






-
刺五加Eleutherococcus senticosus是五加科Araliaceae五加属Eleutherococcus植物,主要分布于中国的黑龙江、吉林、辽宁、河北和山西等地区[1],生长在海拔100~2 000 m的森林或者灌丛中。刺五加用途多样,其叶片翠绿富有光泽,嫩叶可以鲜食;秋季果实颜色由绿色渐变为紫红色,物候期色彩变化显著,季相景观层次丰富。此外,刺五加根或茎皆可入药,具备益气健脾、补肾安神之功效,作为中草药被收录于《中华人民共和国药典》。刺五加兼具观赏、食用和药用价值,在园林绿化中具有广阔的应用前景。
刺五加主要化学成分为酚苷类化合物、多糖和黄酮,还含有香豆素、脂肪酸、挥发油、氨基酸及微量元素等[2]。自2000版《中华人民共和国药典》开始,其药用部位由根及根茎变为根、根茎及茎。相关研究表明:刺五加根及根茎具有抗癌、抗血凝、“适应原样”等作用,刺五加叶中皂苷则具有降血糖、增强免疫、抗疲劳等作用[3]。三萜皂苷类是刺五加的主要活性成分之一,目前已经从其根和叶中分离得到了多种三萜皂苷类化合物[4−8]。随着组学技术、合成生物学及细胞工程等技术的快速发展,刺五加三萜皂苷(triterpenoid saponins)的生物合成分子机制研究也逐步深入,多个三萜皂苷合成酶基因已被成功克隆,其蛋白结构、功能、作用机制等也得到了广泛解析。此外,刺五加三萜皂苷生物合成的分子调控机制也同样有所进展。本文旨在对刺五加三萜皂苷生物合成以及调控的研究进展进行总结与归纳,以期为今后刺五加的多用途深入开发利用提供理论依据。
-
三萜皂苷是由三萜皂苷元和1个或多个糖基和(或)其他化学基团缩合而形成的一系列结构多样的天然化合物[9],主要分布在五加科、石竹科Caryophyllaceae、桔梗科Campanulaceae、毛茛科Ranunculaceae等植物中[10]。三萜皂苷元骨架由30个碳原子构成,常见类型为四环三萜和五环三萜,其中达玛烷型(dammarane)和齐墩果烷型(oleanane)是最常见的三萜母核类型[11]。刺五加含有丰富的三萜骨架,其四环三萜骨架有环阿屯醇,五环三萜骨架则包括齐墩果烷型、去甲齐墩果烷型、羽扇豆烷型、3,4-seco-羽扇豆烷型[12]。以这些不同类型的三萜骨架作为基础,进而形成多种刺五加三萜皂苷[13]。刺五加三萜皂苷一般在齐墩果酸(oleanolic acid) C3位先O-连接1个α-阿拉伯糖基(α-arabinopyrannosyl),然后1→2连接1个α-鼠李糖基(α-rhamnopyranosyl)或葡萄糖基。在C28位首先通过O-糖苷键连接1个β-吡喃葡萄糖基(β-glucopyranosyl,β-Glu),该糖基再以1→6糖苷键与另一分子β-Glu相连,形成β-Glu-(1→6)-β-Glu二糖结构;随后依次通过1→4糖苷键连接α-吡喃鼠李糖基(α-rhamnopyranosyl,α-Rha),并以1→6糖苷键连接乙酰基(acetyl)[12−15]。
三萜皂苷是一类具有多种生物活性的天然产物,简单的三萜是植物表面蜡质和特化膜的组成成分,同时也是植物潜在的信号分子[14];而经糖基化修饰的三萜皂苷则为植物在逆境或胁迫中的生存提供保护[9]。它们不仅参与植物的通信、防御和感觉调控,还具有抗炎、抗癌、抗菌、抗病毒等多种生物活性[16−18]。LI等[18]和GE等 [19]发现:刺五加三萜皂苷会对胰脂肪酶(pancreatic lipase,PL)显示出抑制活性,其化学结构中C28位的游离羧酸基团(free carboxylic acid group,—COOH)能够增强PL的抑制作用。此外,刺五加三萜皂苷也是开发抗阿尔茨海默病药物的先导化合物之一,进一步凸显刺五加三萜皂苷的药用潜力。因此,深入研究并解析刺五加三萜皂苷的代谢途径,对完善植物次生代谢理论体系以及合理利用与开发刺五加药用资源具有重要意义。
-
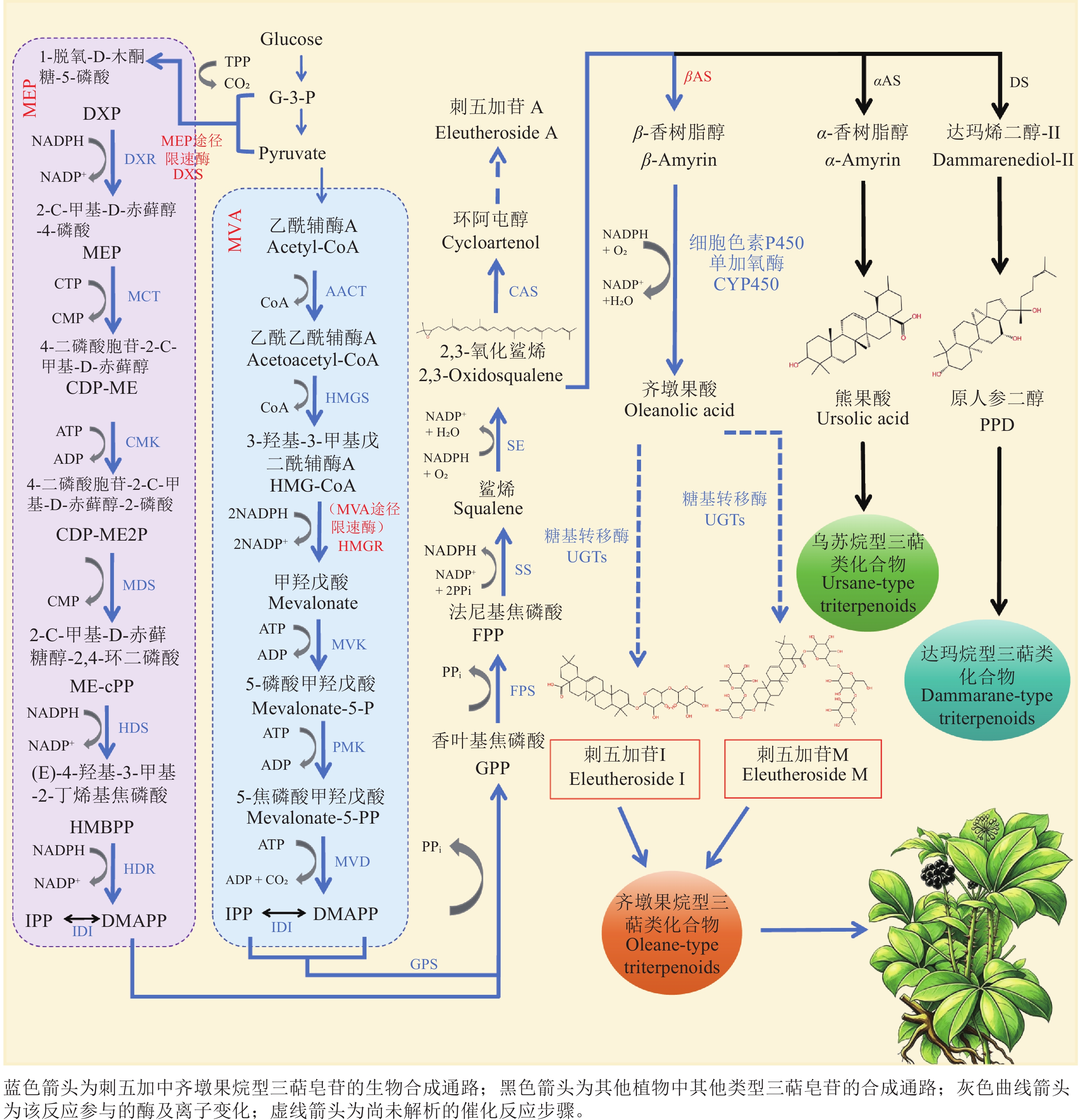
刺五加三萜皂苷生物合成的核心起始物质为异戊烯基焦磷酸(isopentenyl diphosphate,IPP,C5)与二甲基烯丙基焦磷酸(dimethylallyl diphosphate,DMAPP,C5)。这2种物质通过2条独立且协同的途径在植物体内合成,即细胞质中的甲羟戊酸(mevalonate,MVA)途径和质体中的2-C-甲基-D-赤藓醇-4-磷酸(2-C-methl-D-erythritol-4-phospate,MEP)途径(图1)。MVA途径以乙酰辅酶A (acetyl-CoA)为起始底物,经多步酶促反应逐步生成IPP,各关键基因及其编码酶在途径中发挥精准调控作用。MEP途径以丙酮酸(pyruvic acid)和甘油醛-3-磷酸(glyceraldehyde-3-phosphate,GAP)为起始底物,经过7步连续酶促反应合成IPP[20],同时伴随少量DMAPP生成,各关键酶的功能与调控特性共同决定途径的运行效率。MVA途径主要合成倍半萜、三萜、甾醇等次生代谢产物,MEP途径主要合成质体中的单萜、二萜、类胡萝卜素等次生代谢产物。植物中MVA途径与MEP途径并非孤立运行,而是通过跨细胞器代谢物转运、组织发育调控、环境信号响应及代谢产物反馈等多重机制形成协同网络,精准匹配萜类化合物的合成需求[21]。
Figure 1. Biosynthetic pathway of triterpenoid saponins in E. senticosus
IPP与DMAPP生成后,需经过系列酶促缩合反应延长碳链,形成三萜皂苷合成的关键前体:首先在香叶基焦磷酸合成酶(geranylpyrophosphatesynthetase,GPS)催化下,IPP与DMAPP缩合生成香叶基焦磷酸(geranylpyropho-sphate,GPP,C10);随后GPP在法尼基焦磷酸合酶(farnesyl pyrophosphatesynthetase,FPS)作用下结合第2个IPP单元,生成法尼基焦磷酸(farnesylpyrophosphate,FPP,C15);2个FPP分子经鲨烯合成酶(squalene synthase,SS)缩合形成角鲨烯(squalene,SQ);最终角鲨烯在鲨烯氧化酶(squaleneepoxidase,SE)催化下发生环氧化反应,生成2, 3-氧化鲨烯(2, 3-oxidosqualene,2, 3-OS)[11]。
多样化的三萜骨架源于环氧鲨烯环化酶(oxidosqualene cyclases,OSCs)对2,3-OS的催化环化。不同类型的OSCs具有高度的底物立体选择性与产物特异性,能引导环化反应沿特定路径进行,从而生成结构迥异的三萜,例如分别催化形成乌苏烷型、达玛烷型或齐墩果烷型等。随后,形成的三萜骨架会进一步发生修饰。细胞色素P450单加氧酶(cytochrome P450 monooxygenase,CYP450s)通过羟基化等反应,在三萜骨架的特定位置引入含氧化官能团(如羟基)。此外,尿苷二磷酸糖基转移酶(uridine diphosphate-dependent glycosyltransferase,UGTs)会催化糖基从活化的糖供体转移至修饰后的三萜苷元上,最终生成具有不同结构和生物活性的三萜皂苷。
-
在MVA生物合成途径中,3-羟基-3-甲基戊二酰辅酶A还原酶(3-hydroxy-3-methylglutaryl-CoA reductase,HMGR,EC1.1.1.34)是首个关键限速酶,也是植物代谢中的重要调控位点。研究发现:不同产地的刺五加中,HMGR基因的表达量与总皂苷含量存在显著差异,且两者呈极显著正相关[22]。同时,HMGR基因在刺五加的所有器官中均有表达,且其表达水平在开花期最高,在叶片衰老期最低,相差13.4倍[23]。此外,HMGR基因还积极响应茉莉酸甲酯(methyl jasmonate,MeJA)信号,其从萌发期到盛花期的表达量在MeJA处理后显著提高,并且刺五加皂苷含量与HMGR基因表达水平呈极显著正相关[23]。
甲羟戊酸二磷酸脱羧酶(mevalonate diphosphate decarboxylase,MDD,EC 4.1.1.33)能催化三磷酸腺苷(adenosine triphosphate,ATP)与甲羟戊酸焦磷酸盐(mevalonate 5-pyrophosphate,MVA-PP)反应生成IPP,也是萜类化合物生物合成过程中的重要合成酶之一[24]。李志栋等[25]克隆并分析了刺五加MDD基因的启动子序列,发现该序列包含脱落酸响应元件、MeJA响应元件、胚乳表达必需的顺式作用元件、干旱胁迫响应元件、光响应元件等顺式作用元件。邢朝斌等[26]克隆了MDD基因的全长cDNA序列,发现刺五加MDD基因在不同生长阶段和器官中均有表达,且表达水平差异显著。可见,MDD可能受到激素信号和非生物胁迫的调控,并具有时空表达特异性。
FPS也是三萜皂苷合成途径中的重要限速酶之一,它催化两分子IPP和一分子DMAPP在1位和4位进行头尾缩合形成FPP。前人对刺五加FPS基因的表达模式进行了研究,结果表明:其相对表达水平与刺五加叶中的皂苷含量呈显著正相关[27]。此外,前人克隆得到亚精胺合成酶基因(spermidine synthase,SPDS),并证实了内生真菌能显著提高该基因的表达[28],为阐明内生真菌可提高刺五加三萜皂苷含量的机制及刺五加的抗逆性改良奠定了基础。研究提示:SPDS基因可能通过调控亚精胺的合成,间接影响MEP途径中关键酶的表达或活性,进而参与三萜皂苷的合成调控。
-
OSCs是三萜皂苷下游生物合成通路中的首个限速酶,同时也是三萜皂苷与甾醇两大生物合成路径的核心分支节点。该酶以C30为底物,通过精准催化质子化环化、骨架重排及去质子化等一系列反应,完成特定的环化过程,最终生成构成三萜皂苷的基础碳骨架结构。李敏等[29]对刺五加进行了全基因组测序,并预测了其三萜皂苷生物合成中的关键OSC基因,验证了EsOSC5基因的编码产物为β-香树脂醇合酶(β-amyrin synthase,βAS)。因此,EsOSC5基因被认为是促进刺五加中βAS合成和积累的关键基因,直接影响三萜皂苷的合成效率。βAS为OSCs的一员。研究人员发现:βAS基因在所有时期、器官及愈伤组织中均有表达[30−31],但表达水平存在显著差异。βAS1基因在所有器官中持续表达,而βAS2基因仅在叶片中的表达水平最高。此外,βAS1基因对MeJA处理无响应,而βAS2基因的表达水平则被显著诱导[31]。这些差异表明:2类βAS基因可能受不同的调控通路调控,共同应对刺五加在不同环境条件下的三萜皂苷合成需求。
除了上述合成酶,三萜合成通路中还涉及鲨烯合酶(squalene synthase,SS)、鲨烯环氧酶(squaleneepoxidase,SE)等[32]重要合成酶。鲨烯环氧酶基因(SE)是植物三萜皂苷生物合成下游的重要基因,其功能作用于三萜骨架构建阶段。当前研究表明:随着拷贝数的增加,刺五加中SS2和SE1的表达水平以及皂苷含量显著增加,并且三者间存在极显著正相关[33−34]。这一结果不仅证实了SS2与SE1基因在三萜皂苷合成中的核心作用,也为通过基因编辑或调控拷贝数来提高刺五加皂苷产量提供了明确的分子靶点。
-
CYP450s是三萜皂苷生物合成途径中的关键修饰酶,可以催化三萜骨架进行羟基化、羧基化、醛基化、酮基化和脱氢等反应,形成三萜皂苷的中间体[11]。吴鹏等[35]的研究率先构建了刺五加CYP85亚家族的P450基因的重组载体pET30a-P450,实现了刺五加P450基因在原核表达系统中的功能性表达。此前,其所属研究团队通过多维度表达谱分析已揭示了刺五加P450基因的表达特征[36−37]:首先经功能预测与系统进化分析,判断该基因隶属调控三萜皂苷合成的CYP85亚家族,其氨基酸序列进化树结果与传统分类一致。此外,团队对该基因在不同产地、组织器官、发育时期及MeJA处理下的表达谱检测,结果显示其呈组成型表达并具明显组织特异性:叶片丰度最高,约为幼茎(最低)的1.49倍;经MeJA诱导后,表达水平显著上调。相关性分析证实:该基因的表达水平与刺五加总皂苷含量呈极显著正相关,提示其为皂苷合成核心基因,表达强度可作为调控总皂苷积累的关键分子指标。后续研究中,宋鑫等[38]对刺五加CYP450基因(EsP450)开展家族分析,发现EsP450中占比最高的类型为CYP71基因簇,而该基因簇所属家族与已知参与三萜结构修饰的P450分布家族基本吻合[39];其团队通过基于最大似然法的系统发育分析软件(phylogenetic analysis by maximum likelihood,PAML)、进化混合效应模型(mixed effects model of evolution,MEC)、Datamonkey在线分析服务器共3种方法对EsP450进行分析,结果显示:EsP450无正选择位点,推测它们以负选择为主导,受强烈负选择约束,在长期进化中高度保守,进而保护基因核心功能;其研究所用的不同刺五加样本皂苷含量结果也与吴鹏等[37]的分析相互印证,为后续蛋白功能解析奠定了技术基础。
目前关于EsP450基因的公开研究文献较为有限,且由于CYP450家族基因功能的复杂性,其功能预测仍存在较大难度。为进一步解析EsP450基因在三萜皂苷合成中的具体功能,可从多维度开展深入探究:如可通过体外酶活实验验证候选EsP450基因的催化活性;其次,可借助酵母异源表达系统或构建刺五加CRISPR/Cas9基因敲除株系,进而深入验证体内环境中这些基因对皂苷合成的实际调控作用;同时可结合EsP450基因启动子区域的转录因子筛选,揭示该类基因表达调控的分子机制。此外,分析EsP450基因在刺五加不同组织器官、生长发育阶段的时空表达模式,以及在非生物胁迫、激素处理下的响应特征,也能为明确其功能特异性提供重要依据。
-
糖基转移酶(uridine diphosphate-dependent glycosyltransferase,UGTs,EC2.4.1.x)隶属于糖基转移酶(glycosyltransferase,GT)超家族,是介导植物天然产物糖基化的核心酶类[11]。UGTs核心功能是将尿嘧啶核苷二磷酸糖(uridine diphosphate sugar)上的活性糖基转移到植物中的小分子化合物受体上。在三萜皂苷生物合成中,催化三萜苷元糖基修饰的UGTs均归为此类。该修饰不仅是三萜皂苷结构形成的关键步骤,更直接调控其水溶性、生物活性等核心理化与功能特性。此外,UGTs介导的糖基化反应通常被视作三萜皂苷生物合成的终末步骤,亦是其结构多样性与生物活性多样性形成的决定性环节。修乐山等[40−41]为了探明刺五加中GT基因的表达特点及其对刺五加总皂苷含量的影响,分析了不同产地、生长发育时期、器官中GT基因的表达规律,及MeJA对其表达的影响。结果发现:不同产地刺五加的GT基因表达量与总皂苷含量存在显著差异,且两者呈极显著正相关,其中UGT作为GT超家族中负责三萜苷元糖基化修饰的核心功能亚家族,其表达水平与总皂苷含量的相关性最为突出。由此可初步判定:UGTs是决定刺五加中总皂苷含量的关键酶。在此基础上研究人员进一步发现:GT基因在各生长时期和器官中均有表达,但表达量具有显著差异,最大表达量出现在盛花期,此时为最低表达量(叶片衰老期)的1.81倍。在各器官中,叶片表达量最高,幼茎最低[42]。
尽管高通量测序已获得大量植物UGT序列,但当前刺五加UGT基因与总皂苷含量的关联研究,仍局限于基因表达与皂苷含量的相关性分析,缺乏直接功能证据;既未过表达验证UGT表达提升能否直接增加总皂苷含量,也未用基因敲除或沉默技术(如CRISPR-Cas9、RNA干扰)证实UGT功能缺失是否会导致皂苷合成受阻。这类仅依托于相关性的结论,无法排除两者同步变化是受转录因子、信号分子等共同上游调控因子间接驱动的可能,故UGT基因功能判定仍需直接实验支撑。破解该困境需依托技术整合与创新:一是通过蛋白质结构预测结合分子对接,明确UGT酶的活性中心、底物结合口袋等关键位点,为UGT酶功能分析提供理论基础;二是构建高通量功能验证平台(如酵母表面展示酶活筛选、烟草Nicotiana tabacum瞬时共表达路径重构),实现候选UGT基因的快速筛选与催化特性鉴定;三是可通过转录组、代谢组、蛋白质组等多组学整合,挖掘UGT的上游调控因子以及协同代谢酶基因,明确其在三萜皂苷合成路径中的作用节点。将上述技术手段与跨学科分析整合,有望突破刺五加UGT基因功能注释难题,推动刺五加UGT基因从候选基因研究迈向三萜皂苷合成生物学产业化应用。
-
植物中三萜皂苷的生物合成受基因调控,通过增加基因拷贝数或抑制竞争代谢途径基因,可以人为改变三萜生物合成途径基因的表达[43]。前人在刺五加中的研究主要围绕转录因子(transcription factor,TFs)和表观遗传修饰展开。
-
TFs (又称反式作用因子)是真核生物转录调控的核心蛋白,通过与靶基因启动子区顺式作用元件特异性结合调控转录。已知TFs涵盖84个家族,依据DNA结合结构域的特征,分为髓母细胞瘤转录因子家族(myeloblastosis transcription factor family,MYB)、碱性螺旋-环-螺旋转录因子家族(basic Helix-Loop-Helix,transcription factor family,bHLH)、碱性亮氨酸拉链转录因子家族(basic Leucine Zipper transcription factor family,bZIP)等类群,其中MYB是植物最大的转录因子家族之一,可广泛调控生长发育、次生代谢等核心生理过程。在刺五加中,分属36个家族的485条转录因子基因被挖掘出来。这些基因可能参与皂苷的合成调控,其中MYB家族成员数量最丰富,同时还包含bZIP、bHLH及WRKY等转录因子家族(WRKY domain-containing transcription factor family,WRKY)等家族的成员[44]。研究证实,MYB转录因子可通过结合皂苷合成关键酶基因的启动子区域调控基因表达,bZIP、bHLH及WRKY等家族成员也被证实参与这一调控过程[45]。转录因子与启动子的结合能力能够直接影响调控效率,如刺五加NAC (NAM、ATAF1/2、CUC2V)转录因子通过结合关键酶基因启动子发挥调控作用,阻碍两者结合从而间接抑制皂苷生物合成。这一机制为通过分子手段调控三萜皂苷含量提供了明确靶点[46−47]。据最近的研究报道,河北理工大学邢朝斌教授团队首次在刺五加中鉴定出21个远红光响应缺陷蛋白1/远红光下胚轴伸长蛋白3转录因子(far-red impaired response 1/far-red elongated hypocotyls 3 transcription factor,FAR1/FHY3)的基因,并对该基因家族进行了分析,筛选出4个可结合EsFPS启动子的转录因子。这个新发现为解析光质影响刺五加三萜皂苷次生代谢的分子调控机制提供了参考[48]。
-
刺五加中表观遗传修饰研究主要围绕DNA甲基化与组蛋白修饰的协同作用展开。目前,C5-甲基转移酶(C5-MTase)与去甲基化酶(dMTase)家族已被鉴定,发现EsMET1a的SER844-ASP845-LYS846残基为底物结合关键位点[49]。该系统与环境信号密切相关,如干旱可降低皂苷合成关键酶基因启动子甲基化水平,增强转录因子结合能力,促进皂苷积累[50];光照也通过调控表观修饰影响相关基因表达,建立起表观遗传与皂苷含量的关联。
另外,研究发现刺五加MDD基因的启动子甲基化通过多重机制调控皂苷合成[51]:EsC5-MTase (如EsMET1a)与EsdMTase催化特定胞嘧啶位点的甲基化与去甲基化,其活性平衡决定启动子甲基化状态。高甲基化阻碍转录因子结合,抑制MDD表达,降低MVA途径通量及IPP供给,从而抑制皂苷合成;干旱、适宜光照等信号可降低启动子甲基化水平,解除转录抑制,上调MDD表达,为皂苷合成提供充足前体。该机制体现了甲基转移酶家族的核心作用,揭示了环境信号通过表观修饰连接胁迫响应与代谢调节的逻辑,为定向改良刺五加皂苷含量提供了靶点与理论依据。
-
刺五加作为中国濒危珍稀保护植物,因具备突出的观赏特性、丰富的食用价值及显著的药用功效,而具有重大的保护与研究意义。以刺五加为原料的产品在国内外广受好评,加速了刺五加的综合开发和利用,刺五加野生资源遭到严重破坏,人工种植采收周期长,造成供需矛盾日益突出。在1992年出版的《中国植物红皮书——珍稀濒危植物(第1卷)》中,刺五加已被列为濒危物种。保护刺五加资源,研究其活性成分的生物合成机制迫在眉睫。
作为刺五加的核心活性成分,三萜皂苷的生物合成机制是当前植物次生代谢研究的重要方向,与植物与外部环境的相互作用,以及人类生活和发展密切相关。随着现代生物技术的进步,三萜皂苷的生物合成途径日益明晰,在关键酶编码基因、转录调控以及生物工程等领域的研究均取得了显著进展。然而,目前仍缺乏针对刺五加三萜皂苷生物合成过程的系统且深入的认知,仅有少数关键酶基因被克隆,且这些基因在三萜皂苷生物合成中的具体作用机制尚未完全阐明,仍需深入研究。因此,作者建议今后可加强以下几个方面的研究:①进一步明确刺五加三萜皂苷生物合成关键酶的功能,探索、解析该生物合成途径的未知步骤;②进一步解析MYB、bHLH等转录因子以及miRNA相关调控机制;③不同产地刺五加的皂苷含量差异与HMGR、UGT等基因表达相关,但尚未明确环境和基因型的互作效应。例如,由环境诱导的表观修饰(如DNA甲基化)是否会导致同一基因型刺五加在不同产地出现基因表达差异,尚不得而知。这限制了优质种质资源筛选与栽培区域优化的应用转化。
随着分子生物学、生物信息学及组学技术的日趋成熟,多学科交叉融合显著加速了三萜皂苷的基础研究与产业高质量发展进程。截至2025年9月,已公开的刺五加资源数据中,Genbank数据库中已收录有近100个刺五加转录组数据,但是代谢组、蛋白质组等组学数据严重匮乏,小RNA组、单细胞及空间转录组等数据空白,且功能注释不够系统全面。因此,今后研究也需要加强用大数据解析三萜皂苷生物合成及调控分子机制,构建“表观遗传—转录—转录后”完整调控网络,深化多层级调控体系解析,依托现有遗传转化体系,突破单基因调控瓶颈,探索多基因协同增效机制,挖掘优质种质资源并明确环境调控机制,为优质种质培育与栽培调控提供依据。
生物合成技术创新为三萜皂苷拓展了生产路径。通过基因工程与生物工程技术重构微生物代谢途径,可实现目标产物规模化生产。2020年11月16日Nature Communication报道,基于皂苷合成途径中糖基化环节重要细节的突破性发现,研究人员成功在酵母中构建了生产甘草皂苷(齐墩果烷型三萜皂苷)的工程化生物合成途径,为高价值产品商业化生产奠定了基础[52]。研究人员从人参Panax ginseng中鉴定特异性UDP-糖基转移酶(uridine diphosphate glycosyltransferase Pg1,UGTPg1),该酶具有高度底物特异性,可催化达玛烷型四环三萜化合物中C20位的S-构型羟基发生糖基化修饰。在酵母底盘细胞中,将UGTPg1与人工构建的原人参二醇合成途径进行共表达后,成功实现了单糖到稀有人参皂苷CK (20-O-β-D-glucopyranosyl-20(S)-protopanaxadiol)的全生物合成[53]。由此可见,未来在合成生物学领域有望实现大量获取目标代谢物,实现刺五加资源的保护和可持续利用。
刺五加三萜皂苷的生物合成机制的解析也有助于利用体外组织培养技术生产目标代谢物,整合植物组织培养、细胞培养、体细胞胚胎发生及根培养等技术,结合细胞工程与基因工程手段,筛选并构建具有三萜皂苷高效合成能力、遗传性质稳定并且培养特性优良的基因工程细胞系,为刺五加三萜皂苷的工业化生产提供新途径。但该过程涉及多个酶的协同作用和复杂的代谢调控,对于科研工作者来说仍然是一个严峻挑战。
Research progress on the molecular mechanism of biosynthesis and regulation of triterpenoid saponins in Eleutherococcus senticosus
doi: 10.11833/j.issn.2095-0756.20250477
- Received Date: 2025-09-01
- Accepted Date: 2026-04-28
- Rev Recd Date: 2026-04-23
- Available Online: 2026-07-21
- Publish Date: 2026-08-20
-
Key words:
- Eleutherococcus senticosus /
- triterpenoid saponins /
- biosynthetic enzyme /
- transcription factor
Abstract: Systematic elucidation of the biosynthetic pathways and regulatory mechanisms of triterpenoid saponins in Eleutherococcus senticosus is of great scientific significance and practical application value for the efficient exploitation and utilization of this medicinal plant resource. In recent years, research on triterpenoid saponins in E. senticosus has focused primarily on 3 interrelated aspects: the chemical composition and biological activities of triterpenoid saponins, the functional characteristics of key biosynthetic enzymes and their encoding genes, and the regulatory mechanism at the levels of transcription factors and epigenetic modifications. Key findings to date indicate that the triterpenoid saponins in E. senticosus are dominated by oleanane-type and lupane-type structures, and their biological activities are closely associated with glycosylation modifications and their aglycone structures. These saponins are biosynthesized from isopentenyl diphosphate (IPP) and dimethylallyl diphosphate (DMAPP) − the core precursors produced by both the cytosolic mevalonate (MVA) pathway and the plastidial 2-C-methyl-D-erythritol 4-phosphate (MEP) pathway. Through the condensation and epoxidation reactions catalyzed by geranyl pyrophosphate synthase (GPPS), farnesyl pyrophosphate synthase (FPPS), squalene synthase (SS), and squalene epoxidase (SE), 2,3-oxidosqualene (2,3-OS) is obtained, which is then cyclized by oxidosqualene cyclases (OSCs) to form the skeleton. After hydroxylation by cytochrome P450 monooxygenases (CYP450s) and glycosylation by uridine diphosphate-dependent glycosyltransferases (UGTs), triterpenoid saponins with diverse structures and distinct biological activities are generated. Studies on the regulatory network of triterpenoid saponin biosynthesis in E. senticosus mainly focus on gene regulation. On the one hand, 485 transcription factor genes belonging to 36 families have been identified, and the expression regulation of these transcription factors on the promoters of key enzyme genes for saponin biosynthesis has been characterized. On the other hand, synergistic epigenetic regulatory mechanisms including DNA methylation and histone modifications have been found to modulate gene expression by balancing the methylation/demethylation status of promoters of pivotal enzyme genes such as mevalonate diphosphate decarboxylase (MDD, EC 4.1.1.33). Moreover, environmental signals such as drought and light have been shown to affect saponin biosynthesis by regulating epigenetic modifications (including promoter methylation status), providing potential targets and a theoretical basis for molecular modulation of triterpenoid saponin accumulation in E. senticosus. Future research on E. senticosus triterpenoid saponins should focus on the dissection of multi-level regulatory networks, exploration of polygenic synergistic effects, and identification of elite germplasm resources. Based on multi-omics analysis and synthetic biology techniques, solid theoretical support and key technical references should be provided for the exploitation and utilization of E. senticosus, as well as the innovative breeding of high-quality germplasm. [Ch, 1 fig. 53 ref.]
| Citation: | WU Chenwei, XIA Siyu, DU Yuqing, et al. Research progress on the molecular mechanism of biosynthesis and regulation of triterpenoid saponins in Eleutherococcus senticosus[J]. Journal of Zhejiang A&F University, 2026, 43(4): 891−901 doi: 10.11833/j.issn.2095-0756.20250477

|
 DownLoad:
DownLoad: