






-
土壤是生态系统的重要组成部分。近几十年来,中国土壤重金属污染问题日益突出。2014年环境保护部和国土资源部共同发布的《全国土壤污染状况调查公报》显示:镉和铅污染在中国土壤重金属污染问题中尤为突出,其点位超标率分别高达7.0%和1.5%[1]。由于镉和铅具有高毒性、高持久性、不可生物降解性和隐蔽性等特点[2],一般通过自然因素(如土壤成土母质中含有的镉和铅矿物的存积)和人为因素(如不合理不科学的污水灌溉,施用化肥、农药和农膜等)导致其在土壤中积累[3],破坏了土壤生态平衡,严重影响了耕地质量和农产品的安全生产,造成不可估量的经济损失,并且对人类的生命健康造成严重危害。因此,对重金属污染土壤进行治理与修复已刻不容缓。
生物质炭作为一种环境友好型材料,是将生物质废弃物置于缺氧或低氧环境下,经高温热解形成的产物,具有表面官能团丰富、孔隙结构发达和比表面积大等[3]特点。生物质炭对土壤中的重金属具有较强的固定作用,主要包括络合作用、沉淀作用、物理吸附及离子交换作用等[4],因此,生物质炭的施用可以降低土壤中重金属的生物有效性以及迁移性[5]。目前,已有许多学者在生物质炭对土壤中镉和铅生物有效性和毒性的影响方面开展了研究。李冬琴等[6]研究表明:皇竹草Pennisetum sinese炭的添加分别使土壤中有效态镉和铅的含量降低40.7%和45.4%。周雷等[7]研究发现:施用稻草壳炭显著降低了土壤有效态镉和铅的含量(降低幅度为57.1%和74.6%)。不同源废弃物制备的生物质炭对重金属污染土壤的原位修复效果存在一定的差异。随着中国城市化的快速发展,城市生态文明建设取得了显著进展,旅游、餐饮等服务行业也得到了长足发展。然而,这也造成了城镇园林废弃物和厨余废弃物的大量产生[8-9],为城市生态环境建设带来了新的挑战。若将富含有机质的园林废弃物和厨余废弃物制备成生物质炭,并将其用于城市周边重金属(如镉和铅)污染农田的治理,不但可以实现废弃物的资源化利用,也将为利用生物质炭修复重金属污染土壤提供新思路和新途径。
国家统计局2014年的环境统计数据显示:中国木材加工等园林废弃物高达255.1万t,其中细叶榕Ficus microcarpa是中国南方典型的城镇园林植物[10]。另外,中国年产厨余垃圾量约6000万t,其中,北京、上海、重庆、广州等超大城市餐厨垃圾的日产量均超过1000 t,且日产量的年增速在10%以上[8]。中国小龙虾Procambarus clarkii餐饮产业发展迅速,现已成为世界上最大的小龙虾消费国。然而,小龙虾壳大多被作为厨余废弃物随意处置,中国每年约10万t的小龙虾壳被丢弃[11]。基于此,本研究从以废治污的角度出发,利用厨余废弃物小龙虾壳和园林废弃物细叶榕修剪枝为供试原材料制成生物质炭,进而通过盆栽试验探究添加不同质量比(1%、3%)的2种生物质炭对土壤有效养分、土壤镉和铅生物有效性以及土壤酶活性的影响,以期为以生物质炭作为钝化剂固定土壤中的重金属和改善作物品质提供理论依据。
-
供试土壤:采集自浙江省绍兴市上虞区(30º00′N,120º79′E)一处水稻Oryza sativa田的表层土壤(0~20 cm)。该稻田毗邻一处废弃的铅锌矿山,因此该区域受到不同重金属复合污染。供试土壤的基本性质:pH 5.8,有机碳为13.3 g·kg−1,碱解氮、有效磷和速效钾分别为118.0、114.0和1.8 mg·kg−1。总镉和总铅分别为0.5和736.2 mg·kg−1,均已超出了GB 15618—2018《土壤环境质量 农用地土壤污染风险管控标准(试行)》的风险筛选值(总镉为0.4 mg·kg−1,总铅为100.0 mg·kg−1)。将所采土壤剔除碎石及植物根系等杂物,风干后过2 mm不锈钢筛备用。
供试生物质炭制备:细叶榕Ficus microcarpa修剪枝收集自广东省佛山市某公园,将枝条切碎通风晾干。小龙虾壳收集自浙江农林大学附近餐馆,用自来水洗净后于室温下晾干,在105 ℃下烘至恒量。利用小型炭化设备(ECO-8-10,湖州宜可欧环保科技有限公司)在650 ℃限氧条件下热解2 h制得细叶榕炭和小龙虾壳炭,将2种生物质炭研磨并过2 mm筛,混合均匀后取出部分样品用于基本理化性质表征,剩下样品储存备用。
供试植物:小红萝卜Raphanus sativus种子购买于浙江省杭州市种子种苗管理站。
-
盆栽试验于浙江农林大学温室进行。称取3 kg过筛土装入塑料盆(直径为22.6 cm,高为13.5 cm),参照相关研究[12-13]分别加入质量比为1%和3%的小龙虾壳炭(CSB)和细叶榕炭(FMB)。以不添加生物质炭的土壤为对照(ck),共5个处理,每个处理重复3次。选用尿素、磷酸二氢钾为基肥,每盆施用0.25 g·kg−1氮,0.32 g·kg−1五氧化二磷和0.2 g·kg−1氧化钾后充分混合均匀。不同处理盆栽按照随机区组方式摆放(盆栽位置定期更换)。初次浇水至田间持水量的70%,平衡3 d后,于2020年9月撒播小红萝卜种子,每盆均匀播种10颗,发芽7 d后间苗,每盆中留下长势相同的健康植株5株。盆栽期间日常管理为定期浇水、松土以及防治病虫,同时隔3 d记录1次植物长势。生长50 d后分别采集土壤样品和植物样品,将采集的一部分土壤样品风干磨细后过2 mm筛待测;取小红萝卜可食用部分,先后用自来水和去离子水洗净,置于105 ℃烘箱内杀青30 min后于65 ℃条件下烘干至恒量,记录其生物量。
-
土壤理化性质的测定参照《土壤农业化学分析方法》[14]。其中,土壤pH和电导率分别采用1.0∶2.5 (质量体积比)和1.0∶5.0 (质量体积比)土水比,用FE20型pH计[梅特勒-托利多仪器(上海)有限公司]和DDS-307型电导率仪(上海虹益仪器仪表有限公司)测定。土壤有机碳质量分数采用重铬酸钾外加热法测定。有效磷质量分数采用Olsen法,用0.5 mol·L−1 碳酸氢钠浸提后,采用分光光度计(UVA132122,Thermo electron corporation,英国)测定。土壤速效钾质量分数经乙酸铵浸提后用火焰光度计(FP640, 上海仪电分析有限公司)法测定。土壤碱解氮质量分数采用碱解扩散法测定。植物镉和铅质量分数采用硫酸-过氧化氢法消解后用ICP-OES测定。
采用荧光微孔板检测技术测定土壤β-1,4-葡萄糖苷酶(β-1,4-glucosidase, BG)、亮氨酸氨基肽酶(leucine amino peptidase, LAP)、β-N-乙酰基氨基葡萄糖苷酶(β-1-4-N-acetyl-glucosaminidase, NAG)和β-木糖苷酶的酶(β-1,4-xylosidase, XYL)活性。原理是利用底物与酶水解进行荧光检测,以荧光强度反映酶的活性[15]。具体操作步骤如下:将2.0 g土壤鲜样称入100 mL离心管,加入40 mL醋酸缓冲液(50 mol·L−1, pH 5.0),振荡30 min (25 ℃,180 r·min−1)。将振荡好的溶液用60 mL醋酸缓冲液(50 mol·L−1, pH 5.0)洗入烧杯中,制成土壤均质悬浊液。将混有酶和底物混合物的微孔板放入25 ℃培养箱里,在黑暗环境中培养3 h,培养后用多功能酶标仪(SynergyTM H1, Biotek,美国)在荧光激发波长365 nm和检测光波长450 nm下测定反应液荧光值。
供试生物质炭的pH采用炭水比1∶20 (质量体积比)法制备悬浊液,用FE20型pH计[梅特勒-托利多仪器(上海)有限公司]测定;灰分使用ASTM D1726-84法测定;碳(C)、氢(H)和氮(N)质量分数采用元素分析仪(Flash EA1112, Thermo Finnigan,意大利)测定;比表面积(BET)在77 K氮气条件下用比表面分析仪(TristarII3020,Micromeritica Instrument Corporation,美国)测定;利用X射线能量色谱仪(EDS) (K-Alpha+; Thermo Fisher Scientific,美国)测定生物质炭的矿物元素组成;表面形貌特征采用扫描电镜(SEM)分析仪(SU-8010,日立公司,日本)测定;表面官能团采用傅里叶红外光谱仪(FTIR) (NICOLET iS10,Thermo Fisher Scientific,美国)测定。供试生物质炭重金属全量采用硝酸-氢氟酸-高氯酸消解法,用ICP-MS测定。
-
应用Excel 2010和SPSS 19.0进行数据处理和分析,采用单因素方差分析和Duncan多重比较分析不同处理间土壤理化性质、有效养分和重金属有效态质量分数的差异显著性(P<0.05),相关性分析采用皮尔逊(Pearson’s)相关分析法进行检验确定显著性,并用Origin 2021作图。
-
由表1可知:与细叶榕炭相比,小龙虾壳炭具有更高的pH和灰分,这主要取决于原材料属性和热解温度[16]。研究表明:小龙虾壳主要由碳酸钙(约40%)、甲壳素(30%~35%)、蛋白质(25%~30%)以及微量脂类物质组成[17]。碳酸钙等矿物组分在热解过程中可以被保留和浓缩,使得所制得的生物质炭灰分较高[18],生物质炭的碱性主要与这些碳酸盐类矿物组分有关。有研究表明:随热解温度升高,生物质炭中的碳酸盐总量增加,从而使其pH有所提高[16]。此外,小龙虾壳在高温热解过程中酸性官能团(如—COOH)的破坏分解也会导致生物质炭pH升高。与小龙虾壳炭相比,细叶榕炭具有更大的比表面积和孔径(表1)。这是由于细叶榕所含纤维素、半纤维素和木质素等物质在高温热解过程中出现分解和结构塌陷所致[19],从而形成了更多的微孔结构(图1A)。
表 1 供试生物质炭的基本理化性质
Table 1. Selected physicochemical properties of the biochars studied
生物质炭 pH 灰分/
(g·kg−1)碳/
(g·kg−1)氮/
(g·kg−1)氢/
(g·kg−1)铁/
(mg·kg−1)锰/
(mg·kg−1)铜/
(mg·kg−1)锌/
(mg·kg−1)镉/
(mg·kg−1)铅/
(mg kg−1)比表面积/
(m2·g−1)孔径/
nm小龙虾壳炭 10.6 477.9 254.7 13.3 8.2 1 891.0 319.0 48.0 91.0 0.3 2.9 12.1 16.5 细叶榕炭 9.6 84.6 762.7 5.2 22.5 779.0 16.0 11.0 52.0 0.1 3.8 21.6 153.5 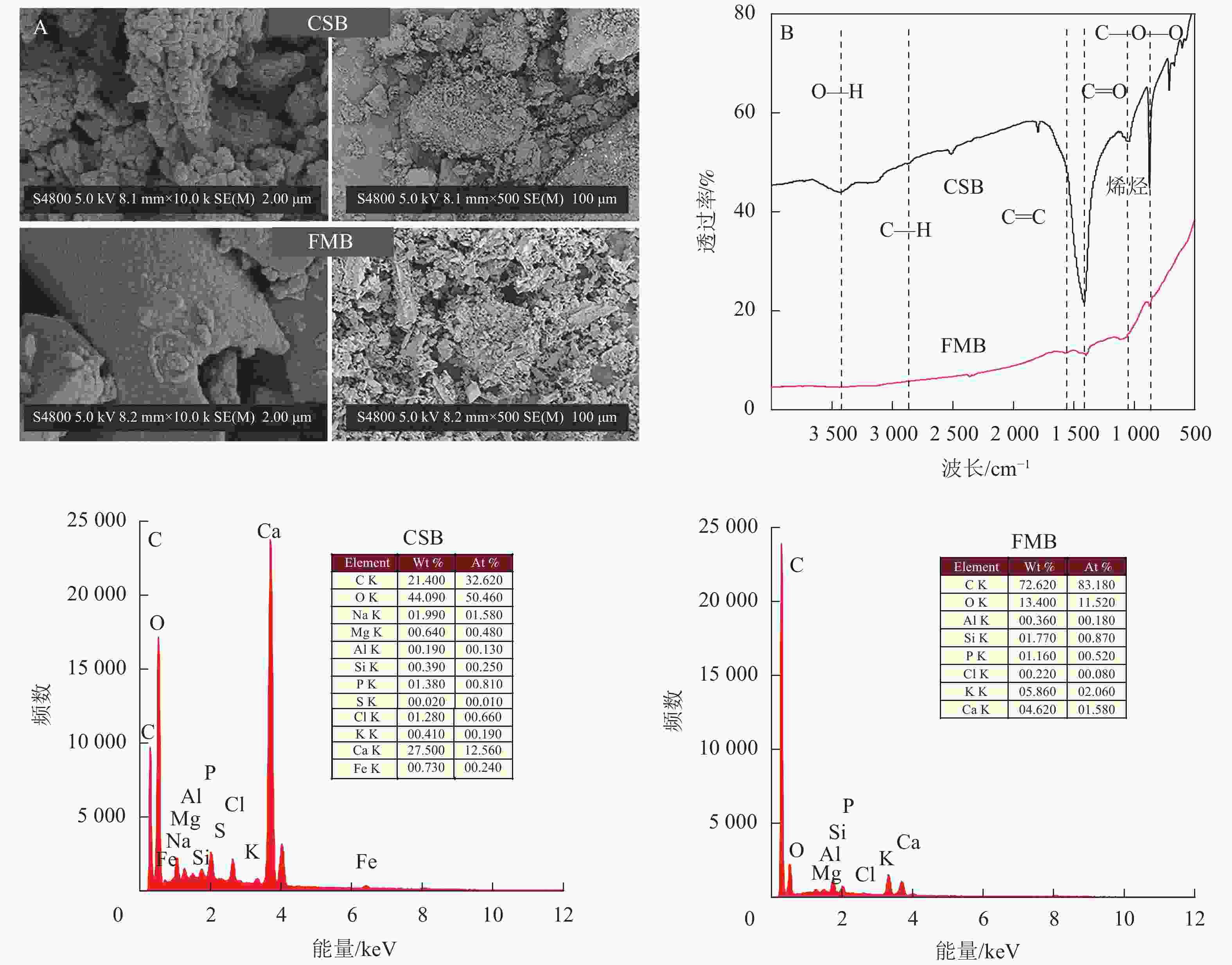
图 1 小龙虾壳炭(CSB)和细叶榕炭(FMB)的扫描电镜(SEM)图谱(A)、傅里叶变换红外(FTIR)图谱(B)和X射线能谱分析(EDS)图谱(C)
Figure 1. Scanning electron microscope (SEM) images (A), Fourier transform infrared (FTIR) spectra(B) and energy dispersive X-ray spectra (EDS) spectra (C) of crawfish shell biochar (CSB) and F. microcarpa biochar (FMB)
SEM图谱(图1A)显示:与小龙虾壳炭相比,细叶榕炭表面呈现出较为光滑的不规则块状骨架结构,这可能是因为经过炭化后,细叶榕本身的导管结构被部分保存[20]。小龙虾壳炭表面呈现出较为粗糙的团簇结构,且表面附着大量颗粒状物质。由于较高温度下生物质炭表面形成的孔隙结构出现破裂和塌陷,这些团簇颗粒可能是因生物质在热解过程中物理破碎而形成[21]。通过FTIR图谱可知(图1B):2种生物质炭上均检出—OH(3420 cm−1)、芳香性官能团C=O(1380 cm−1)、醚类官能团C—O—C(1050~1100 cm−1)和烯烃(650~1000 cm−1)等基团的特征峰。此外,与小龙虾壳炭相比,细叶榕炭还含有芳香族C=C(1596 cm−1)。这可能是在高温热解过程中细叶榕炭的芳香化程度逐渐增强所致[9]。小龙虾壳在热解过程中,随着挥发分的析出,碱性矿物元素(如钾、钠、钙等)被保留和富集。EDS图谱(图1C)也显示:与细叶榕炭相比,小龙虾壳炭中钙、钠较高,而细叶榕炭的碳质量分数明显高于小龙虾壳炭。
-
与ck相比,不同生物质炭处理下土壤pH均有显著提高(P<0.05)(图2A),且随着配施比例的增加而增加(pH从高到低依次为3%CSB、1%CSB、3%FMB、1%FMB、ck),这是由于CSB(pH 10.6)和FMB (pH 9.6)本身呈碱性,将其施入土壤后可直接提高土壤pH。此外,3%CSB处理下土壤pH较ck增加最为显著(增幅为25.8%),这可能与CSB在热解过程中形成较多的碳酸盐(MgCO3和CaCO3)有关。这些碳酸盐进入土壤后通过水解作用产生OH−从而提高pH[16]。
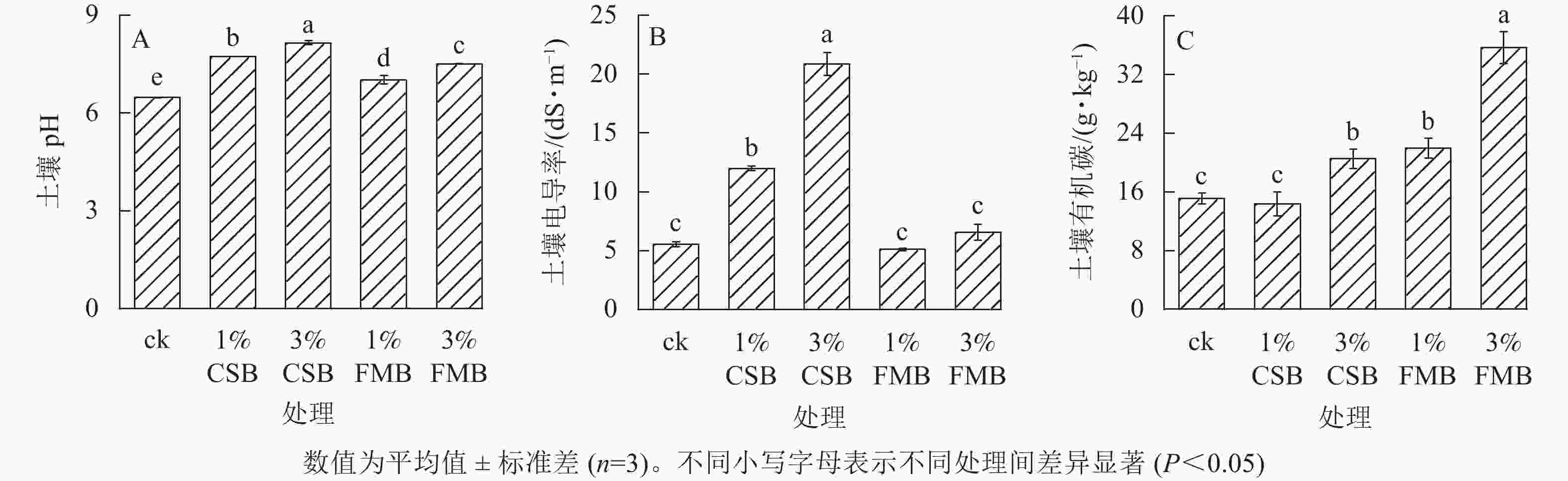
图 2 施用小龙虾壳炭(CSB)和细叶榕炭(FMB)对土壤pH(A)、电导率(B)和有机碳(C)质量分数的影响
Figure 2. Effect of crawfish shell biochar (CSB) and F. microcarpa biochar (FMB) applications on soil pH(A), electrical conductivity(B) and organic carboncontent(C)
土壤电导率通常情况下可以反映土壤的盐分状况,是影响土壤养分循环和土壤微生物活性的主要因素之一。本研究表明:施用不同比例的FMB后,土壤电导率较ck并无显著差异,而施用CSB则显著提高了土壤电导率(增幅为116.9%~277.1%),且随施用量提高效果更显著(图2B)。这是因为CSB属于动物源生物质炭,其本身灰分较高(表1)。另外,在热解小龙虾壳的过程中,随着挥发分的析出,碱性矿物元素(如钾、钠、钙等)被保留和富集(图1C),将CSB施入土壤后引入了较多的盐基离子,从而提高土壤电导率[22]。有研究表明:较高的土壤电导率可能会对作物的生长造成负面影响[23]。对于施用CSB对土壤盐分和电导率的潜在影响还需后续进一步探索,比如如何通过调整CSB的施用方式、施用比例,或者将其改性等措施以避免因其施用向土壤中引入过多的盐分。
与ck相比,除1%CSB处理外,其余生物质炭处理下土壤的有机碳质量分数均显著提高(P<0.05) (图2C),且FMB处理下土壤有机碳质分数显著高于CSB处理。这是因为FMB是植物源类生物质炭,其碳质量分数较高且芳香化结构稳定[24],因此将其施入土壤后可直接提高土壤有机碳质量分数。其次,FMB相较于CSB有更大的比表面积和更丰富的孔径结构(表1),因而具有更强的吸附能力,可将土壤中的小分子有机物吸附至其孔径内,从而在一定程度上有利于提高土壤的固碳能力。此外,3%的FMB处理下土壤有机碳提升效果最显著,较ck升高了76.5%。曾爱等[25]研究发现:土壤有机碳含量的提高幅度与生物质炭的施用量呈显著正相关。
-
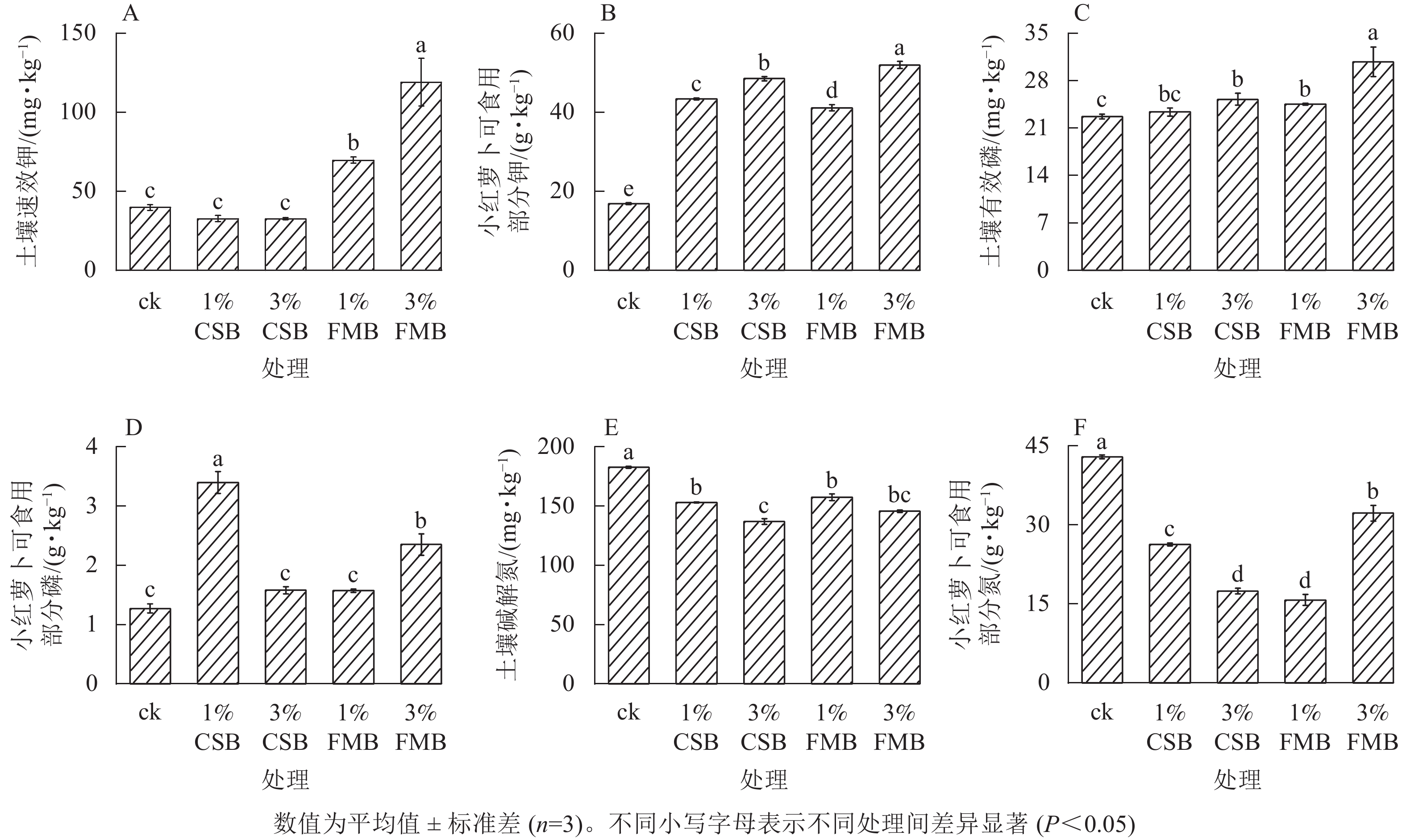
与ck相比,施用不同比例的FMB后,土壤速效钾质量分数显著(P<0.05)提高,且3%FMB处理效果最显著,较ck提高了174%,而施入不同比例的CSB后,土壤速效钾质量分数无显著变化(图3A)。FMB中含有植物生长所必需的钾元素(图1C),施入土壤后,其释放的可溶性钾可直接增加土壤速效钾质量分数[26]。与ck相比,不同施炭处理均提高了小红萝卜可食用部分的钾质量分数(图3B),提升效果从大到小依次为3%FMB(208.1%)、3%CSB(187.7%)、1%CSB(157.4%)、1%FMB(143.9%)。这是因为生物质炭施入土壤后,一方面可以直接释放有效钾,另一方面可以通过吸附作用间接减少土壤中钾的淋溶损失[27],从而提高土壤中钾素的有效性,促进小红萝卜可食用部分对土壤中钾的吸收。
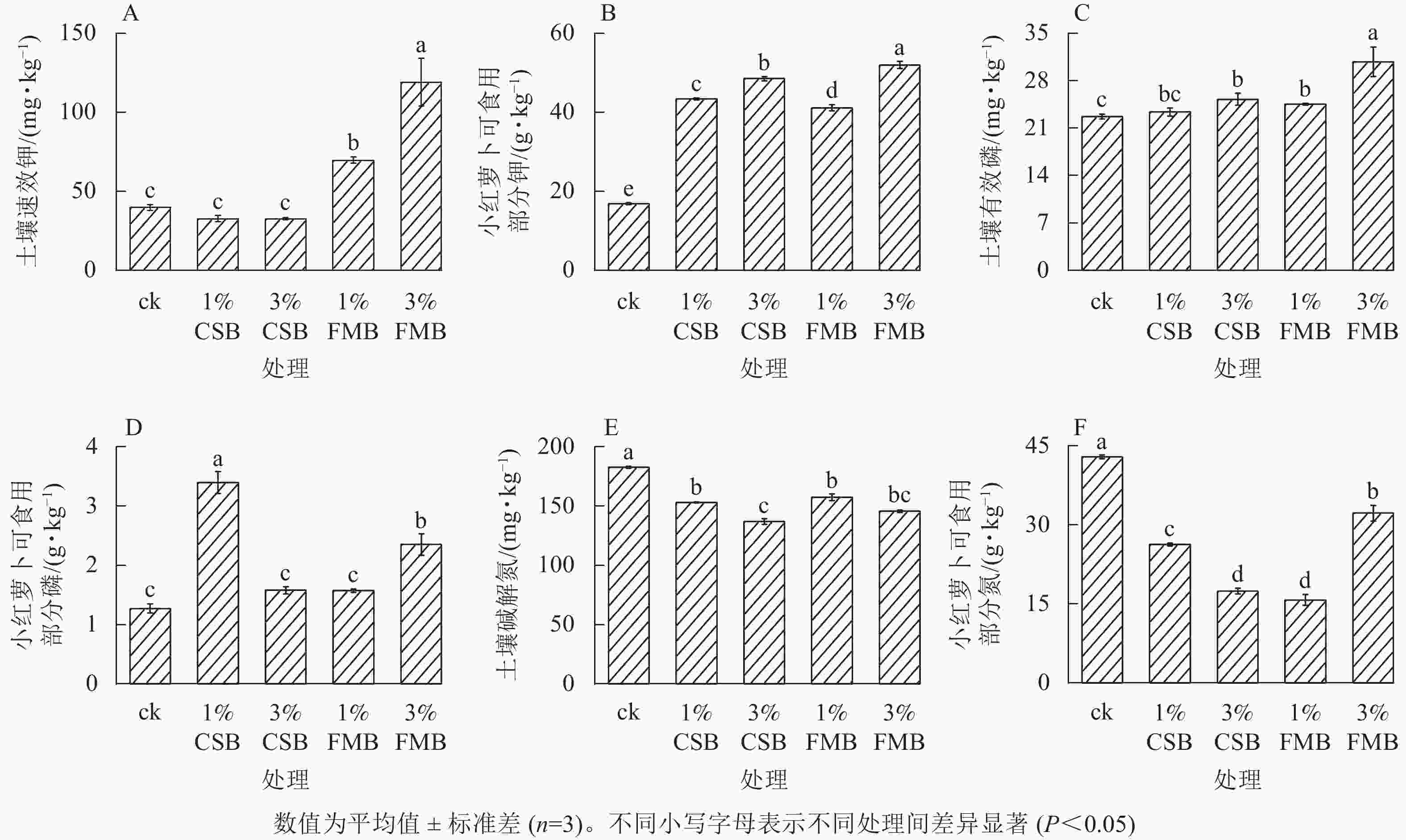
图 3 施用小龙虾壳炭(CSB)和细叶榕炭(FMB)对土壤有效养分和小红萝卜可食用部分的氮、磷和钾质量分数的影响
Figure 3. Effect of crawfish shell biochar (CSB) and F. microcarpa biochar (FMB) applications on N, P and K availability and their uptake in the edible part of radish
施用不同生物质炭后,土壤中有效磷质量分数提高了9.7%~37.3%(图3C)。其中3%生物质炭处理提高土壤有效磷的效果较1%处理更显著。与ck相比,3%FMB处理下土壤有效磷质量分数提高幅度达37.3%。这一方面是由于FMB含有较高的磷(图1A),另一方面与3%FMB处理下土壤pH显著提高有关。DELUCA等[28]发现:生物质炭可以通过提高土壤pH和阳离子交换量促进磷素有效性的提高,且生物质炭的施用可通过减少铁和铝的交换量增加磷的活性。同时,与ck相比,3%FMB处理显著(P<0.05)提高了小红萝卜可食用部分的磷质量分数(图3D),这与土壤中有效磷升高有关。此外,生物质炭的施用可提高微生物活性,进而将难以被植物利用的有机磷矿化为无机磷,从而可被植物吸收利用[29]。
与ck相比,施用不同生物质炭后土壤碱解氮质量分数均显著(P<0.05)降低(图3E),且随两者比例的增加而降低。可能是由于CSB和FMB用量的增加提高了土壤pH,进而可以通过调控土壤碳氮比影响氮素的有效性[30]。ASAI等[31]研究发现:随着生物质炭用量的增加,土壤中碳氮比升高,氮有效性降低。此外,与对照相比,不同施炭处理均显著(P<0.05)降低了小红萝卜可食用部分的氮质量分数(图3F),这可能是由于不同施炭处理均降低了土壤碱解氮质量分数,从而间接影响了小红萝卜可食用部分氮的积累。
-
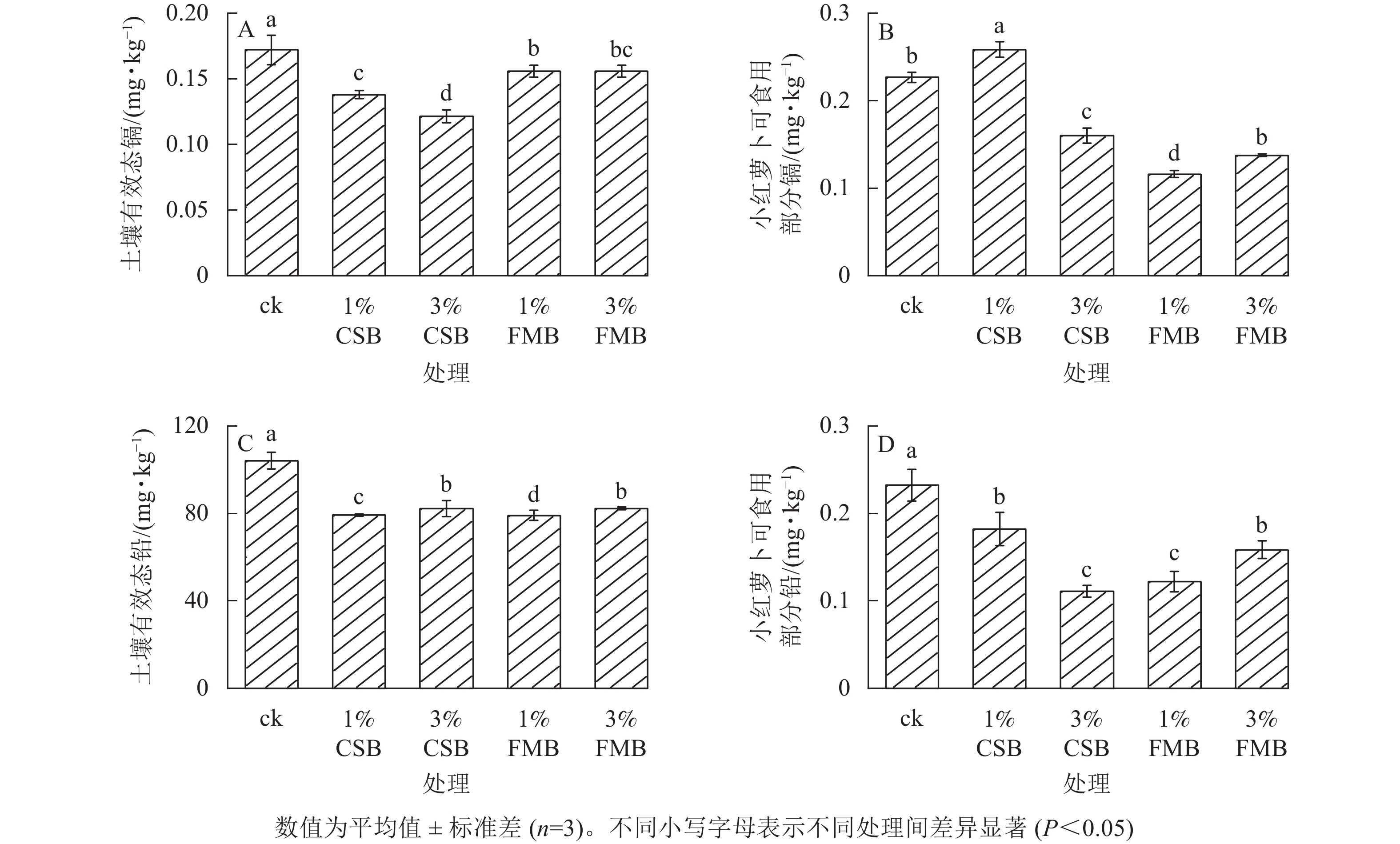
与ck相比,施用CSB和FMB均显著(P<0.05)降低了土壤有效态镉,降幅从大到小依次为3%CSB(91.1%)、1%CSB(73.3%)、3%FMB(64.3%)、1%FMB(60.7%)(图4A)。首先,施用CSB和FMB后使得土壤pH升高,增加了土壤胶体表面的负电荷,从而提高土壤对Cd2+的吸附能力[7]。其次,相较于FMB处理,CSB处理对土壤pH的增幅更大(图2A),促进CdCO3、Cd3(PO4)2和Cd(OH)2等难溶物质的形成[32],从而降低土壤有效态镉质量分数;并且相较于FMB处理,CSB显著提高了土壤电导率(图2B),引入了较多的盐基离子,可提高土壤的盐基离子饱和度,促进重金属的离子交换,进而提高生物质炭对镉的吸附[22]。此外,土壤有效态镉和pH、电导率均呈极显著(P<0.01)负相关(图5)也证实了这一观点。与此同时,生物质炭具有较大的比表面积、丰富的孔隙结构,通过静电吸附将镉吸附至生物质炭表面,降低镉的移动性[32]。另外,生物质炭表面的官能团(图1B)(如氨基、烃基和羟基等)也可与镉发生络合反应从而降低土壤有效态镉含量[33]。与ck相比,施用3%CSB、1%FMB和3%FMB可使小红萝卜可食部分中的镉质量分数显著(P<0.05)降低(图4B)。这是由于生物质炭的施用显著降低了土壤中镉的有效性,并且由于生物质炭表面具有多孔性结构和较强的表面吸附能力,通过吸附土壤中的重金属而影响镉在土壤-植物体系的迁移能力[34],从而降低小红萝卜可食用部分镉的富集。此外,1%CSB处理下小红萝卜可食部分中的镉质量分数显著(P<0.05)提高,这可能由于施用一定量的生物质炭改变了土壤环境,在植物根际作用下原本被钝化的镉可能再次转化为植物可吸收利用态[35]。另外,郭荣荣等[36]研究发现:红油麦菜Lactuca sativa可食部位重金属的变化还与其根系生理、根际微生物代谢活动及重金属由根系向地上部的转移有关。然而,1%CSB对植物吸收镉的促进作用的相关机制还需结合其他手段(如微生物、镉结合形态等)进行进一步探讨。

图 4 施用小龙虾壳炭(CSB)和细叶榕炭(FMB)对土壤有效态镉和铅质量分数和小红萝卜可食用部分的镉和铅质量分数的影响
Figure 4. Effect of crawfish shell biochar (CSB) and F. microcarpa biochar (FMB) applications on the availability of Cd and Pb and their accumulation in the edible part of radish

图 5 土壤理化性质、酶活性和有效态镉和铅的相关性分析
Figure 5. Correlation analyses between soil physicochemical properties, enzyme activities and concentrations of available Cd and Pb
与ck相比,CSB和FMB处理均显著(P<0.05)降低了土壤有效态铅质量分数。降低幅度由大到小依次为1%FMB(26.1%)、1%CSB(23.9%)、3%CSB(21.2%)、3%FMB(21.0%)(图4C)。首先,与ck相比较,生物质炭的施加均提高了土壤pH,使得Pb2+与土壤中的OH−在碱性条件下形成沉淀[37]。其次,铅可以与土壤中OH−和
${\rm{PO}}_{4}^{{\rm{3-}}}$ 相结合,形成Pb9(PO4)6及Pb3(CO3)OH2等不溶性氢氧化物和磷酸盐[38],从而降低了土壤有效态铅质量分数。总体而言,FMB的施用对降低土壤铅有效性的效果优于CSB。这是因为在土壤中施用FMB后,土壤的有效磷质量分数增加,铅可与${\rm{PO}}_{4}^{{\rm{3-}}}$ 离子形成难溶性的磷酸盐[39]。另外,由于FMB具有较大的比表面积和丰富的孔隙结构(表1和图1A),为表面或界面相关反应提供丰富的活性位点[12],更有利于生物质炭对土壤中铅的物理吸附,从而降低土壤溶液中铅的有效性[5]。在本研究中,不同生物质炭处理下,土壤中有效态铅质量分数和小红萝卜可食用部分铅质量分数(图4D)也具有较大差异,其原因可能是由于铅质量分数在小红萝卜不同器官内的占比不同所致。与此相似,陈新红等[40]研究发现:由于铅在稻谷各器官中的处理比例不同,稻谷中铅质量分数与土壤中铅质量分数也存在较大差异。 -
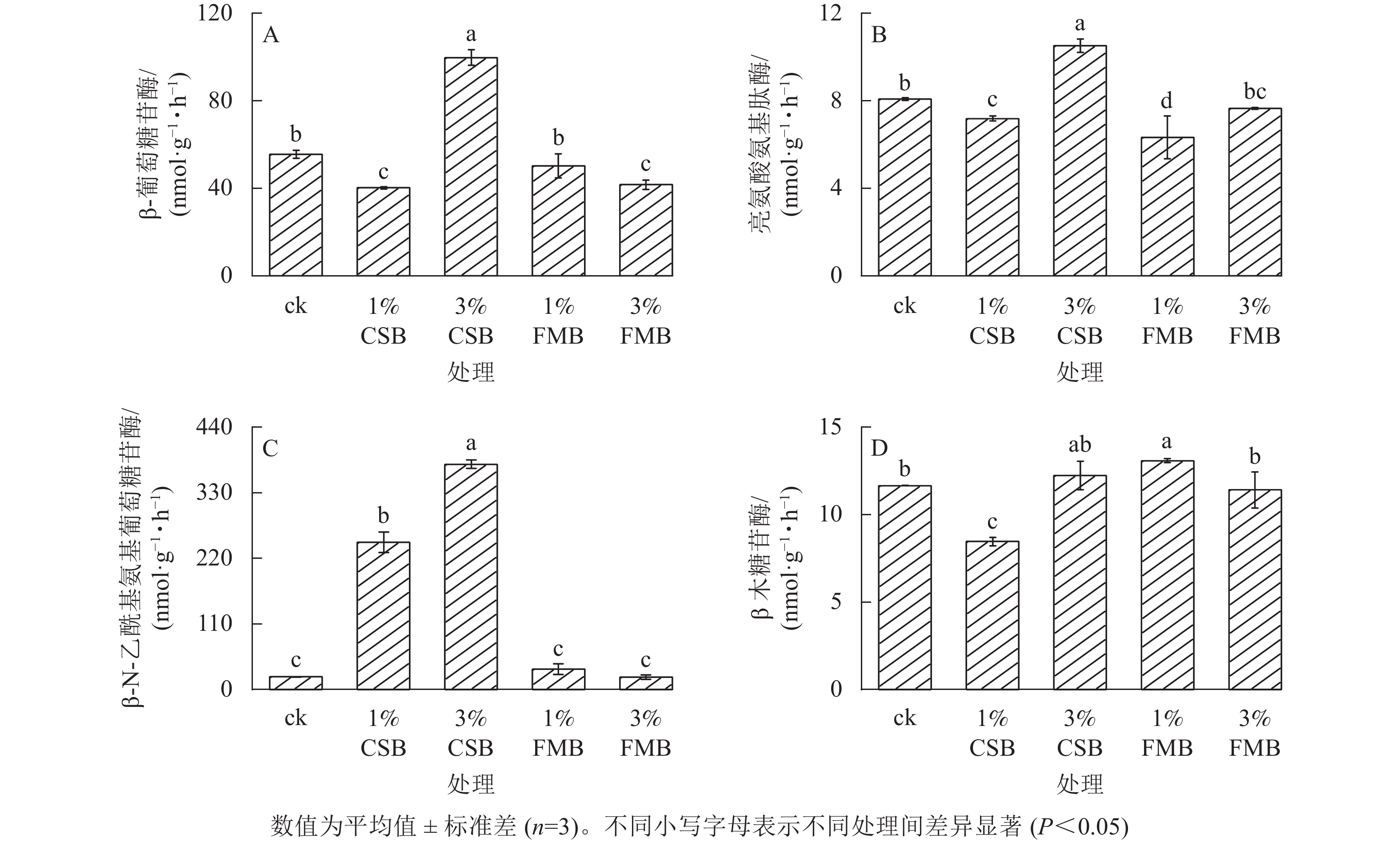
与ck相比,3%CSB处理均显著(P<0.05)提高了土壤中β-葡萄糖苷酶、亮氨酸氨基肽酶和β-N-乙酰基氨基葡萄糖苷酶活性(图6A~C),较ck分别提高了79.7%、30.3%和1668.6%。1%CSB处理可提高β-N-乙酰基氨基葡萄糖苷酶活性(图6C),但不同施用量的FMB对β-N-乙酰基氨基葡萄糖苷酶活性均无显著影响。另外,与ck相比,1%FMB处理提高了β木糖苷酶活性(图6D),而对β-葡萄糖苷酶和β-N-乙酰基氨基葡萄糖苷酶活性无显著影响。β-葡萄糖苷酶、亮氨酸氨基肽酶、β-N-乙酰基氨基葡萄糖苷酶和β木糖苷酶均属于水解酶,能够催化土壤中糖类和蛋白质的分解,在土壤碳氮循环过程中起着重要影响[15]。相关性分析表明(图5):土壤中有效态镉与土壤中β-葡萄糖苷酶和亮氨酸氨基肽酶活性呈显著(P<0.05)负相关,与土壤中的β-N-乙酰基氨基葡萄糖苷酶呈极显著负(P<0.01)相关;而土壤中有效态铅质量分数和土壤酶活性无显著相关性。施用3%CSB后土壤酶活性提高的原因主要归因于土壤中有效态镉的降低(图4A),从而降低了重金属对土壤微生物的胁迫,并且由于CSB含有较高的灰分及碳、氮等营养元素(表1),能够促进微生物的活性及其对底物的利用速率,当施加3%CSB后在一定程度上促进了土壤养分循环,微生物生长环境因子得以改善,进而提高土壤酶活性[41]。而1%FMB和3%FMB处理对土壤酶活性没有显著影响,原因可能是FMB本身难降解(表1、图1),易分解组分质量分数低,惰性炭质量分数高并不能作为促进土壤微生物活性的可利用底物,从而无法诱导土壤酶活性提高[15]。与此同时,FMB相较于CSB具有更大的比表面积和孔径表面积(表1),吸附能力强,迫使更多的酶与生物质炭结合或导致酶活性位点改变,进而阻碍了酶与底物的结合,从而影响酶活性[42]。

图 6 施用小龙虾壳炭(CSB)和细叶榕炭(FMB)对土壤酶活性的影响
Figure 6. Effect of crawfish shell biochar (CSB) and F. microcarpa biochar (FMB) applications on soil enzyme activities
-
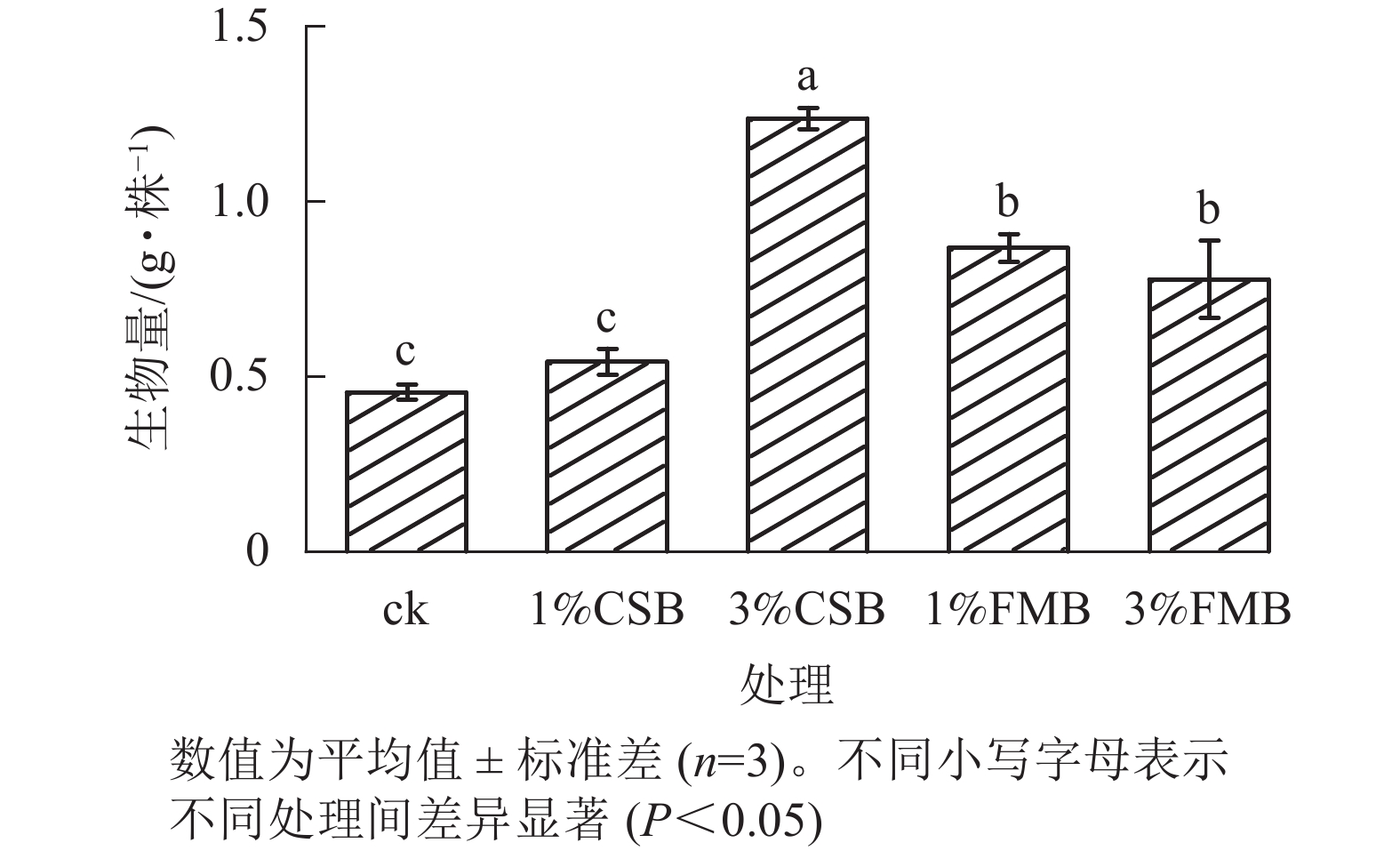
由图7可知:与ck相比,除1%CSB外,其余生物质炭的施用均显著(P<0.05)提高了小红萝卜可食用部分的生物量,且3%CSB提升效果最佳,较ck提高了171.48%。这是由于将CSB和FMB施入土壤后钝化了污染土壤中的镉和铅,降低了其对土壤和植物的胁迫作用,从而提高了小红萝卜可食用部分生物量。此外,由于生物质炭(尤其CSB)含有较高的中微量元素(如铁、锰、锌等)(表1),将其施入土壤后可以直接提高土壤养分水平并促进作物对这些养分的吸收,进而促进作物生长。张丽娜[43]研究发现:中微量元素(如钙、镁、锌、硼、铁等)可以通过参与小红萝卜肉质根可溶性蛋白质、可溶性糖、维生素C等多种物质的合成促进小红萝卜生长和产量提升。另一方面,施用生物质炭后,土壤中磷、钾等养分的有效性显著提高,从而促进小红萝卜生长,增加其可食用部分的生物量[44]。另外,CSB的施用显著提高了土壤中各种生物酶的活性(图6A~C),促进了土壤中的养分循环,从而可以有效促进小红萝卜的生长和生物量的提高。同样,LEBRUN等[45]发现:以白桦Betula platyphylla 为原材料制得的1%和3%的生物质炭均可提高亚麻Linum usitatissimum生物量,且3%生物质炭处理对促进亚麻生长具有较好效果。刘晓霞等[46]研究亦表明:施用高配比的骨炭(300和400 kg·hm−2)促进小红萝卜产量的效果优于低配比(150 kg·hm−2)。
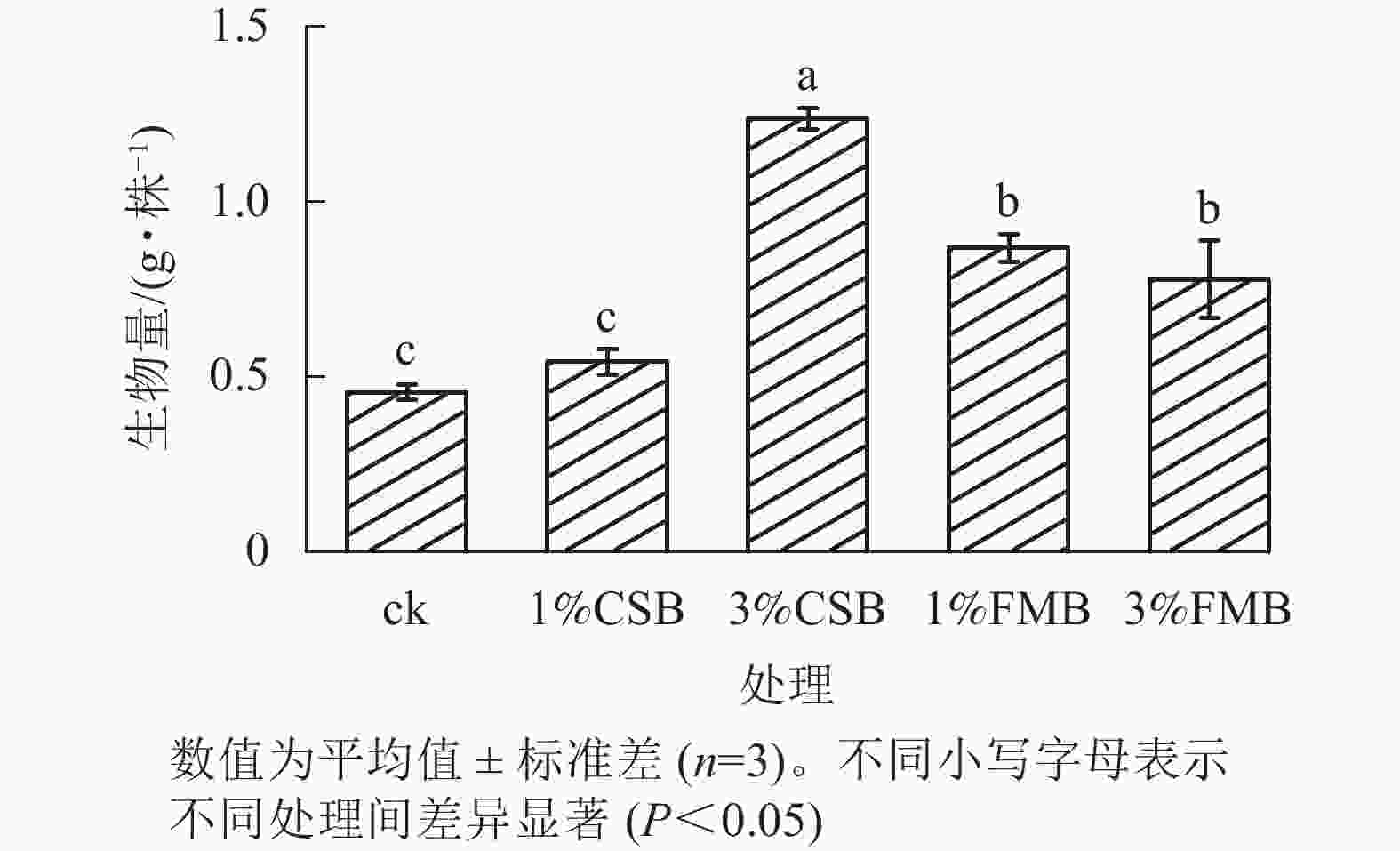
图 7 施用小龙虾壳炭(CSB)和细叶榕炭(FMB)对小红萝卜可食用部分生物量的影响
Figure 7. Effect of crawfish shell biochar (CSB) and F. microcarpa biochar (FMB) applications on the biomass of the edible part of radish
-
施用的3%小龙虾壳炭可以显著改善土壤肥力,提高土壤中养分的有效性,进而显著提高小红萝卜可食用部分生物量和土壤酶活性,在改善土壤肥力,促进作物品质和产量提升方面具有较大应用潜力。
施用小龙虾壳炭和细叶榕炭均显著降低了土壤有效态镉和铅的质量分数,且小龙虾壳炭效果更明显,因此,小龙虾壳炭是一种可作为修复镉和铅复合污染土壤的良好土壤改良剂。
综上所述,将农林废弃物细叶榕和厨余废弃物小龙虾壳炭化还田均可有效改善土壤肥力并缓解土壤重金属污染状况。同时,与细叶榕炭相比,小龙虾壳炭的应用前景更佳,为利用生物质炭修复重金属复合污染农田土壤提供了新思路。
Effects of biochar from Procambarus clarkii shells and Ficus microcarpa branches on soil nutrients and bioavailability of Cd and Pb
-
摘要:
目的 探讨施用小龙虾Procambarus clarkii壳炭(CSB)和细叶榕Ficus microcarpa炭(FMB)对复合污染土壤理化性质及作物生长的影响。 方法 在 650 ℃限氧条件下热解制备厨余废弃物小龙虾壳炭和园林废弃物细叶榕炭。以不同质量比(0、1%、3%)施入小红萝卜Raphanus sativus盆栽土壤,测定和分析施用小龙虾壳炭和细叶榕炭对土壤中镉和铅有效性、养分转化、土壤酶活性及小红萝卜生长的影响。 结果 3%FMB处理对土壤有机碳、有效磷和速效钾质量分数提升效果最显著(P<0.05),较对照的增幅分别为135.8%、35.4%和173.7%。除1%CSB处理外,其余生物质炭处理下土壤中有效态镉和铅质量分数较对照均显著降低(P<0.05),降幅分别为60.7%~91.1%和21.0%~26.1%。3%CSB处理对土壤β-葡萄糖苷酶、亮氨酸氨基肽酶和β-N-乙酰基氨基葡萄糖苷酶活性提升效果最显著(P<0.05),较对照分别提高79.7%、30.3%和1 668.5%。不同比例CSB和FMB的施用均显著(P<0.05)提高了小红萝卜可食部分的生物量,且3%CSB处理的提升效果最显著(P<0.05),较对照提高了171.5%。 结论 与细叶榕炭相比,小龙虾壳炭在提高土壤酶活性,降低土壤中镉和铅生物有效性以及提升作物品质和产量方面效果更为优越,更适合作为镉-铅复合污染土壤修复的潜在应用材料。图7表1参46 Abstract:Objective This study aims to investigate the effects of biochar derived from Procambarus clarkii shells (CSB) and Ficus microcarpa (FMB) on physicochemical properties of contaminated soil and plant growth. Method The biochar was prepared by pyrolysis of both typical kitchen and garden wastes at 650 ℃ with limited oxygen supply. A pot trial was conducted using Raphanus sativus with different mass ratios (0, 1%, 3%) to investigate the effects of different doses of CSB and FMB on cadmium (Cd) and lead (Pb) bioavailability in soil, nutrient transformation, soil enzyme activity, and R. sativus growth. Result 3% FMB treatment had the most significant effect on improving contents of soil organic carbon, available phosphorus and available potassium (P<0.05), which increased by 135.8%, 35.4%, and 173.7% respectively compared with the control. Except 1% CSB treatment, other biochar treatments significantly (P<0.05) decreased the concentrations of available Cd by 60.7%−91.1% and Pb by 21.0%−26.1%. 3% CSB treatment showed significant increase in soil enzyme activities such as β-glucosidase, leucine aminopeptidase and β-N-acetylglucosamine (P<0.05), which were 79.7%, 30.3% and 1668.5% higher than that of the control, respectively. All biochar treatments significantly (P< 0.05) increased the biomass of the edible part of R. sativus. 3% CSB treatment had the most significant improvement effect (P< 0.05), which increased by 171.5% compared with the control. Conclusion The CSB is more effective than the FMB in improving soil enzyme activity, reducing Cd and Pb bioavailability in soil, and improving crop quality and yield. It is more suitable as a potential material for the remediation of soils co-contaminated with Cd and Pb. [Ch, 7 fig. 1 tab. 46 ref.] -
Key words:
- biochar /
- heavy metal /
- soil contamination /
- in-situ remediation /
- waste disposal
-
图 1 小龙虾壳炭(CSB)和细叶榕炭(FMB)的扫描电镜(SEM)图谱(A)、傅里叶变换红外(FTIR)图谱(B)和X射线能谱分析(EDS)图谱(C)
Figure 1 Scanning electron microscope (SEM) images (A), Fourier transform infrared (FTIR) spectra(B) and energy dispersive X-ray spectra (EDS) spectra (C) of crawfish shell biochar (CSB) and F. microcarpa biochar (FMB)
图 2 施用小龙虾壳炭(CSB)和细叶榕炭(FMB)对土壤pH(A)、电导率(B)和有机碳(C)质量分数的影响
Figure 2 Effect of crawfish shell biochar (CSB) and F. microcarpa biochar (FMB) applications on soil pH(A), electrical conductivity(B) and organic carboncontent(C)
图 3 施用小龙虾壳炭(CSB)和细叶榕炭(FMB)对土壤有效养分和小红萝卜可食用部分的氮、磷和钾质量分数的影响
Figure 3 Effect of crawfish shell biochar (CSB) and F. microcarpa biochar (FMB) applications on N, P and K availability and their uptake in the edible part of radish
图 4 施用小龙虾壳炭(CSB)和细叶榕炭(FMB)对土壤有效态镉和铅质量分数和小红萝卜可食用部分的镉和铅质量分数的影响
Figure 4 Effect of crawfish shell biochar (CSB) and F. microcarpa biochar (FMB) applications on the availability of Cd and Pb and their accumulation in the edible part of radish
图 5 土壤理化性质、酶活性和有效态镉和铅的相关性分析
Figure 5 Correlation analyses between soil physicochemical properties, enzyme activities and concentrations of available Cd and Pb
图 6 施用小龙虾壳炭(CSB)和细叶榕炭(FMB)对土壤酶活性的影响
Figure 6 Effect of crawfish shell biochar (CSB) and F. microcarpa biochar (FMB) applications on soil enzyme activities
图 7 施用小龙虾壳炭(CSB)和细叶榕炭(FMB)对小红萝卜可食用部分生物量的影响
Figure 7 Effect of crawfish shell biochar (CSB) and F. microcarpa biochar (FMB) applications on the biomass of the edible part of radish
表 1 供试生物质炭的基本理化性质
Table 1. Selected physicochemical properties of the biochars studied
生物质炭 pH 灰分/
(g·kg−1)碳/
(g·kg−1)氮/
(g·kg−1)氢/
(g·kg−1)铁/
(mg·kg−1)锰/
(mg·kg−1)铜/
(mg·kg−1)锌/
(mg·kg−1)镉/
(mg·kg−1)铅/
(mg kg−1)比表面积/
(m2·g−1)孔径/
nm小龙虾壳炭 10.6 477.9 254.7 13.3 8.2 1 891.0 319.0 48.0 91.0 0.3 2.9 12.1 16.5 细叶榕炭 9.6 84.6 762.7 5.2 22.5 779.0 16.0 11.0 52.0 0.1 3.8 21.6 153.5  下载: 导出CSV
下载: 导出CSV
-
[1] 环境保护部, 国土资源部. 全国土壤污染状况调查公报(2014年4月17日)[J]. 环境教育, 2014(6): 8 − 10. Ministry of Environment Protection, Ministry of Natural Resources. National soil pollution survey bulletin (2014-04-17) [J]. Environmental Education, 2014(6): 8 − 10. [2] 郑越. 土壤重金属污染的现状及其治理[J]. 中国资源综合利用, 2019, 37(5): 114 − 116. ZHENG Yue. Current situation and treatment of heavy metal pollution in soil [J]. China Resources Comprehensive Utilization, 2019, 37(5): 114 − 116. [3] WANG Yangyang, LIU Yidan, ZHAN Wenhao, et al. Stabilization of heavy metal-contaminated soils by biochar: challenges and recommendations[J/OL]. Science of the Total Environment, 2020, 729: 139060[2022-01-20]. doi: 10.1016/j.scitotenv.2020.139060. [4] 张建云, 高才慧, 朱晖, 等. 生物质炭对土壤中重金属形态和迁移性的影响及作用机制[J]. 浙江农林大学学报, 2017, 34(3): 543 − 551. ZHANG Jianyun, GAO Caihui, ZHU Hui, et al. Mechanism and effects of biochar application on morphology and migration of heavy metals in contaminated soil [J]. Journal of Zhejiang A&F University, 2017, 34(3): 543 − 551. [5] YANG Xing, PAN He, SHAHEEN S M, et al. Immobilization of cadmium and lead using phosphorus-rich animal-derived and iron-modified plant-derived biochars under dynamic redox conditions in a paddy soil[J/OL]. Environment International, 2021, 156: 106628[2022-01-13]. doi: 10.1016/j.envint.2021.106628. [6] 李冬琴, 李冰倩, 梁静, 等. 硅和生物炭互作对空心菜-土壤系统中重金属生物有效性的影响[J]. 生态科学, 2021, 40(6): 99 − 105. LI Dongqin, LI Bingqian, LIANG Jing, et al. The interactive effects of silicon and biochar on heavy metal bioavailability in a water spinach-soil system [J]. Ecological Science, 2021, 40(6): 99 − 105. [7] 周雷, 嵇梦圆, 桑文静, 等. 稻草秸秆生物炭对土壤中重金属Cd和Pb的固化稳定化机制[J]. 化工环保, 2021, 41(5): 612 − 617. ZHOU Lei, JI Mengyuan, SANG Wenjing, et al. Solidification and stabilization mechanism of rice straw biochar to heavy metal Cd and Pb in soil [J]. Environmental Protection of Chemical Industry, 2021, 41(5): 612 − 617. [8] 刘郡. 城市厨余垃圾资源化回收利用现状分析与研究[J]. 资源节约与环保, 2021(10): 146 − 148. LIU Jun. Analysis and research on the current situation of urban kitchen waste recycling [J]. Resources Economization &Environmental Protection, 2021(10): 146 − 148. [9] 鞠艳艳, 丁元君, 邵前前, 等. 城市园林废弃物生物质炭性质及其应用潜力[J]. 林业科学, 2020, 56(8): 107 − 120. JU Yanyan, DING Yuanjun, SHAO Qianqian, et al. Properties and application potential of biochar from urban green wastes [J]. Scientia Silvae Sinicae, 2020, 56(8): 107 − 120. [10] 刘之欣, 郑森林, 方小山, 等. ENVI-met乔木模型对亚热带湿热地区细叶榕的模拟验证[J]. 北京林业大学学报, 2018, 40(3): 1 − 12. LIU Zhixin, ZHENG Senlin, FANG Xiaoshan, et al. Simulating validation of ENVI-met vegetation model to Ficus microcarpa in hot-humid region of subtropical zone [J]. Journal of Beijing Forestry University, 2018, 40(3): 1 − 12. [11] PENG Qian, NUNES L M, GREENFIELD B K, et al. Are Chinese consumers at risk due to exposure to metals in crayfish? A bioaccessibility-adjusted probabilistic risk assessment [J]. Environment International, 2016, 88: 261 − 268. [12] 戴志楠, 杨兴, 陈翰博, 等. 原始及铁改性生物质炭对污染土壤中As、Pb生物有效性和微生物群落结构的影响[J]. 环境科学学报, 2022, 42(6): 1 − 10. DAI Zhinan, YANG Xing, CHEN Hanbo, et al. Effect of raw and iron-modified biochars on the bioavailability of As and Pb and functional diversityof the microbial community in soils [J]. Acta Scientiae Circumstantiae, 2022, 42(6): 1 − 10. [13] 贝美容, 黄艳艳, 井玉丹, 等. 施用不同配比生物炭对橡胶树根系生长的短期响应[J]. 热带作物学报, 2019, 40(6): 1041 − 1045. BEI Meirong, HUANG Yanyan, JING Yudan, et al. Short-term effect of different rate of biochar application on rootdevelopment of Hevea brasiliensis [J]. Chinese Journal of Tropical Crops, 2019, 40(6): 1041 − 1045. [14] 鲁如坤. 土壤农业化学分析方法[M]. 北京: 中国农业科技出版社, 2000. LU Rukun. The Analysis Method of Soil Agricultural Chemistry[M]. Beijing: China Agricultural Science and Technology Press, 2000. [15] 包建平, 袁根生, 董方圆, 等. 生物质炭与秸秆施用对红壤有机碳组分和微生物活性的影响[J]. 土壤学报, 2020, 57(3): 721 − 729. BAO Jianping, YUAN Gensheng, DONG Fangyuan, et al. Effects of biochar application and straw returning on organic carbon fractionations and microbial activities in a red soil [J]. Acta Pedologica Sinica, 2020, 57(3): 721 − 729. [16] SUN Tao, XU Yingming, SUN Yuebing, et al. Crayfish shell biochar for the mitigation of Pb contaminated water and soil: characteristics, mechanisms, and applications[J/OL]. Environmental Pollution, 2021, 271: 116308[2022-01-18]. doi: 10.1016/j.envpol.2020.116308. [17] RECH A S, RECH J, CAPRARIO J, et al. Use of shrimp shell for adsorption of metals present in surface runoff [J]. Water Science and Technology, 2019, 79(12): 2221 − 2230. [18] PARK J H, WANG J J, XIAO R, et al. Effect of pyrolysis temperature on phosphate adsorption characteristics and mechanisms of crawfish char [J]. Journal of Colloid &Interface Science, 2018, 525: 143 − 151. [19] LIU Zhaoyun, DEMISIE W, ZHANG Mingkui, et al. Simulated degradation of biochar and its potential environmental implications [J]. Environmental Pollution, 2013, 179: 146 − 52. [20] BRUUN E W, HAUGGAARD-NIELSEN H, IBRAHIM N, et al. Influence of fast pyrolysis temperature on biochar labile fraction and short-term carbon loss in a loamy soil [J]. Biomass &Bioenergy, 2011, 35(3): 1182 − 1189. [21] LEE X J, LEE L Y, HIEW B Y Z, et al. Multistage optimizations of slow pyrolysis synthesis of biochar from palm oil sludge for adsorption of lead [J]. Bioresource Technology, 2017, 245: 944 − 953. [22] 聂天宏, 韩学博, 王海龙, 等. 不同种类生物质炭对植烟土壤保育及烤烟生长和品质的影响[J]. 水土保持学报, 2018, 32(6): 346 − 351, 358. NIE Tianhong, HAN Xuebo, WANG Hailong, et al. Effect of diffreent biochars on conservation, growth and quality of the flue-cured tobacco [J]. Journal of Soil and Water Conservation, 2018, 32(6): 346 − 351, 358. [23] XIAO Liang, YUAN Guodong, FENG Lirong, et al. Biochar to reduce fertilizer use and soil salinity for crop production in the Yellow River Delta [J]. Journal of Soil Science and Plant Nutrition, 2022, 22(2): 1478 − 1489. [24] van ZWIETEN L, KIMBER S, MORRIS S, et al. Effects of biochar from slow pyrolysis of papermill waste on agronomic performance and soil fertility [J]. Plant and Soil, 2010, 327(1/2): 235 − 246. [25] 曾爱, 廖允成, 张俊丽, 等. 生物炭对塿土土壤含水量、有机碳及速效养分含量的影响[J]. 农业环境科学学报, 2013, 32(5): 1009 − 1015. ZENG Ai, LIAO Yuncheng, ZHANG Junli, et al. Effects of biochar on soil moisture, organic carbon and available nutrient contents in manural loessial soils [J]. Journal of Agro-Environment Science, 2013, 32(5): 1009 − 1015. [26] 武春成, 李天来, 曹霞, 等. 添加生物炭对连作营养基质理化性质及黄瓜生长的影响[J]. 核农学报, 2014, 28(8): 1534 − 1539. WU Chuncheng, LI Tianlai, CAO Xia, et al. Physicochemical properties of nutrition medium and cucumber growth under continuous cropping [J]. Journal of Nuclear Agricultural Sciences, 2014, 28(8): 1534 − 1539. [27] 聂新星, 陈防. 生物炭对土壤钾素生物有效性影响的研究进展[J]. 中国土壤与肥料, 2016(2): 1 − 6. NIE Xinxing, CHEN Fang. Advances of the effects of biochar application on soil potassium bioavailability [J]. Soil and Fertilizer Sciences in China, 2016(2): 1 − 6. [28] DELUCA T, GUNDALE M J, MACKENZIE M D, et al. Biochar effects on soil nutrient transformations[M]// LEHMANN F, FOSEPH S. Biochar for Environmental Management: Science and Technology. London: Earthscan Ltd. , 2009: 251 − 270. [29] ANGST T E, SOHI S P. Establishing release dynamics for plant nutrients from biochar [J]. GCB Bioenergy, 2013, 5: 221 − 226. [30] 李丽, 王雪艳, 田彦芳, 等. 生物质炭对土壤养分及设施蔬菜产量与品质的影响[J]. 植物营养与肥料学报, 2018, 24(5): 1237 − 1244. LI Li, WANG Xueyan, TIAN Yanfang, et al. Effects of biochar on soil nutrients, yield and quality of vegetables [J]. Journal of Plant Nutrition and Fertilizers, 2018, 24(5): 1237 − 1244. [31] ASAI H, SAMSON B K, STEPHAN H M, et al. Biochar amendment techniques for upland rice production in Northern Laos (1) Soil physical properties, leaf SPAD and grain yield [J]. Field Crops Research, 2009, 111(1): 81 − 84. [32] 刘振刚, 夏宇, 孟芋含, 等. 生物质炭材料修复重金属污染土壤的研究进展: 修复机理及研究热点分析[J]. 环境工程学报, 2021, 15(4): 1140 − 1148. LIU Zhengang, XIA Yu, MENG Yuhan, et al. Research advances in biomass-based carbon materials for remediation of heavy metal contaminated soil: immobilization mechanism and analysis of related studies [J]. Chinese Journal of Environmental Engineering, 2021, 15(4): 1140 − 1148. [33] 钱钱, 杨兴, 郭明, 等. 生物质炭对土壤吸附Zn2+-DEP复合污染溶液中Zn2+的影响[J]. 浙江农林大学学报, 2019, 36(6): 1051 − 1061. QIAN Qian, YANG Xing, GUO Ming, et al. Adsorption of Zn2+ from a Zn2+-DEP (diethyl phthalate)composite solution using biochars in soil [J]. Journal of Zhejiang A&F University, 2019, 36(6): 1051 − 1061. [34] 徐炜杰, 郭佳, 赵敏, 等. 重金属污染土壤植物根系分泌物研究进展[J]. 浙江农林大学学报, 2017, 34(6): 1137 − 1148. XU Weijie, GUO Jia, ZHAO Min, et al. Research progress of soil plant root exudates in heavy metal contaminated soil [J]. Journal of Zhejiang A&F University, 2017, 34(6): 1137 − 1148. [35] 郝金才, 李柱, 吴龙华, 等. 铅镉高污染土壤的钝化材料筛选及其修复效果初探[J]. 土壤, 2019, 51(4): 752 − 759. HAO Jincai, LI Zhu, WU Longhua, et al. Preliminary study on cadmium and lead stabilization in soil highly polluted with heavy metals using different stabilizing agents [J]. Soils, 2019, 51(4): 752 − 759. [36] 郭荣荣, 黄凡, 易晓媚, 等. 混合无机改良剂对酸性多重金属污染土壤的改良效应[J]. 农业环境科学学报, 2015, 34(4): 686 − 694. GUO Rongrong, HUANG Fan, YI Xiaomei, et al. Remediation of multi-metals contaminated acidic soil by mixed inorganic amendments [J]. Journal of Agro-Environment Science, 2015, 34(4): 686 − 694. [37] 朱光耀, 何丽芝, 秦鹏, 等. 施用猪炭对土壤吸附Pb2+的影响[J]. 浙江农林大学学报, 2019, 36(3): 573 − 580. ZHU Guangyao, HE Lizhi, QIN Peng, et al. Dead pig-derived biochar treatments and soil adsorption of Pb2+ [J]. Journal of Zhejiang A&F University, 2019, 36(3): 573 − 580. [38] 温小情, 林亲铁, 肖荣波, 等. 镁基膨润土和水泥对砷铅复合污染土壤的钝化效能与机理研究[J]. 环境科学学报, 2020, 40(9): 3397 − 3404. WEN Xiaoqing, LIN Qintie, XIAO Rongbo, et al. Study on passivation efficiency and mechanism of Mg-bentonite and cement on As/Pb contaminated soil [J]. Acta Scientiae Circumstantiae, 2020, 40(9): 3397 − 3404. [39] 张迪, 丁爱芳. 组配钝化剂对镉铅复合污染土壤修复效果研究[J]. 农业环境科学学报, 2018, 37(12): 2718 − 2726. ZHANG Di, DING Aifang. Effects of combined passivating agents on remediation of Cd and Pb compound-contaminated soil [J]. Journal of Agro-Environment Science, 2018, 37(12): 2718 − 2726. [40] 陈新红, 叶玉秀, 潘国庆, 等. 杂交水稻不同器官重金属铅浓度与累积量[J]. 中国水稻科学, 2014, 28(1): 57 − 64. CHEN Xinhong, YE Yuxiu, PAN Guoqing, et al. Concentration and accumlation of lead in different organs of hybrid rice [J]. Chinese Journal of Rice Science, 2014, 28(1): 57 − 64. [41] JEFFERY S, VERHEIJEN F G A, ven der VELDEM, et al. A quantitative review of the effects of biochar application to soils on crop productivity using meta-analysis [J]. Agriculture,Ecosystems &Environment, 2011, 144(1): 175 − 187. [42] BAILEY V L, FANSLER S J, SMITH J L, et al. Reconciling apparent variability in effects of biochar amendment on soil enzyme activities by assay optimization [J]. Soil Biology &Biochemistry, 2011, 43(2): 296 − 301. [43] 张丽娜. 微生物菌肥和中微量元素在冀西北坝上地区萝卜上的应用研究[D]. 张家口: 河北北方学院, 2017. ZHANG Lina. Application of Microbial Bacterial Frtilizer and Medium and Trace Elements on Radish in the Bashang Area of Northwest Hebei Province[D]. Zhangjiakou: Heibei North University, 2017. [44] YANG Xing, LU Kouping, MCGROUTHER K, et al. Bioavailability of Cd and Zn in soils treated with biochars derived from tobacco stalk and dead pigs [J]. Journal of Soils and Sediments, 2017, 17(3): 751 − 762. [45] LEBRUN M, MIARD F, NANDILLON R, et al. Biochar application rate: improving soil fertility and Linum usitatissimum growth on an arsenic and lead contaminated technosol [J]. International Journal of Environmental Research, 2021, 15(1): 125 − 134. [46] 刘晓霞, 陈红金, 虞轶俊. 骨炭的土壤改良培肥和小萝卜增产提质效应研究[J]. 中国土壤与肥料, 2021(5): 150 − 155. LIU Xiaoxia, CHEN Hongjin, YU Yijun. Studies on the effects of bone-biochar on soil quality promotion and radish production improvement [J]. Soil and Fertilizer Sciences in China, 2021(5): 150 − 155. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20220182
 点击查看大图
点击查看大图

计量
- 文章访问数: 1696
- HTML全文浏览量: 552
- PDF下载量: 89
- 被引次数: 0


