






-
氧化亚氮(N2O)是一种强效温室气体,具有极强的臭氧层破坏能力,其百年温室效应潜势为二氧化碳的310倍,生命周期长达114 a[1−3]。农林业活动贡献了全球60%到70%的氧化亚氮排放量[4−5],其中氮肥过量施用是主要驱动因素[6]。而中国氮肥利用率不足35%[7−9],远低于全球平均水平,导致大量活性氮通过硝化和反硝化作用转化为氧化亚氮[10−11]。因此,减少氧化亚氮排放已成为亟待解决的科学和技术难题。硅肥的施用是减少氧化亚氮排放的一种有效管理措施。已有研究表明:水稻Oryza sativa生长期施用钢渣[氧化硅(SiO2)为40.7%,氧化铁(Fe2O3)为4.8%]减少了98.0%氧化亚氮的排放量[10]。稻田土壤施用偏硅酸钠和矿渣基硅酸盐肥料(均含有SiO2和Fe2O3)分别降低氧化亚氮排放通量66.6%和40.3%,且均能降低反硝化潜势[11]。然而,目前为止,硅肥施用抑制土壤氧化亚氮排放的具体机制仍不明确。硅肥可能通过多种机制来减少土壤氧化亚氮排放,包括降低底物浓度(铵态氮、硝态氮)[12−14],改变土壤水分和氧气含量[15−19],降低硝化和反硝化作用相关基因的丰度[20−22]。例如,钢渣硅钙肥能够吸附并固定土壤中的游离铵态氮和硝态氮,影响土壤水分和铁离子浓度,从而减少氧化亚氮的排放[23]。矿渣基硅酸盐肥料显著降低了与反硝化作用相关的nirS和nirK基因丰度,使土壤氧化亚氮排放通量降低40.34%[11]。然而,这些研究大多集中在长期淹水环境中,且硅肥通常含有氧化铁等影响氧化亚氮排放的因素。因此,有必要研究单独添加硅对旱地土壤氧化亚氮排放的实际影响。
毛竹Phyllostachys edulis是一种富硅型植物[24],其林地每年从土壤中吸收大量硅素养分,导致硅素供应失衡问题日益严重[25]。施用硅肥可有效缓解毛竹林硅元素供需矛盾的现实需求[26]。由于毛竹林具有较高的经济效益,为了追求高产,部分竹林每年化肥的施用量达450.0 kg·hm−2,这一数值是谷类作物氮肥投入量的2倍。集约经营毛竹林氧化亚氮排放量达10.1 kg·hm−2·a−1,明显高于中国农业土壤氧化亚氮排放的最高水平(5.0 kg·hm−2·a−1),并且比常绿阔叶林和粗放经营毛竹林的排放量分别高出131%和149%[27−28]。为此,本研究通过分析不同硅氮添加条件下土壤氧化亚氮排放速率动态、氮素形态转化过程及关键酶活性的变化特征,探讨硅氮耦合作用对毛竹林土壤氧化亚氮排放的影响及其内在调控机制,以期为缓解毛竹林土壤氧化亚氮排放和优化氮肥管理策略提供科学支持。
-
研究区位于浙江省杭州市临安区青山镇(30°19′N,119°22′E),该区年平均气温为15.8 ℃,年平均降水量为1 613.9 mm,无霜期为234.0 d,土壤类型为红壤土类黄红壤亚类,研究样地为人工经营的毛竹林,林龄为26 a,该林分已进入成熟稳定期,其乔木层以毛竹为绝对优势种,林分结构单一,平均胸径为9.9 cm,密度为2 900株·hm−2。经营目标以材用为主,兼顾生态效益,年施尿素为200 kg·hm−2、氯化钾为70 kg·hm−2、过磷酸钙为60 kg·hm−2,人工清除林下植被。
2024年9月,在研究样地内随机布设10个采样点,用土钻采集0~10 cm表层新鲜土壤,将各采样点土壤样品充分混匀,合并为一份混合待测样品。土样带回实验室,除去新鲜土样中的石头、根系、植物残体等杂物,过2 mm筛后,将所获土样分为两份:一份用于土壤初始理化性质测定;另一份置于4 ℃冷藏保存,以备培养实验使用。研究区土壤初始理化性质:铵态氮为(11.34±0.74) mg·kg−1,硝态氮为(29.41±2.11) mg·kg−1,pH为3.33±0.10,脲酶活性为(15.72±0.06) μg·kg−1·h−1,硝酸还原酶活性为(4.02±0.50) mg·g−1·d−1,亚硝酸还原酶活性为(5.68±0.40) mg·g−1·d−1。
-
本研究采用双因素完全随机试验设计,设置硅(Si)和氮(N) 2个因素。硅因素设2个水平:S0 (不施硅)和S1 (施加0.66 g·kg−1九水偏硅酸钠,折合纯硅施用量为315 kg·hm−2);氮因素设3个水平:N0 (无氮,不施尿素)、N1 (中氮,施加0.2 g·kg−1尿素,折合尿素施用量为300 kg·hm−2)、N2 (高氮,施加0.4 g·kg−1尿素,折合尿素施用量为600 kg·hm−2)。2个因素完全交叉形成6个处理组合(S0N0、S0N1、S0N2、S1N0、S1N1、S1N2),每个处理重复3次。
培养实验操作如下:培养箱温度设定为25 ℃,称取相当于20 g烘干土的4 ℃储存新鲜土样于培养瓶中,使瓶内土面平整。按处理设计配置对应质量分数的尿素和九水偏硅酸钠溶液,用移液管均匀滴入1 mL对应溶液(S0N0处理仅添加等量的去离子水)。随后补充去离子水使土壤含水量保持在60%田间最大持水量,用保鲜膜封口并均匀扎孔,置于25 ℃黑暗条件下培养。参考蒋文婷等[29]对毛竹林土壤氧化亚氮排放的研究,本研究设置培养周期为30 d,并分别在第0、1、3、7、15、30天进行破坏性采样。每次采样时,先去除培养瓶保鲜膜并置于室内进行气体置换30 min,随后抽取20 mL瓶内气体作为第1次采气样品,密封培养4 h后,再次抽取20 mL瓶内气体作为第2次采气样品,2次气体样品均用气相色谱仪测定氧化亚氮质量分数,土壤样品用于铵态氮、硝态氮和pH测定,其中第0、7、30天的样品额外测定脲酶、硝酸还原酶、亚硝酸还原酶活性,分别代表培养起始期、氧化亚氮排放高峰期和培养结束期。培养期内隔2 d称取培养瓶质量,补充灭菌水以维持土壤水分。试验总计设置6个培养时间×6个处理×3个重复,共108个培养瓶。
-
土壤铵态氮采用靛酚蓝比色法测定,土壤硝态氮采用酚二磺酸比色法测定,土壤pH采用去离子水浸提法,水土比(质量比)为2.5∶1.0。
-
土壤脲酶采用苯酚钠-次氯酸钠比色法测定[30]。硝酸还原酶采用酚二磺酸比色法测定[31]。亚硝酸还原酶采用α-萘胺比色法[32]。
-
氧化亚氮排放通量计算公式[33]如下:
其中:F为氧化亚氮排放通量(μg·kg−1·h−1);ρ为氧化亚氮气体密度(1.25 kg·m−3);V为培养瓶中气体的有效空间(m3);W为置于培养瓶内的烘干土质量(kg);∆C为2次采样时间间隔的气体浓度差(μL·L−1);∆t为2次采样的时间间隔(h);T为培养时的温度(℃)。
氧化亚氮累积排放量计算公式[34]为:
其中:M为氧化亚氮累积排放量(μg·kg−1);Fi+Fi+1为同一采样点连续2次气体排放通量之和(μg·kg−1·h−1);i表示第i次采集气体样品;ti+1–ti为连续2次采集样品所隔天数(d)。
采用SPSS 27对数据进行统计分析。数据经正态性和方差齐性检验后,用双因素方差分析评估硅、氮及其交互作用,用Duncan法进行多重比较(α=0.05)。时间序列数据采用重复测量方差分析(Greenhouse-Geisser校正)。Spearman相关分析用于评估氧化亚氮排放与各指标的关系,多重比较采用Bonferroni校正,绘图使用Origin 2024。
-
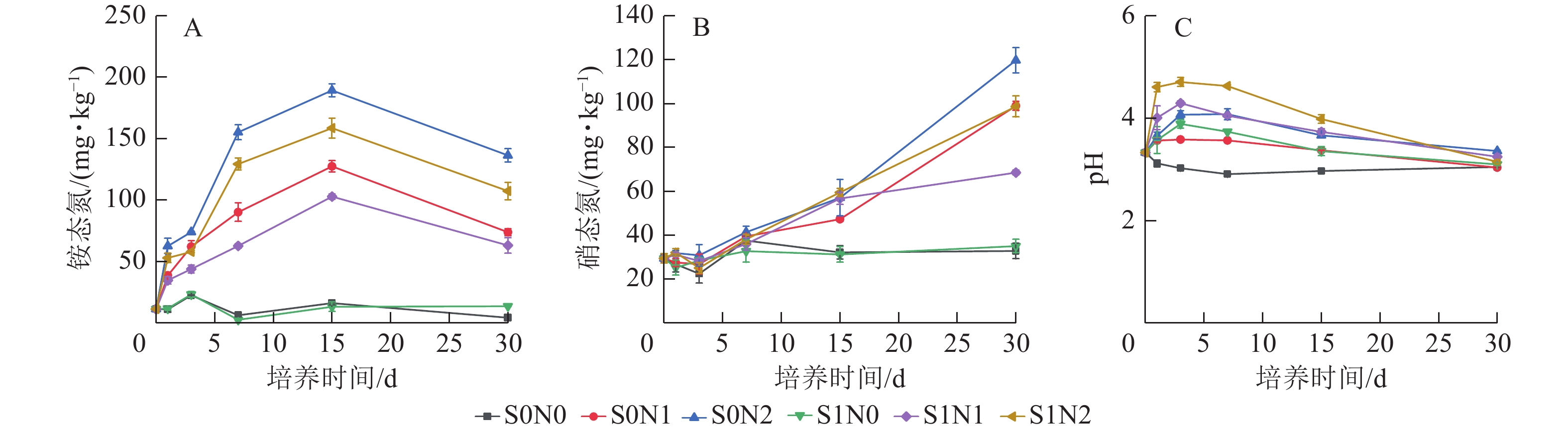
土壤铵态氮质量分数在0~15 d呈快速上升趋势,在15~30 d缓慢下降(图1A)。高氮处理(N2)的铵态氮质量分数显著高于中氮(N1)和无氮(N0)处理。在相同氮水平下,S1N2处理组在第15 天的铵态氮质量分数为 158.58 mg·kg−1,较S0N2处理下降16.2 %。统计检验显示:硅、氮及培养时间对铵态氮质量分数均产生极显著主效应(P <0.001),且硅与氮的交互作用、硅与培养时间的交互作用、氮与培养时间的交互作用以及硅、氮、时间三者的交互作用均达到极显著水平(P < 0.001)。
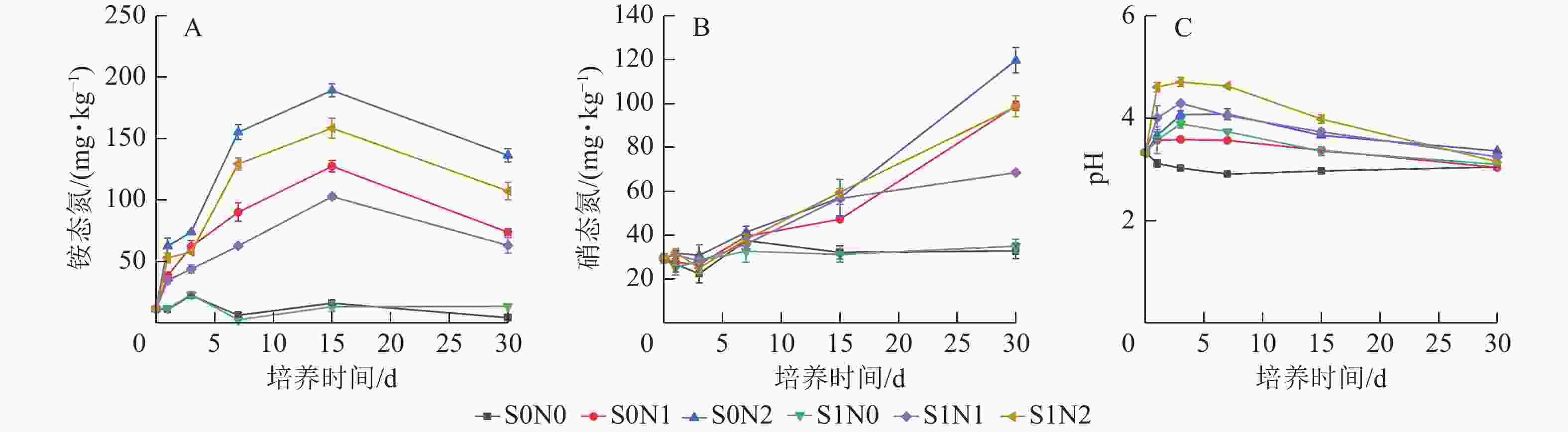
Figure 1. Temporal dynamics of soil ammonium nitrogen content (A), soil nitrate nitrogen content (B) and soil pH (C) in different treatments
硝态氮对氮添加的响应与铵态氮一致(高氮处理下明显高于中氮、无氮),但各处理硝态氮质量分数均呈现随培养时间延长而持续上升的总体特征(图1B)。在相同氮水平下,施硅组(S1N1、S1N2)的硝态氮质量分数明显低于不施硅组(S0N1、S0N2)。
所有处理组(除S0N0外)的土壤pH在培养0~3 d内均出现短暂上升,随后又下降的整体趋势(图1C)。随着氮水平的提高,土壤pH变化幅度增大,初始pH升高更明显,随后下降趋势也更为剧烈。而硅的添加虽能在1~3 d暂时提升pH值,使含硅处理组(S0N1、S0N2)的pH明显高于无硅对照组(S0N0),但其后续下降趋势与不施硅组相似。
-
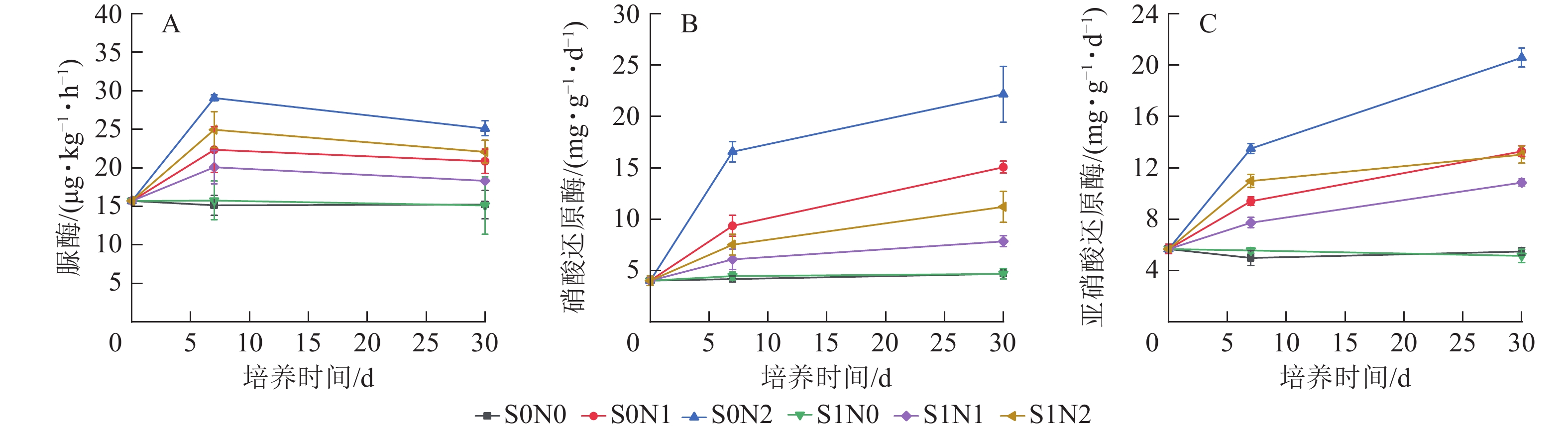
脲酶活性在0~7 d快速上升,7~30 d缓慢下降,且高氮处理组脲酶活性高于中氮和无氮处理(图2A),S0N2处理第7天脲酶(29.06 μg·kg−1·h−1)较S0N0 (15.14 μg·kg−1·h−1)高92.1%。双因素方差分析显示:硅和氮对脲酶活性均具有显著主效应(P<0.05),但硅与氮之间的交互作用不显著。
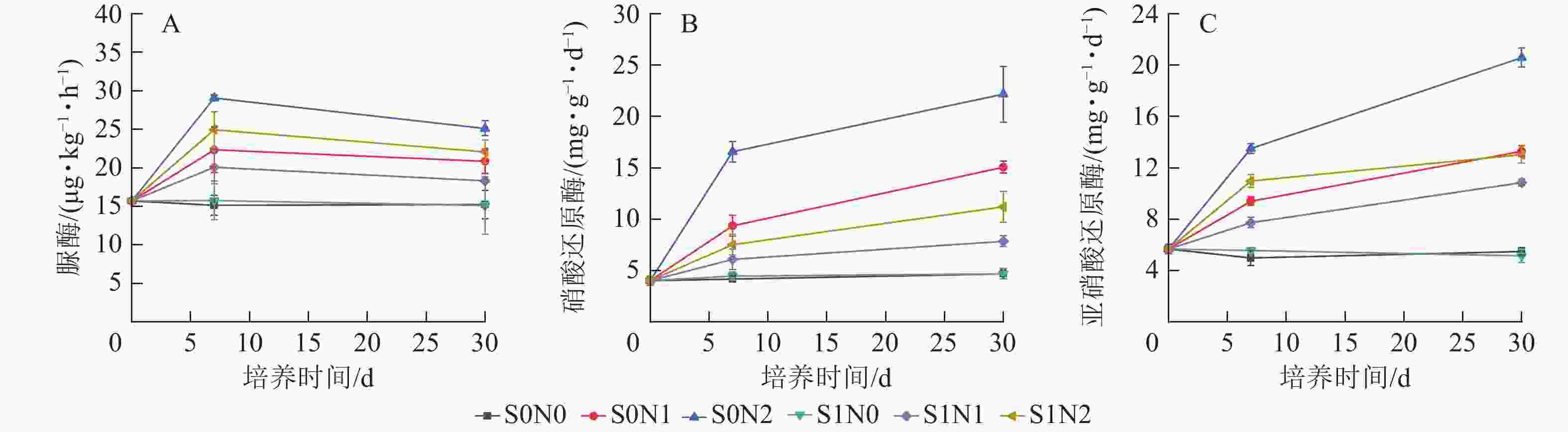
Figure 2. Temporal dynamics of urease activity (A), nitrate reductase activity (B) and nitrite reductase activity (C) in different treatments
不同处理下硝酸还原酶和亚硝酸还原酶的时间变化趋势在各处理组中表现出高度一致性(图2B和图2C)。高氮处理下硝酸还原酶活性明显高于中氮、无氮处理,且施硅在氮添加处理下具有抑制效应。双因素方差分析显示:硅、氮及两者交互作用对硝酸还原酶和亚硝酸还原酶活性均存在显著影响(P<0.05)。
-
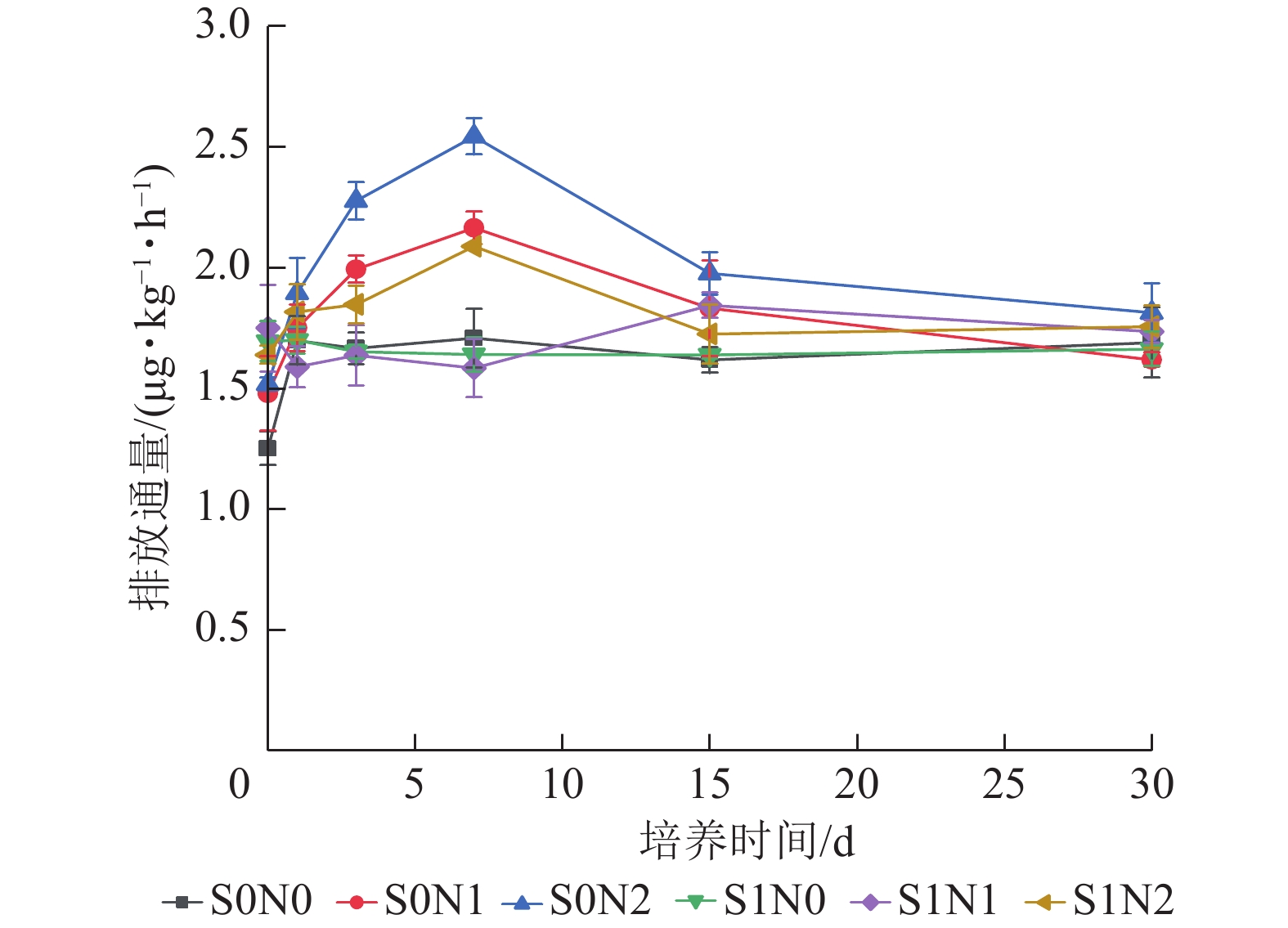
氧化亚氮排放通量整体呈现初期上升、中期出现峰值、后期逐步衰减的动态特征(图3),培养初期(0~7 d)各处理氧化亚氮排放通量明显上升,在第7天出现排放峰值,之后逐步减少,最终趋于稳定。高氮水平下,施硅处理(S1N2)氧化亚氮排放通量峰值较S0N2处理降低5.1%;中氮水平下,S1N1处理峰值较S0N1降低14.3%。双因素方差和重复测量分析表明:硅、氮及培养时间对氧化亚氮排放通量均存在极显著主效应(P<0.001),且硅与氮、硅与时间、氮与时间的交互作用均极显著(P<0.001),三者的交互作用显著(P<0.01)。
Figure 3. Temporal dynamics of nitrous oxide emission flux in different treatments
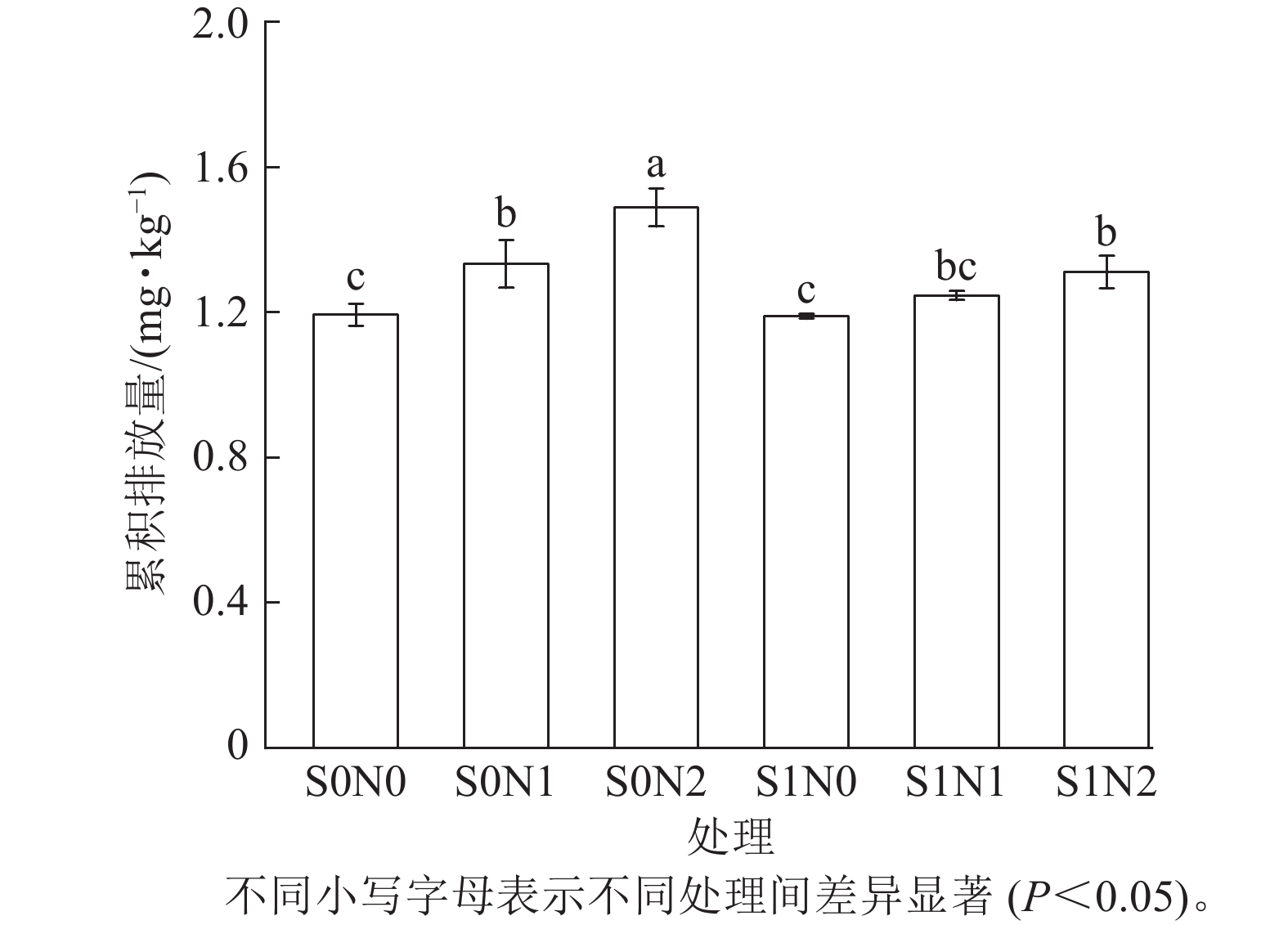
不同处理的土壤氧化亚氮累积排放量有明显差异(图4),S0N2处理的土壤氧化亚氮累积排放量最高,达1.55 mg·kg−1,S1N0处理的累积排放量最低,为1.16 μg·kg−1。
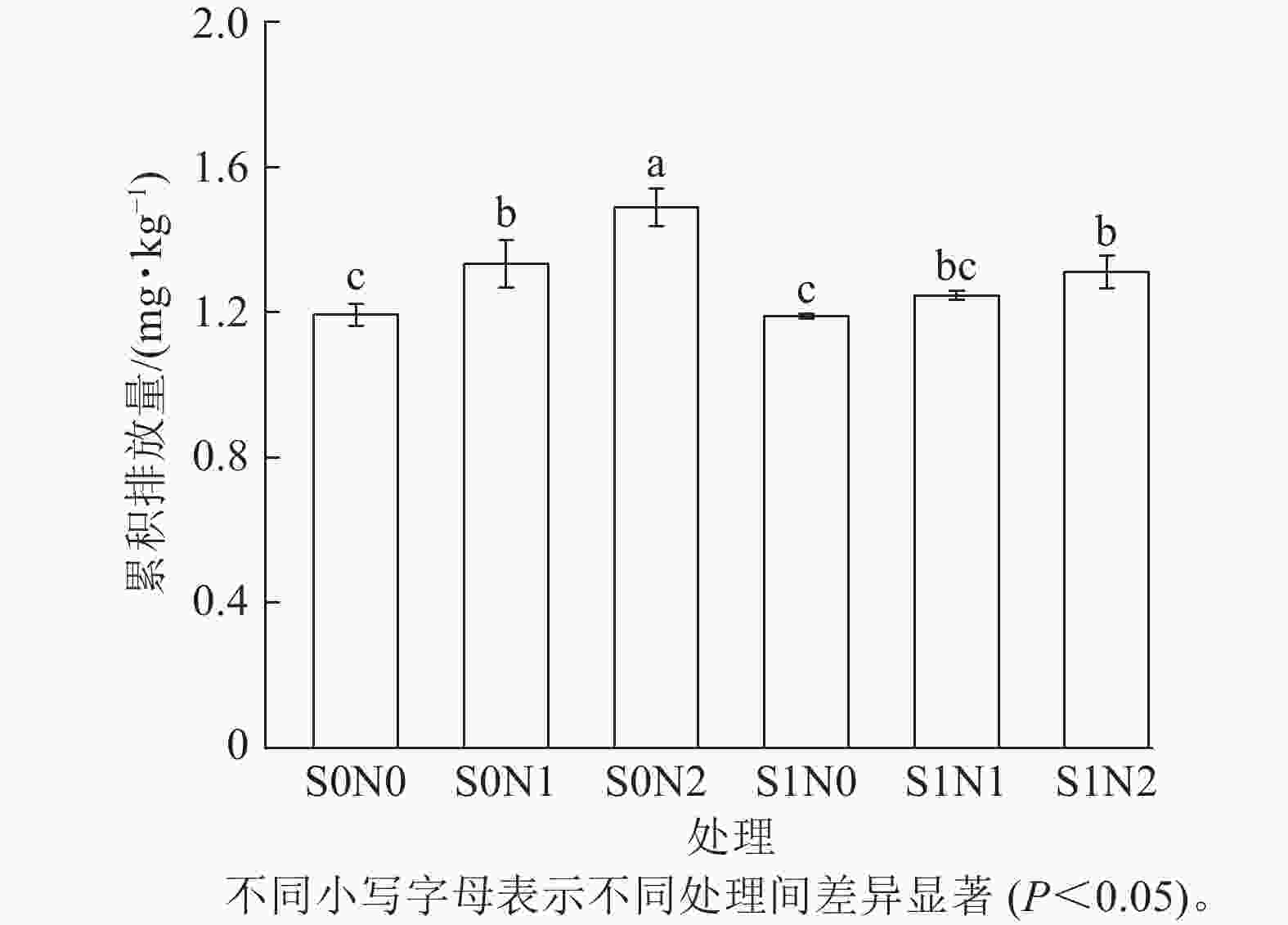
Figure 4. Cumulative nitrous oxide emissions in different treatments
在相同硅水平下,氮添加显著提升了氧化亚氮累积排放(P<0.05),且高氮处理效应强于中氮。无硅条件下,S0N2处理的氧化亚氮累积排放量(1.49 mg·kg−1)较S0N0处理(1.19 mg·kg−1)增加25.2%,较S0N1处理(1.33 mg·kg−1)增加12.0%。
在相同氮水平下,硅对氮添加引起的氧化亚氮累积排放量具有显著抑制效应 (P<0.05),且该效应随氮水平升高而增强。相同氮水平下,S1N2处理的氧化亚氮累积排放量(1.31 mg·kg−1)较S0N2处理降低12.1%,S1N1处理的氧化亚氮累积排放量(1.25 mg·kg−1)较S0N1处理降低6.0%。
双因素方差分析显示:硅与氮的添加均对土壤氧化亚氮累积排放量产生了极显著的主效应(P<0.001),且硅与氮的交互作用对土壤氧化亚氮累积排放量也达显著水平(P<0.05)。
-
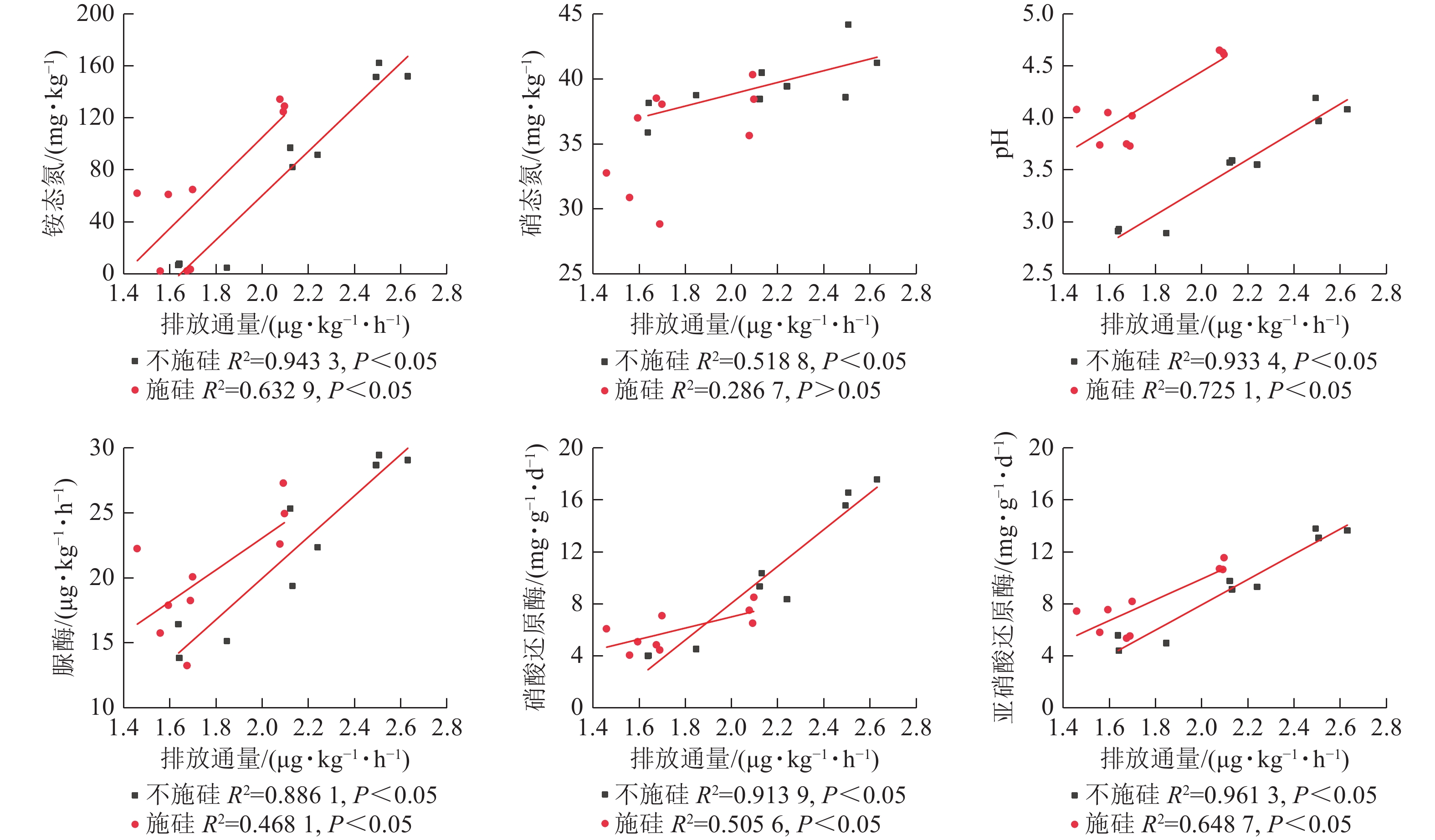
在不施硅处理中,氧化亚氮排放通量和累积排放量与铵态氮、硝态氮、pH、脲酶、硝酸还原酶及亚硝酸还原酶均呈显著正相关(表1,P<0.05),其中铵态氮和亚硝酸还原酶的相关性最强。施硅处理后,氧化亚氮排放通量与上述指标的关联性普遍减弱,硝态氮与累积排放量甚至无显著相关性。线性回归分析显示:施硅组的回归线性斜率均低于不施硅组(图5),表明硅添加降低了土壤理化性质对氧化亚氮排放的驱动强度。
处理 相关系数 铵态氮 硝态氮 pH 脲酶 硝酸还原酶 亚硝酸还原酶 不施硅处理氧化亚氮排放通量 0.971** 0.720* 0.966** 0.941** 0.956** 0.980** 不施硅处理氧化亚氮累积排放量 0.978** 0.749* 0.971** 0.944** 0.968** 0.974** 施硅处理氧化亚氮排放通量 0.796* 0.535 0.851** 0.684* 0.711* 0.805** 施硅处理氧化亚氮累积排放量 0.778* 0.457 0.862** 0.761* 0.613 0.821** 说明:*表示相关显著(P<0.05);**表示相关极显著(P<0.01)。 Table 1. Correlation between nitrous oxide emission flux and soil physicochemical properties under different fertilization treatments
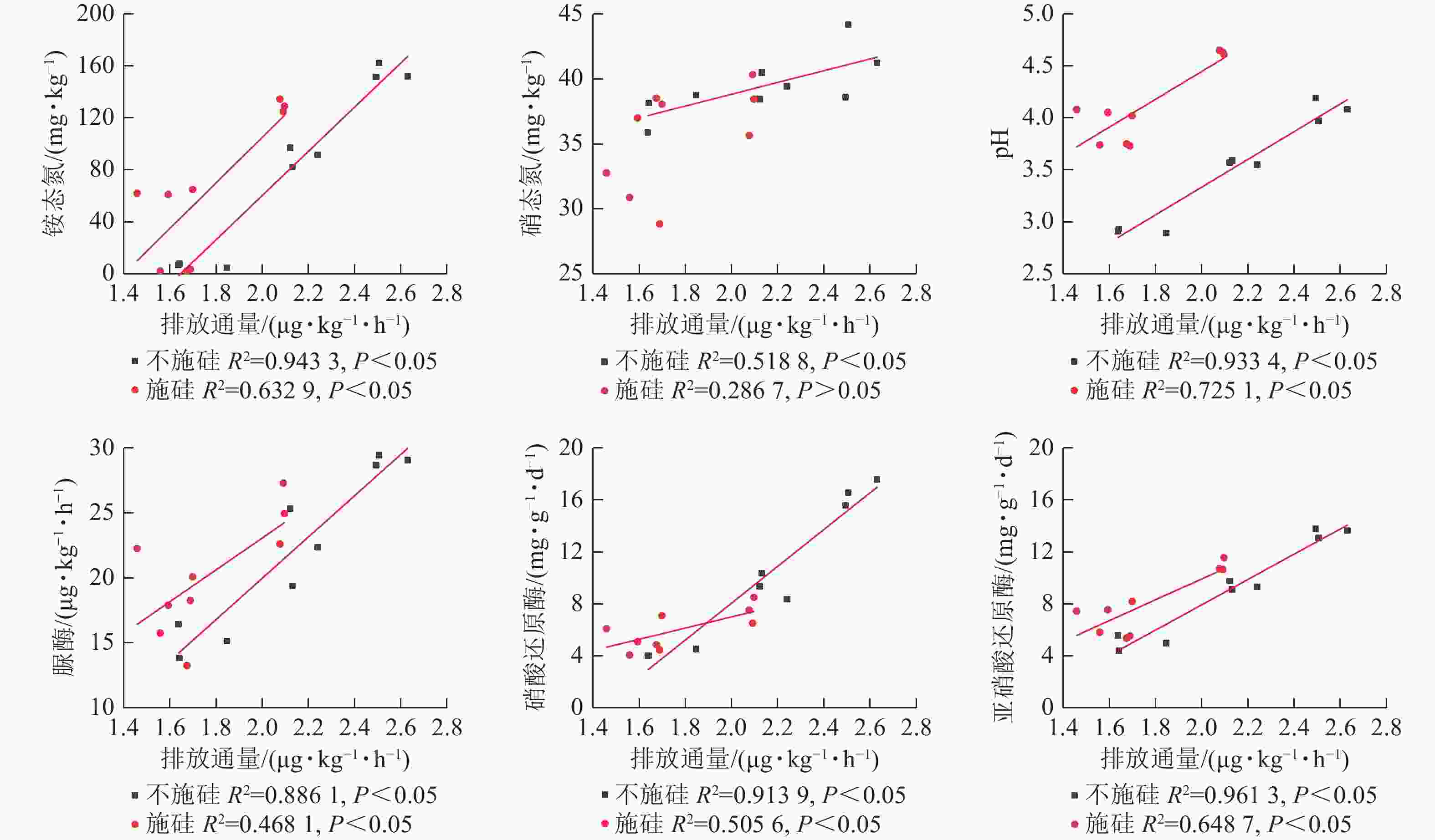
Figure 5. Linear regression relationships of nitrous oxide emission flux with nitrogen, pH, and enzyme activities across different treatment groups
-
硅添加明显抑制了毛竹林土壤氧化亚氮的排放。施硅处理的氧化亚氮最大排放通量和累积排放量均显著低于不施硅处理。这与曾冬萍[23]使用钢渣硅肥、SONG等[11]使用偏硅酸钠和矿渣基硅酸盐肥料对水稻田温室气体排放影响的研究结果一致。但本研究硅肥的减排效应明显低于上述水稻田研究。可能的原因包括:①曾冬萍[23]、SONG等[11]使用的硅肥中含有Fe2O3等成分,这些成分可能放大了硅肥的减排效果[35];②硅肥施用提高了土壤水分和氧气含量,增强了植物对氮素底物的吸收,降低了土壤和田面水中底物的浓度,从而减少了土壤氧化亚氮的产生[36−37]。排除硅肥其他成分和植物对土壤氧化亚氮的贡献后,硅肥对土壤本身也会产生影响,特别是对土壤铵态氮、pH、硝酸还原酶和亚硝酸还原酶活性的调控,从而改变土壤氧化亚氮的生成过程。此外,硅肥的施用使氧化亚氮排放高峰延迟至第15天出现,进一步延缓了氧化亚氮的排放过程。
-
本研究表明:硅氮耦合对氧化亚氮累积排放量具有拮抗效应,且这种效应随氮添加水平的增加而增强。在无氮、中氮和高氮水平下,硅添加分别减少氧化亚氮累积排放量0.1%、6.6%和11.9%。可能的机制包括:①硅添加降低了土壤氮浓度[38−39],从而缓解了高氮浓度对氧化亚氮还原酶的抑制作用;②硅添加提高了土壤pH[40],改变了氮添加导致的酸化环境。土壤pH提高可以增加氧化亚氮还原酶的活性,从而促进氧化亚氮还原为氮气[41]。此外,土壤pH可以调控氨氧化细菌和氨氧化古菌的群落结构[42],进而影响硝化细菌的硝化作用和氧化亚氮的产生。
-
本研究表明:在未施用硅肥的土壤中,氧化亚氮的产生主要受到硝酸还原酶、铵态氮和脲酶的调控。而在施用硅肥后,氧化亚氮的产生主要受到亚硝酸还原酶、pH和铵态氮的调控。此外,硅添加在第7天显著提高了土壤中脲酶、硝酸还原酶和亚硝酸还原酶的活性,从而降低了尿素的水解(减少了土壤中的铵氮底物)和土壤的反硝化过程(改变了反硝化过程的限速步骤),进而影响了氧化亚氮的排放通量。本研究表明:在未施用硅肥的土壤中,氧化亚氮的产生受到脲酶水解和硝氮向亚硝氮转化速率的影响,而在施用硅肥后,氧化亚氮的产生受到亚硝氮向一氧化氮转化速率的影响。硅肥的施用降低了硝氮向亚硝氮以及亚硝氮向一氧化氮的转化速率,从酶活性的角度支持了SONG等[11]在水稻田中发现的硅添加能降低nirS/nirK基因丰度,从而改变氧化亚氮排放的结果。
-
本研究表明:硅肥添加通过调节土壤pH和关键酶活性,有效降低了毛竹林土壤的氧化亚氮排放。这一发现不仅具有重要的环境价值,也展现出可观的经济效益潜力。在环境方面,硅肥不仅能有效降低氧化亚氮排放,还可能通过改变氮素转化过程提高氮肥利用率,减少氮素淋溶所带来的水体污染风险。在经济方面,硅肥能直接促进竹笋的产量与品质提升,实现增产增收。覃其云等[43]研究显示:麻竹Dendrocalamus latiflorus笋用林在施用专用硅肥后,竹笋年均增产达5 912 kg·hm−2,可新增收入约11 825元·hm−2。因此,在毛竹林中推广施用硅肥,兼具环境与经济效益的双重优势,具有广阔的推广前景。
-
施用硅肥能够显著抑制土壤中氧化亚氮的排放通量。同时,硅肥的添加减少了土壤中的铵态氮、硝态氮质量分数以及脲酶、硝酸还原酶和亚硝酸还原酶的活性,改变了氮素的转化过程。此外,硅肥的添加使得氧化亚氮的减排效果随着氮水平的升高而增强,同时,硅肥的添加推迟了土壤中氧化亚氮排放高峰的出现时间。
Effects of silicon-nitrogen interaction on nitrous oxide emissions from soil in a Moso bamboo forest under laboratory incubation
doi: 10.11833/j.issn.2095-0756.20250511
- Received Date: 2025-09-24
- Accepted Date: 2026-05-08
- Rev Recd Date: 2026-04-24
- Available Online: 2026-07-21
- Publish Date: 2026-08-20
-
Key words:
- silicon-nitrogen coupling /
- nitrous oxide emissions /
- Moso bamboo forest /
- soil enzyme activity /
- silicon fertilizer
Abstract:
| Citation: | HUANG Junhao, CAO Xuexian, YANG Jianwei, et al. Effects of silicon-nitrogen interaction on nitrous oxide emissions from soil in a Moso bamboo forest under laboratory incubation[J]. Journal of Zhejiang A&F University, 2026, 43(4): 714−722 doi: 10.11833/j.issn.2095-0756.20250511

|
 DownLoad:
DownLoad: