f L1N0 458.1±8.8 c 1396.7±22.8 d 2.06±0.08 c 0.671±0.009 bc 0.783±0.110 ef L1N1 458.8±9.8 c 1193.9±66.0 ef 1.61±0.14 d 0.606±0.025 d 0.644±0.105 f L1N2 381.9±8.7 d 1298.3±33.0 de 2.40±0.05 ab 0.706±0.004 ab 2.014±0.225 a L2N0 503.7±7.4 b 1798.9±22.0 ab 2.58±0.06 a 0.720±0.005 a 1.381±0.121 cd L2N1 461.2±6.0 c 1627.2±24.6 c 2.53±0.04 a 0.716±0.003 ab 1.800±0.103 ab L2N2 469.2±13.0 c 1673.2±41.7 bc 2.57±0.06 a 0.719±0.004 a 1.686±0.152 abc L3N0 565.3±8.5 a 1786.1±47.9 ab 2.16±0.05 bc 0.682±0.006 ab 1.102±0.052 de L3N1 543.0±12.6 a 1822.4±43.5 a 2.36±0.01 ab 0.702±0.001 ab 1.437±0.063 bcd L3N2 503.2±8.0 b 1711.2±32.4 abc 2.40±0.03 ab 0.706±0.002 ab 1.525±0.069 bc 说明:数值为平均值±标准误,同一参数不同小写字母表示不同处理间差异显著(P<0.05) Table 5. Chlorophyll fluorescence induction parameters of S. sinensis seedlings under different light intensity and nitrogen treatments
-
由表6可知:光强、氮素对紫茎幼苗叶绿素荧光诱导参数的影响均达极显著水平(P<0.001);光强对紫茎幼苗叶绿素荧光参数的影响均大于氮素。光强和氮素的交互作用对紫茎幼苗Fm的影响显著(P<0.05),对Fo、Fv/Fo、Fv/Fm和PIABS的影响极显著(P<0.001)。
参数 光强 氮素 光强×氮素 Fo 130.311*** 21.499*** 5.391*** Fm 194.315*** 10.694*** 2.664* Fv/Fo 73.114*** 13.156*** 6.197*** Fv/Fm 49.274*** 10.960*** 5.284*** PIABS 39.101*** 20.694*** 8.134*** 说明:*和***分别表示影响效应的显著性为P<0.05和P< 0.001 Table 6. F-value and significance analysis of effect of light intensity, nitrogen and their interaction on chlorophyll fluorescence induction parameters of S. sinensis seedlings
-
大部分研究认为光照过强对植物叶片光系统造成损伤,光合能力降低[19-20];氮素供应过量反而会导致叶片碳同化能力降低,最终植株生长不良[21-22]。本研究中,L2N1的紫茎幼苗Pn、Gs和Tr较高,说明中度遮光和低氮处理后,紫茎幼苗的光合机构运转良好,光合能力较强,研究结果与闽楠Phoebe bournei[7]、高山杜鹃Rhododendron lapponicum等相似[23]。
光饱和点(LSP)和光补偿点(LCP)可以反映植物的需光性和需光量[24],进而表征植物光强生态幅的变化范围。本研究中,紫茎幼苗的LSP变化范围为571.3~931.4 μmol·m−2·s−1,LCP变化范围为4.8~26.0 μmol·m−2·s−1,表明紫茎幼苗具有一定耐荫性。已有研究证实,耐荫性较强的植物Pmax随着遮光程度增加仅出现轻微降低[25-27],本研究紫茎幼苗经遮光处理紫茎幼苗的Pmax高于全光对照,而且通过降低LCP提高了净光合速率,同时AQY的升高也说明紫茎幼苗具有较强的利用弱光能力,有助于紫茎幼苗适应森林群落下层遮蔽的生境。
-
在弱光环境下,植物叶片的光合色素质量分数趋于增加,在强光环境下光合色素质量分数出现下降[28];施氮能够促进光合色素的合成并提高植物的光合能力,但高水平的氮素起到相反作用[29]。本研究中,紫茎幼苗的光合色素质量分数和比值受光强和氮素的影响均为极显著(P<0.001)。与闽楠[7]、石灰花楸Sorbus folgneri[30]、风吹楠Horsfieldia glabra[31]等植物一样,紫茎幼苗叶片Chl质量分数和Car质量分数经过一段时间的遮光生长后显著增加,Car/Chl下降,说明紫茎幼苗通过提高叶绿素的相对含量来适应弱光生境,而Car质量分数提高增加有助于对蓝紫光的吸收,Chl a/b下降,推测在弱光下需要更多Chl b维持光能的捕获传递。通常,阳性植物叶片的Chl a/b约3.0,阴性植物叶片的Chl a/b约2.3[32],本研究中未施氮紫茎幼苗叶片的Chl a/b为2.0~2.5,表明紫茎在幼苗阶段的需光性更偏向于阴性树种,具有较强的弱光适应性。
施氮提高紫茎幼苗叶片Chl质量分数,尤其是Chl b质量分数增加最为显著。因此,施氮增强了叶片对光能的捕获率。本研究结果与黄檗Phellodendron amurense幼苗[33]相同,紫茎幼苗Car质量分数随着施氮的增加先升高后降低,类胡萝卜素可以通过吸收过剩光能保护紫茎幼苗的光合器官。
-
研究表明:强光会破坏植物光合机构,出现光抑制现象,甚至会诱导产生有害物质单线态氧破坏光合反应,Fv/Fm因强光胁迫明显下降被视为植物产生光抑制的敏感指标[34],PSⅡ性能指数PIABS相比Fv/Fm更能准确地反映植物光合机构对环境胁迫的响应[18]。
本研究中,全光下紫茎幼苗的叶绿素荧光强度最弱,Fv/Fm明显下降且PIABS也较小,紫茎幼苗的荧光强度随着光强的减弱而上升,Fv/Fm和PIABS随着光强减弱先升高后降低,均说明紫茎幼苗在全光下出现光抑制,相反经遮光处理具有更大的光合潜能,适度遮光可以提高PSⅡ反应中心开放度,轻度和中度遮光改善了紫茎幼苗的光能吸收状态,结果PSⅡ性能指数较高。还需要指出的是,中度遮光下紫茎幼苗的Fv/Fm相较其他光强下高,但仍低于0.75~0.85,推测幼苗光合机构性能还可能受到高温、湿度等因素的制约。
施氮提高紫茎幼苗的PIABS,对Fo有一定降低作用,说明施氮提高了紫茎光合机构的光合能力,因而PSⅡ受体侧接受电子的能力增加。全光下经施氮处理后,紫茎幼苗Fv/Fm明显降低,推测施氮加剧了全光下紫茎幼苗的胁迫程度。
-
野外调查中观察到紫茎多散生于常绿落叶阔叶林和沟谷地带,在紫茎群落下层几乎未见实生幼苗;当垂直带谱更替为常绿阔叶林或落叶阔叶林,群落内的紫茎植株更为稀少。紫茎群落外缘的土壤裸露地段可以看到绝大部分的紫茎实生幼苗,此地段侧方受光且土壤贫瘠,此时幼苗存活率成为决定紫茎天然更新能力的关键问题。可见,野外幼苗幼树的就地保护工作亟待加强。阔叶落叶林在春夏期间郁闭度较高,可在群落竞争评估基础上对密林进行适当修枝或疏伐,形成林窗以促进紫茎幼树生长。对生长林缘的幼苗进行适当遮光和施氮,实现光强和氮素的供需平衡。
-
光强、氮素及其交互作用对紫茎幼苗的光合色素质量分数和叶绿素荧光参数的影响达到显著水平(P<0.05)。紫茎在幼苗阶段对弱光有较强适应性,随着遮光程度的增加,光合色素质量分数和叶绿素荧光强度均提高,光合能力先增强后减弱,在全光下出现典型的光抑制;施氮显著提高了紫茎幼苗光合色素质量分数,低氮对紫茎幼苗光合能力的提升作用较显著,高氮的促进效应降低甚至产生胁迫抑制。本研究中,经中度遮光和低氮处理后,紫茎幼苗的光合参数处于最适宜范围。
Effects of light intensity and nitrogen treatments on photosynthetic characteristics of Stewartia sinensis seedlings
doi: 10.11833/j.issn.2095-0756.20210771
- Received Date: 2021-11-26
- Accepted Date: 2022-03-29
- Rev Recd Date: 2022-03-29
- Available Online: 2022-11-21
- Publish Date: 2022-12-20
-
Key words:
- Stewartia sinensis /
- light intensity /
- nitrogen /
- gouble factor control experiment /
- light response /
- photosynthetic pigment /
- chlorophyll fluorescence
Abstract:
| Citation: | ZHANG Tingyu, ZHANG Mingru, LI Qingxiang, et al. Effects of light intensity and nitrogen treatments on photosynthetic characteristics of Stewartia sinensis seedlings[J]. Journal of Zhejiang A&F University, 2022, 39(6): 1247-1256. DOI: 10.11833/j.issn.2095-0756.20210771

|
 DownLoad:
DownLoad: