




-
土壤呼吸是陆地生态系统向大气排放二氧化碳(CO2)的主要途径,在调节生态系统碳循环中发挥着重要作用[1−2]。作为陆地生态系统的主体,森林土壤汇集了全球土壤碳库的73%[3],森林土壤呼吸研究成为陆地生态系统碳循环的研究热点。森林间伐是森林经营的重要措施之一,对促进人工林生长,改善碳固存能力有积极作用,通过改变林地土壤微气候、微生物群落以及根系生长等因子引起森林土壤呼吸变化[4−6]。因此,研究森林间伐对土壤呼吸及其影响机制对评估区域土壤碳循环有重要意义。杉木Cunninghamia lanceolata是中国特有速生丰产的优质树种,其人工林面积高达1 096万 hm2,在缓解木材需求压力、支持天然林保护等生态工程实施方面作用突出[7−9]。杉木人工林因其巨大的固碳潜力,在缓解气候变化上的作用不容忽视[10−11]。然而,由于杉木人工林长期多代连作,引起林地生产力下降、土壤退化,形成了大面积的低质低效杉木林[12]。森林间伐对杉木林土壤呼吸速率的响应结果差异较大。TIAN等[13]研究表明:杉木林土壤呼吸速率随间伐后恢复年限的增加先升高后降低,最终恢复到采伐前水平。LI等[14]研究发现:间伐后第1年杉木林土壤呼吸速率升高,其中异养呼吸增加是土壤呼吸增加的主要原因,但间伐对土壤呼吸的影响随时间推移而减弱。WANG等[15]研究表明:土壤呼吸随间伐强度的增加而增加。丁驰等[16]研究发现:杉木林轻度间伐的土壤呼吸速率显著高于中度和重度间伐处理。然而,探讨间伐对杉木林土壤呼吸的响应时多考虑土壤温湿度的影响,而其他环境因子对土壤呼吸速率的影响仍有待进一步研究。此外,间伐作业对高密度萌生杉木林的土壤呼吸速率影响的研究较少。综上所述,本研究以浙北杉木人工林为研究对象,比较不同间伐强度下森林土壤呼吸速率及其主要影响因子,为杉木林经营及碳汇管理提供科学依据。
-
研究区位于浙江省临安市於潜镇泗州村(30°14′N,119°25′E),该区为典型亚热带季风气候,年均气温为16.2 ℃,最高气温在7—8月,最低气温在1月,年均降水量为1 470.2 mm,降水天数为182.0 d。海拔为40~100 m,样地主要土壤类型为黄壤。杉木林下灌木主要有山茶Camellia japonica、土茯苓Camellia oleifera、菝葜Smilax china等;草本主要有山莓Rubus corchorifolius、金毛耳草Hedyotis Chrysotricha、芒属Miscanthus类植物等。
-
试验地为杉木萌生纯林,该林分是2006年遭受火灾后采伐形成的萌生林,截至2017年12月未进行经营活动。该样地林分密度为4 200株·hm−2,林龄为11 a,林分平均胸径为7.3 cm,平均树高为5.2 m。于2017年12月进行采伐和每木检尺,采用随机区组设计在坡度和坡位相近的杉木纯林设置3种间伐处理,分别为对照(间伐0%)、中度间伐(间伐45%)和重度间伐(间伐70%),每种处理设置3个重复,共设置了9块20 m×20 m的标准样地,样地基本情况见表1。样地0~20 cm土层土壤理化性质:土壤容重为1.07 g·cm−3,土壤有机碳为36.40 g·kg−1,土壤全氮为1.31 g·kg−1,土壤碱解氮为39.51 mg·kg−1,土壤有效磷为1.14 mg·kg−1,土壤速效钾为64.52 mg·kg−1,pH 5.03。
表 1 杉木林样地基本情况
Table 1. Basic conditions of the plots
间伐处理 林分密度/(株·hm−2) 平均胸径/cm 平均树高/m 郁闭度/% 伐前 伐后 伐前 伐后 伐前 伐后 伐前 伐后 对照 4 188±45 4 188±45 7.6±0.1 7.6±0.1 5.0±0.1 5.0±0.1 95 95 中度 4 140±225 1 833±55 7.3±0.1 7.8±0.4 5.1±0.1 5.3±0.2 95 62 重度 4 317±114 1 103±48 7.1±0.1 7.4±0.2 4.8±0.2 5.0±0.3 95 38 于2018年2月至2020年1月,采用静态箱-气象色谱法进行土壤呼吸速率监测,每个样地安置2个30 cm×30 cm×30 cm的PVC静态箱(下坡处静态箱基座没入土壤5 cm),共18个静态箱。每月中旬9:00—11:00采集气体样品。用25 mL注射器分别于密封后的0、10、20、30 min采集气体样品,注入密封的真空气瓶中,带回实验室,48 h内用Agilent 7890气相色谱仪完成测定。
每月采气时,测定静态箱附近土壤5 cm处温度,并采集5 cm处土壤样品使用烘干法测定土壤含水率[16]。每季度采集0~20 cm土层土壤样品,过2 mm筛后,分别风干后置于4 ℃冰箱保存。土壤水溶性有机碳采用水浸提法测定[17],土壤易氧化有机碳采用高锰酸钾氧化法测定[18],土壤微生物生物量碳采用氯仿熏蒸浸提法测定[19],土壤pH采用电位法测定[20]。
-
使用模型拟合土壤呼吸和土壤温湿度的关系,同时估计温度敏感性系数(Q10)。
$$ \begin{array}{l} {R}_{\mathrm{s}}=a{{\rm{e}}}^{bT} \text{;} \\ {R}_{\mathrm{s}}=a{M}^{2}+bM+c \text{;}\\ {Q}_{10}={{\rm{e}}}^{10b} 。 \end{array} $$ 其中:Rs为土壤呼吸速率(mg·m−2·h−1),T为土壤5 cm土层处温度(℃),M为土壤表层含水率(%),a、b和c为系数参数。
采用SPSS 22.0进行数据统计分析。采用重复观测方差分析(repeated measures ANOVA)检验杉木林土壤呼吸速率与土壤温湿度的差异性,采用单因素方差分析(one-way ANOVA)探讨不同间伐处理下土壤呼吸速率和土壤环境因子差异。利用线性回归分析土壤呼吸速率与土壤环境因子的关系。
-
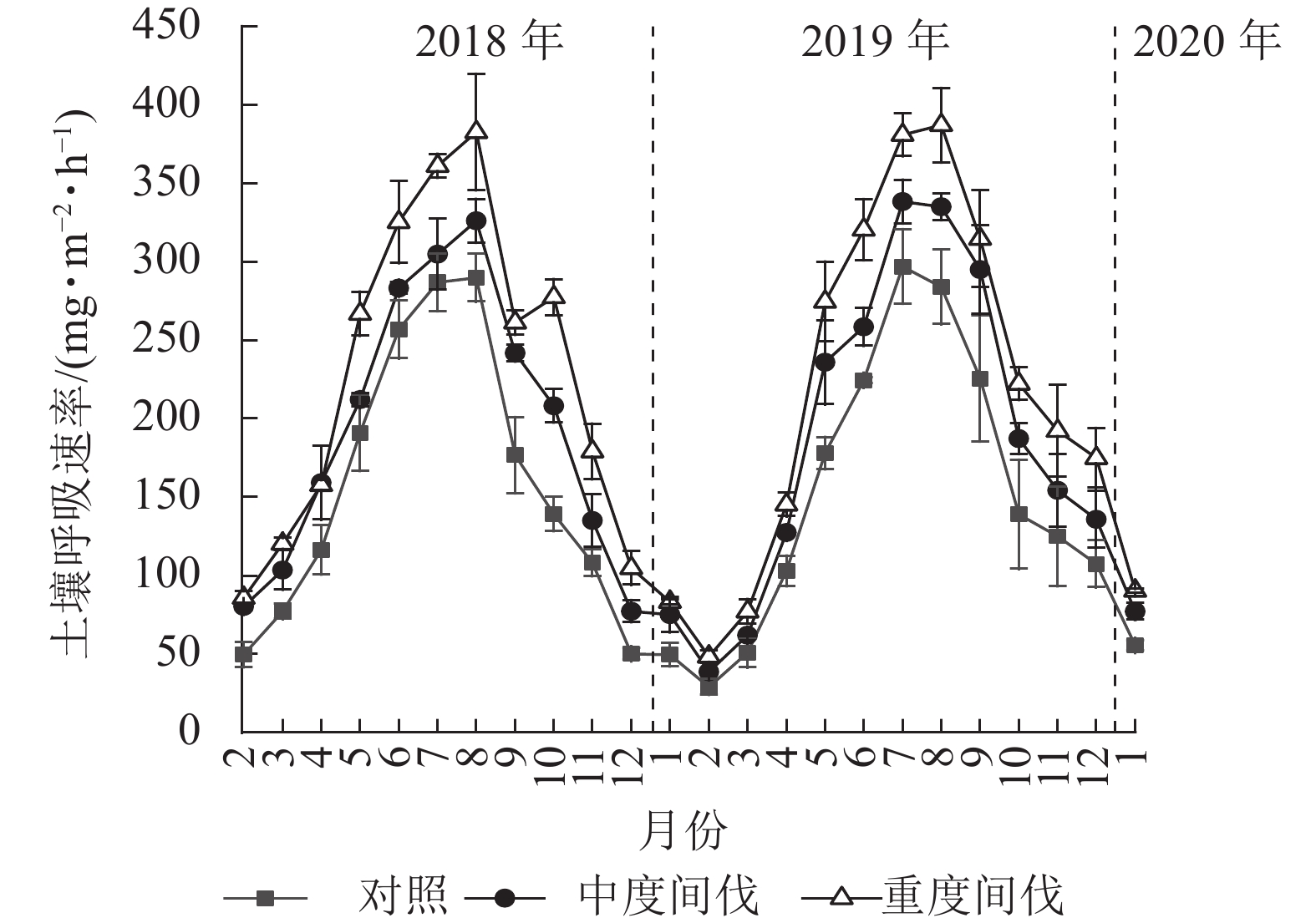
由图1可知:不同间伐处理的杉木林土壤呼吸速率均呈现明显的季节动态,最高值出现在7—8月,最低值出现在1—2月。从月动态来看,5月开始各间伐处理土壤呼吸速率出现较大差异。对照、中度、重度间伐样地土壤呼吸速率分别为29.09~297.00、38.66~338.31、48.36~387.87 mg·m−2·h−1。间伐显著增加了杉木林土壤呼吸速率(P<0.05),且随着间伐强度的增加而增加。与对照相比,中度和重度间伐土壤呼吸速率分别增加了23.30%、44.94%(表2)。从年际变化看,各间伐处理对土壤呼吸速率无明显差异。
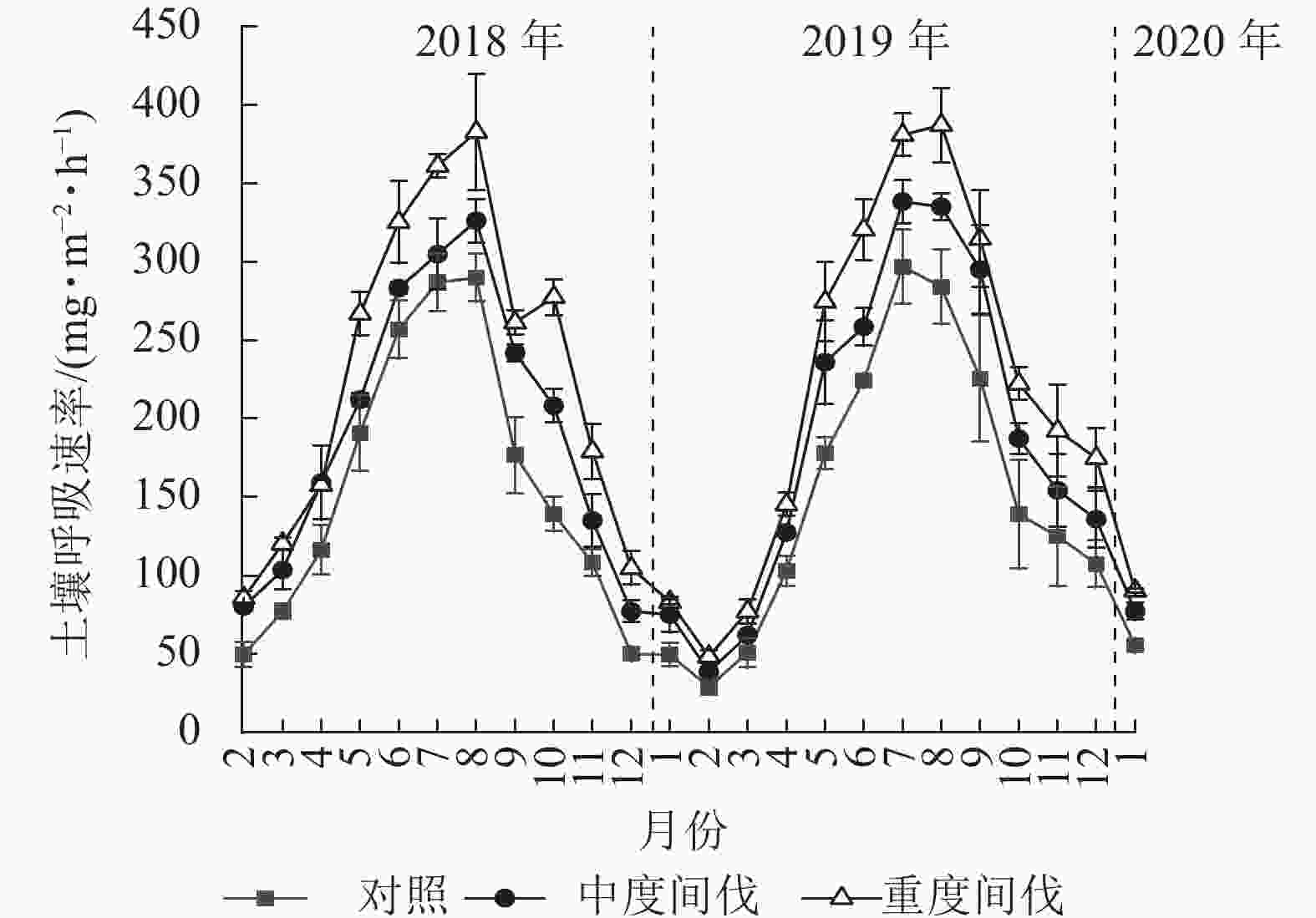
图 1 不同处理杉木林土壤呼吸速率的月动态
Figure 1. Monthly dynamic of soil respiration rate in C. lanceolata plantation under different thinning treatments
表 2 不同处理杉木林土壤呼吸速率及环境因子的年平均值
Table 2. Annual average values of soil respiration rate and environmental factors in C. lanceolata plantation under different thinning treatments
时间 间伐处理 土壤呼吸速率/
(mg·m−2·h−1)土壤温度/℃ 含水率/% 水溶性有机碳/
(mg·kg−1)微生物生物量碳/
(mg·kg−1)易氧化有机碳/
(mg·g−1)间伐后第1年 对照 149.32±2.92 c 15.00±0.19 b 27.54±0.89 a 96.80±2.18 b 266.21±6.84 c 10.74±0.66 b 中度 183.89±5.95 b 15.96±0.10 a 30.49±1.37 a 109.14±4.85 a 295.60±8.19 b 13.30±.0.81 a 重度 217.15±1.79 a 16.16±0.23 a 29.74±1.52 a 119.51±6.16 a 331.07±8.96 a 14.34±0.45 a 间伐后第2年 对照 151.61±6.01 c 15.67±0.23 b 21.47±0.73 a 101.83±1.57 c 244.72±9.71 b 9.28±0.32 b 中度 187.13±6.10 b 16.65±0.22 a 24.85±1.61 a 112.92±3.10 b 282.76±7.20 a 11.69±0.21 a 重度 219.02±2.18 a 16.91±0.29 a 23.39±2.31 a 122.21±2.94 a 307.12±14.35 a 11.73±0.36 a 平均 对照 150.46±1.61 c 15.34±0.15 b 24.51±0.62 a 99.31±1.05 c 255.47±7.01 c 10.01±0.31 b 中度 185.51±5.51 b 16.30±0.15 a 27.67±1.49 a 111.03±3.95 b 289.18±7.29 b 12.49±0.32 a 重度 218.09±1.44 a 16.53±0.26 a 26.56±1.91 a 120.86±3.80 a 319.09±9.62 a 13.04±0.40 a 说明:数值为平均值±标准误。不同字母表示不同处理间差异显著(P<0.05)。间伐后第1年为2018年2月至2019年1月,间伐后第2年为2019年2月至2020年1月。 -
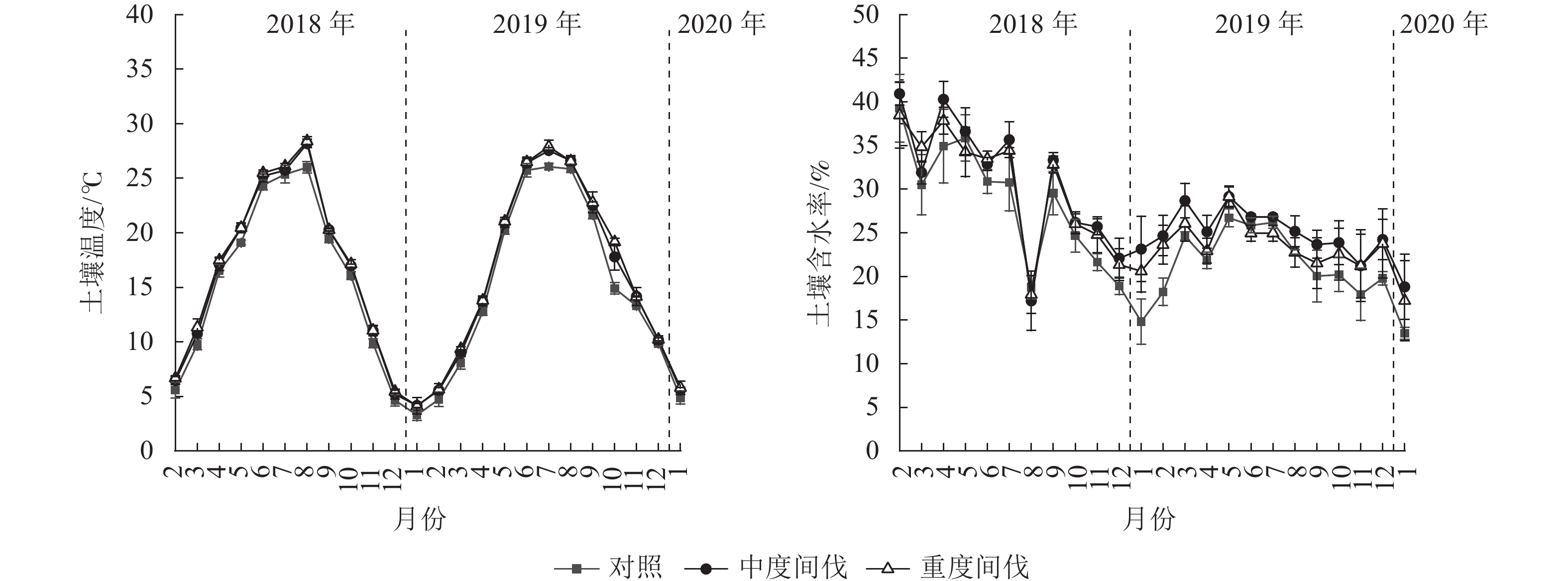
各间伐处理的土壤温度表现为春冬季低,夏季高的单峰曲线(图2),间伐处理样地均显著高于对照(P<0.05),与对照相比,中度和重度间伐处理的土壤温度分别增加了6.31%和7.81%(表2)。土壤含水率在间伐第1年(2018年2月至2019年1月)呈现降低趋势,第2年(2019年2月至2020年1月)呈先升高后下降的波动趋势,但各处理土壤含水量差异不显著(P>0.05)。
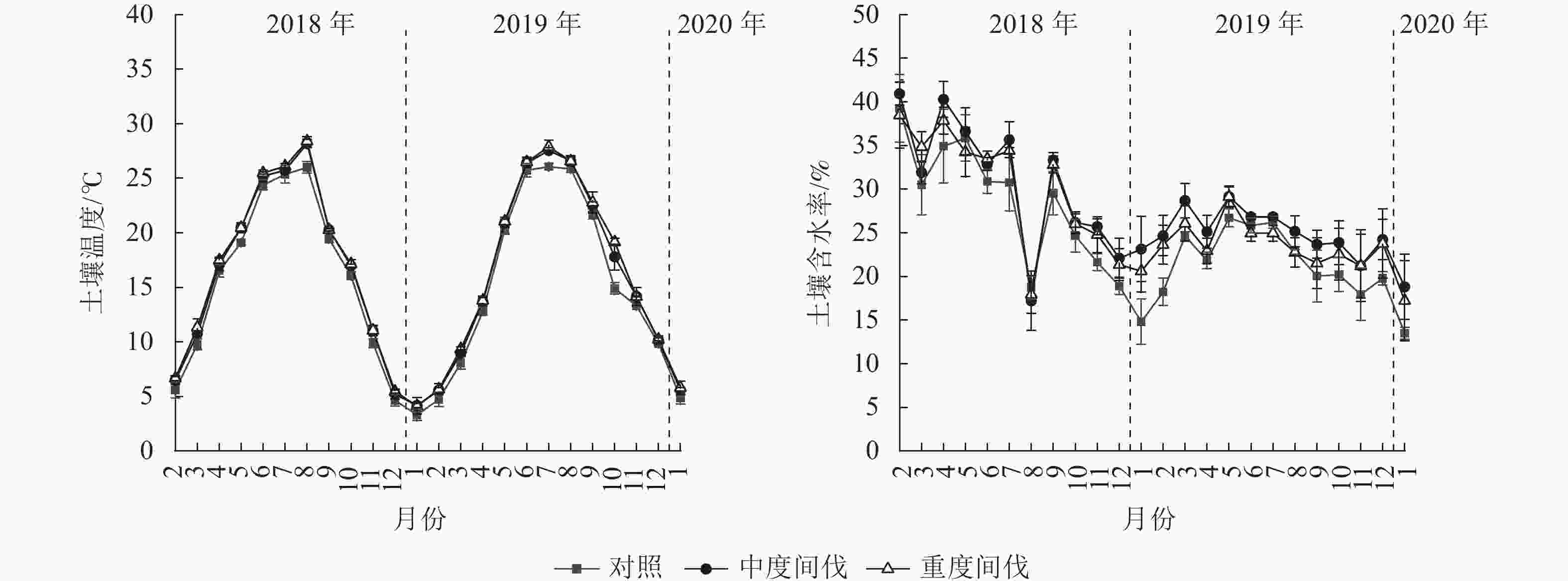
图 2 不同处理杉木林土壤温湿度的动态变化
Figure 2. Dynamics of soil temperature and moisture in C. lanceolata plantation under different treatments
-
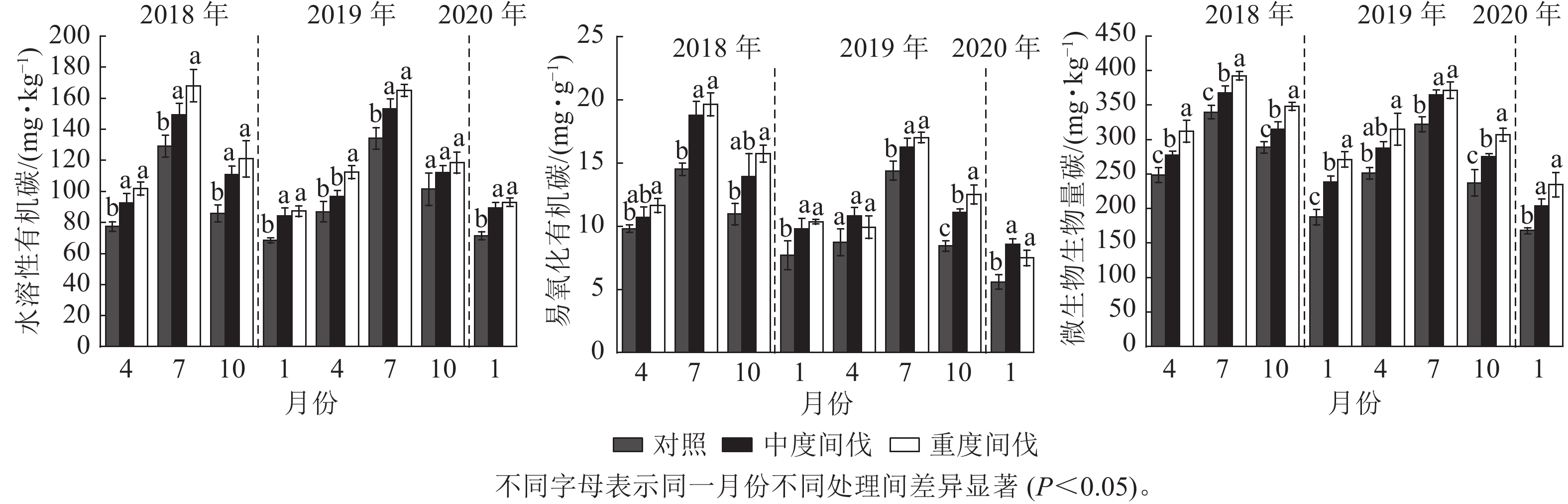
由图3可知:各处理土壤水溶性有机碳质量分数7月最高,1月最低。与对照相比,除间伐第2年的4和10月外,中度和重度间伐处理下土壤水溶性有机碳质量分数均显著增加(P<0.05),分别增加了11.80%和21.70%(表2),间伐处理之间差异不显著。
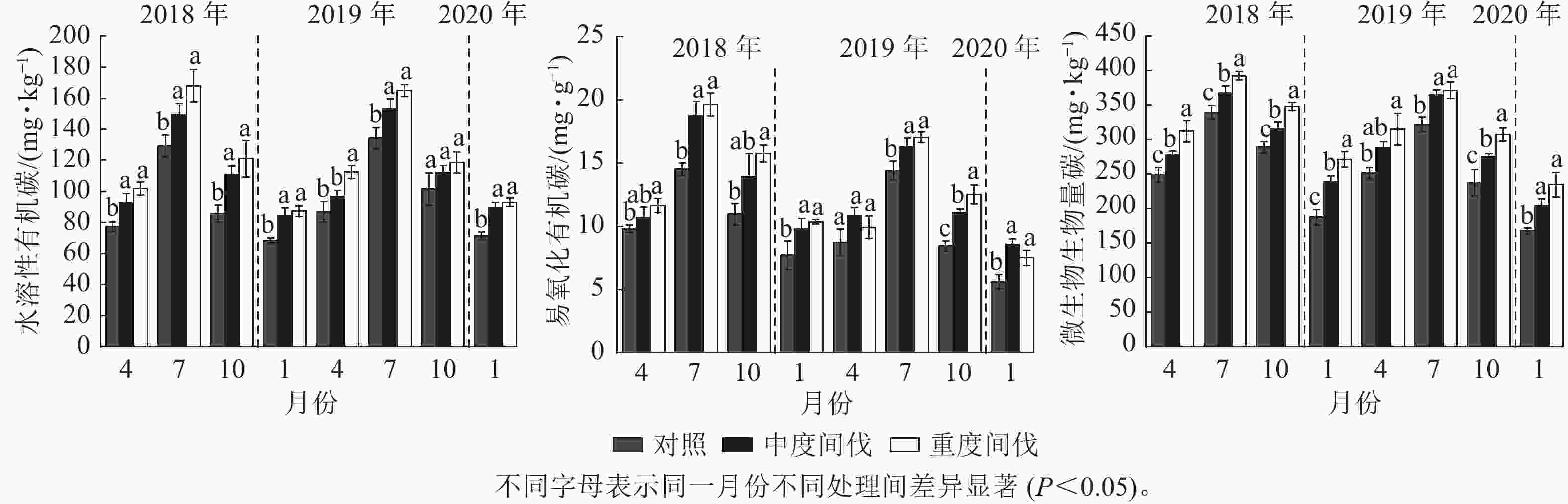
图 3 不同处理杉木林土壤活性有机碳的季节变化
Figure 3. Seasonal variation of soil active organic carbon in C. lanceolata forest under different treatments
除间伐第2年的4月外,重度间伐处理的土壤易氧化有机碳显著增加(P<0.05);除间伐第1年的4和10月、第2年的4月外,中度间伐处理的土壤易氧化有机碳显著增加(P<0.05)。整体上表现为间伐增加土壤易氧化有机碳质量分数,且与间伐强度无关。与对照相比,中度和重度间伐的土壤易氧化有机碳质量分数分别增加了24.83%和30.27%。
间伐后第1年,与对照相比,随着间伐强度的增加,微生物生物量碳质量分数显著增加(P<0.05)。间伐后第2年除4月外,间伐处理显著增加了微生物生物量碳质量分数(P<0.05)。整体上看,间伐增加了土壤微生物生物量碳,且不同间伐处理间差异显著(P<0.05),中度和重度间伐分别增加了14.01%和24.91%(表2)。
-
从表3可见:间伐后第1年和第2年各处理土壤呼吸速率与土壤温度均呈显著指数相关(P<0.01)。对照的土壤呼吸速率仅间伐第2年与土壤含水率呈正相关(R2=0.268,P<0.01),而间伐处理的土壤呼吸速率与土壤含水率无显著关系。因此,杉木林土壤呼吸速率随着土壤温度的升高而增加,几乎不受土壤含水率的影响。Q10反映土壤呼吸速率对土壤温度变化的敏感性。研究期间,对照、中度、重度间伐的Q10分别为2.10、1.86、1.84,间伐降低了杉木林土壤呼吸的敏感性。
表 3 土壤呼吸速率与土壤温湿度回归模型以及温度敏感性系数(Q10)
Table 3. Regression models of soil respiration rate with soil temperature, moisture and temperature sensitivity (Q10)
时间 间伐处理 土壤温度 土壤湿度 Q10 a b R2 a b c R2 间伐后第1年 对照 0.252 0.077 0.934** −32.151 18.368 −1.481 0.151 2.16±0.09 中度 0.405 0.059 0.941** 1.251 −1.168 1.393 0.004 1.80±0.05 重度 0.491 0.057 0.882** −23.721 13.159 −0.322 0.034 1.77±0.07 间伐后第2年 对照 0.263 0.072 0.858** 16.971 0.095 0.129 0.268** 2.05±0.12 中度 0.351 0.066 0.884** −56.700 30.036 −2.710 0.067 1.93±0.10 重度 0.408 0.066 0.904** −41.085 21.324 −1.283 0.046 1.93±0.08 总 对照 0.262 0.074 0.904** −35.500 20.339 −1.730 0.192 2.10±0.08 中度 0.377 0.062 0.907** −11.228 6.651 0.238 0.011 1.86±0.06 重度 0.448 0.061 0.891** −26.646 14.972 −0.598 0.040 1.84±0.06 说明:间伐后第1年为2018年2月至2019年1月,间伐后第2年为2019年2月至2020年1月,总表示2018年2月到2020年1月总计。**表示相关极显著(P<0.01)。 各处理杉木林土壤呼吸速率与土壤温度、土壤水溶性有机碳、土壤微生物生物量碳和土壤易氧化有机碳均呈极显著正相关(P<0.01),而与土壤含水率不相关(表4)。由表5可知:土壤温度是影响杉木林土壤呼吸速率的最主要因子。除中度间伐外,土壤易氧化有机碳是影响土壤呼吸速率的次要因子。土壤水溶性有机碳是中度间伐处理下影响土壤呼吸速率的次要因子。
表 4 土壤呼吸速率与环境因子的相关性
Table 4. Correlation between soil respiration rate and environmental factors
间伐处理 水溶性有机碳 易氧化有机碳 微生物生物量碳 土壤温度 土壤含水率 对照 0.838** 0.903** 0.860** 0.944** 0.201 中度 0.895** 0.850** 0.915** 0.957** 0.028 重度 0.864** 0.897** 0.881** 0.960** 0.014 总计 0.874** 0.885** 0.880** 0.924** 0.106 说明:双尾检验。**表示极显著相关(P<0.01)。 表 5 土壤呼吸速率与环境因子的多元回归模型
Table 5. Multiple regression model of soil respiration rate with environmental factors
间伐处理 模型 n F R2 P 对照 Rs=0.947xT 24 192.818 0.893 <0.001 Rs=0.689 xT +0.291 xPOXC 24 113.839 0.908 <0.001 Rs =0.890 xT +0.486 xPOXC +0.406 xMBC 24 92.408 0.923 <0.001 中度 Rs =0.967 xT 24 321.305 0.933 <0.001 Rs =0.724 xT +0.293 xWSOC 24 268.603 0.959 <0.001 重度 Rs =0.960 xT 24 261.820 0.919 <0.001 Rs =0.681 xT +0.343 xPOXC 24 267.618 0.959 <0.001 总计 Rs =0.935 xT 72 489.252 0.875 <0.001 Rs =0.630 xT +0.383 xPOXC 72 447.716 0.928 <0.001 Rs =0.580 xT +0.278 xPOXC +0.167 xWSOC 72 322.352 0.934 <0.001 说明:Rs为土壤呼吸速率;xT为土壤温度;xPOXC为土壤易氧化有机碳;xWSOC为土壤水溶性有机碳;xMBC为土壤微生物生物量碳。 -
森林间伐降低了林分密度,对土壤产生强烈扰动,改变了林分结构和土壤理化性质,进而影响土壤呼吸速率[21]。因间伐强度、森林类型、地表植被恢复和气候条件的不同对土壤呼吸影响差异较大,间伐对土壤呼吸速率可能增加、抑制或无影响[22−24]。本研究表明:间伐增加土壤呼吸速率,且随着间伐强度的增加而增加,这与前人在美洲黑杨Populus deltoides林、杉木人工林以及油松Pinus tabuliformis人工林的研究结果一致[15, 25−26]。这是因为间伐降低了林分郁闭度,更多的太阳辐射通过林窗到达地面,林地土壤温度升高,促进土壤有机质的分解和土壤微生物异养呼吸[27]。本研究中,中度和重度间伐处理的土壤温度显著高于对照样地,且各处理土壤温度均与土壤呼吸速率呈显著正相关,这与前人在亚热带杉木林和落叶云杉Picea asperata林的研究结果一致[28−29]。土壤温度作为土壤呼吸速率的主导因子,与土壤呼吸速率呈显著指数关系。随着间伐时间的推移,土壤温度呈现季节性升高和降低,影响林地植物生长代谢的周期性变化,土壤呼吸速率表现为夏季高冬季低的单峰曲线[30]。多元逐步回归结果也表明:土壤温度是影响土壤呼吸的主要因素。本研究中土壤含水率对土壤呼吸速率无显著影响。可能是本研究区降水充沛,土壤水分在整个试验过程变化有限,土壤温度对土壤呼吸变化的强烈影响可能掩盖土壤含水率的影响[31]。另外,林木移除虽然减少了植物的蒸腾散失,但间伐后林地太阳辐射和土壤温度的增加可能促进土壤水分蒸发,减弱土壤水分对土壤呼吸的响应[32]。与对照林分相比,间伐区郁闭度减小,促进了林下植被的恢复进而增加林地土壤根系的自养呼吸[26]。土壤活性有机碳是土壤有机碳中易被生物直接利用转化、最活跃、易氧化的组分,土壤CO2的排放依赖于土壤有机化合物的转化[33]。本研究中,间伐后土壤水溶性有机碳、微生物生物量碳和易氧化有机碳显著增加,且杉木林土壤呼吸速率与土壤各活性有机碳组分均呈显著正相关,这与在杉木林和云南松Pinus yunnanensis林的研究结果一致[34−35]。不同间伐强度的土壤活性有机碳差异是影响土壤呼吸速率的重要原因,间伐初期采伐剩余物输入和死根分解增加了微生物可分解的碳源,林下植被的快速恢复导致植物根系分泌物增多,进而增加了土壤水溶性有机碳质量分数[36−37]。另外,随着采伐后土壤温度的升高,土壤微生物活性提高,加快土壤有机质的矿化,土壤活性有机碳质量分数增加,进而促进土壤呼吸速率增加[38]。土壤呼吸是一系列酶促反应的过程体现,夏季林下适宜的水热环境以及丰富的碳源刺激了土壤微生物活性,这可能是本研究中土壤活性有机碳质量分数夏季较高的原因[39−40]。多元逐步回归分析表明:土壤水溶性有机碳和易氧化有机碳分别是中度和重度间伐的次要影响因子,表明随着间伐对林地干扰程度的增加,影响土壤CO2排放的因子重要性可能发生改变。
Q10作为评估土壤呼吸对全球变暖响应的重要参数,因地理位置、生态系统类型和土壤微生物等不同而不同[16]。在本研究中,与对照相比,间伐处理明显降低了Q10,这与人工油松林和杉阔混交林的研究结果一致[41−42]。研究表明:根系自养呼吸的Q10显著高于异养呼吸[43],Q10的降低可能是间伐后土壤保留的根系减少,自养呼吸减弱导致的[44]。
-
本研究表明:间伐处理显著增加了杉木人工林土壤呼吸速率,且随着间伐强度的增加而增加。间伐处理显著降低土壤Q10。土壤温度是影响土壤呼吸速率的主导因子,土壤活性有机碳是影响土壤呼吸速率的重要因子。
Short-term effects of different thinning intensities on soil respiration rate in the Cunninghamia lanceolata plantation
-
摘要:
目的 比较不同间伐强度下杉木Cunninghamia lanceolata人工林土壤呼吸速率,解释影响土壤呼吸速率的主要因子,为森林经营及碳汇管理提供科学依据。 方法 采用随机区组设计,设置对照(间伐0%)、中度间伐(间伐45%)、重度间伐(间伐70%) 3种间伐处理,采用静态箱-气象色谱法对杉木人工林土壤呼吸速率进行短期原位监测。 结果 间伐显著增加了杉木林土壤呼吸速率(P<0.05),与对照相比,中度和重度间伐的土壤呼吸速率分别增加了23.30%和44.94%。土壤呼吸速率与5 cm土壤温度呈显著指数相关,而与土壤含水量无关。不同间伐处理下,土壤呼吸温度敏感性系数(Q10)为1.77~2.16,对照处理下Q10最高,间伐降低了杉木林土壤呼吸的温度敏感性。杉木人工林土壤呼吸速率与土壤水溶性有机碳、微生物生物量碳和易氧化有机碳呈显著正相关(P<0.01)。 结论 间伐初期,间伐对杉木林土壤呼吸速率有促进作用,且随着间伐强度的增加而增加。土壤温度是土壤呼吸速率变化的主要影响因子,土壤活性有机碳是重要因子。图3表5参44 Abstract:Objective This study, with a comparison of the differences in soil respiration rates of Cunninghamia lanceolata plantations under different thinning intensities, is aimed to investigate the main factors affecting soil respiration rates, so as to provide a scientific basis for forest management and carbon sink management. Method With a randomized block design, three thinning treatments, namely control (thinning 0%), moderate thinning (thinning 45%) and heavy thinning (thinning 70%) were arranged before short-term in-situ monitoring was conducted of soil respiration rates in C. lanceolata plantations using static box-meteorological chromatography. Result Thinning significantly increased the soil respiration rate in C. lanceolata forest, with an increase of 23.30% and 44.94% for the ones treated with moderate and heavy thinning respectively. Soil respiration rate was exponentially correlated with soil temperature at 5 cm depth, but not with soil water content. The temperature sensitivity coefficient (Q10) of soil respiration ranged from 1.77 to 2.16 under different thinning treatments, with the Q10 being the highest under the control treatment, and thinning decreased the temperature sensitivity of soil respiration in C. lanceolata stands. Soil respiration rate of C. lanceolata plantation was positively correlated with soil water-soluble organic carbon, microbial biomass carbon and permanganate-oxidizable carbon (P<0.01). Conclusion Thinning promoted the soil respiration rate of C. lanceolata forest which increased with the increase of thinning intensity. Soil temperature is the main factor affecting the change in soil respiration rate, with soil active organic carbon being an important factor. [Ch, 3 fig. 5 tab. 44 ref.] -
图 1 不同处理杉木林土壤呼吸速率的月动态
Figure 1 Monthly dynamic of soil respiration rate in C. lanceolata plantation under different thinning treatments
图 2 不同处理杉木林土壤温湿度的动态变化
Figure 2 Dynamics of soil temperature and moisture in C. lanceolata plantation under different treatments
图 3 不同处理杉木林土壤活性有机碳的季节变化
Figure 3 Seasonal variation of soil active organic carbon in C. lanceolata forest under different treatments
表 1 杉木林样地基本情况
Table 1. Basic conditions of the plots
间伐处理 林分密度/(株·hm−2) 平均胸径/cm 平均树高/m 郁闭度/% 伐前 伐后 伐前 伐后 伐前 伐后 伐前 伐后 对照 4 188±45 4 188±45 7.6±0.1 7.6±0.1 5.0±0.1 5.0±0.1 95 95 中度 4 140±225 1 833±55 7.3±0.1 7.8±0.4 5.1±0.1 5.3±0.2 95 62 重度 4 317±114 1 103±48 7.1±0.1 7.4±0.2 4.8±0.2 5.0±0.3 95 38  下载: 导出CSV
下载: 导出CSV
表 2 不同处理杉木林土壤呼吸速率及环境因子的年平均值
Table 2. Annual average values of soil respiration rate and environmental factors in C. lanceolata plantation under different thinning treatments
时间 间伐处理 土壤呼吸速率/
(mg·m−2·h−1)土壤温度/℃ 含水率/% 水溶性有机碳/
(mg·kg−1)微生物生物量碳/
(mg·kg−1)易氧化有机碳/
(mg·g−1)间伐后第1年 对照 149.32±2.92 c 15.00±0.19 b 27.54±0.89 a 96.80±2.18 b 266.21±6.84 c 10.74±0.66 b 中度 183.89±5.95 b 15.96±0.10 a 30.49±1.37 a 109.14±4.85 a 295.60±8.19 b 13.30±.0.81 a 重度 217.15±1.79 a 16.16±0.23 a 29.74±1.52 a 119.51±6.16 a 331.07±8.96 a 14.34±0.45 a 间伐后第2年 对照 151.61±6.01 c 15.67±0.23 b 21.47±0.73 a 101.83±1.57 c 244.72±9.71 b 9.28±0.32 b 中度 187.13±6.10 b 16.65±0.22 a 24.85±1.61 a 112.92±3.10 b 282.76±7.20 a 11.69±0.21 a 重度 219.02±2.18 a 16.91±0.29 a 23.39±2.31 a 122.21±2.94 a 307.12±14.35 a 11.73±0.36 a 平均 对照 150.46±1.61 c 15.34±0.15 b 24.51±0.62 a 99.31±1.05 c 255.47±7.01 c 10.01±0.31 b 中度 185.51±5.51 b 16.30±0.15 a 27.67±1.49 a 111.03±3.95 b 289.18±7.29 b 12.49±0.32 a 重度 218.09±1.44 a 16.53±0.26 a 26.56±1.91 a 120.86±3.80 a 319.09±9.62 a 13.04±0.40 a 说明:数值为平均值±标准误。不同字母表示不同处理间差异显著(P<0.05)。间伐后第1年为2018年2月至2019年1月,间伐后第2年为2019年2月至2020年1月。
下载: 导出CSV
表 3 土壤呼吸速率与土壤温湿度回归模型以及温度敏感性系数(Q10)
Table 3. Regression models of soil respiration rate with soil temperature, moisture and temperature sensitivity (Q10)
时间 间伐处理 土壤温度 土壤湿度 Q10 a b R2 a b c R2 间伐后第1年 对照 0.252 0.077 0.934** −32.151 18.368 −1.481 0.151 2.16±0.09 中度 0.405 0.059 0.941** 1.251 −1.168 1.393 0.004 1.80±0.05 重度 0.491 0.057 0.882** −23.721 13.159 −0.322 0.034 1.77±0.07 间伐后第2年 对照 0.263 0.072 0.858** 16.971 0.095 0.129 0.268** 2.05±0.12 中度 0.351 0.066 0.884** −56.700 30.036 −2.710 0.067 1.93±0.10 重度 0.408 0.066 0.904** −41.085 21.324 −1.283 0.046 1.93±0.08 总 对照 0.262 0.074 0.904** −35.500 20.339 −1.730 0.192 2.10±0.08 中度 0.377 0.062 0.907** −11.228 6.651 0.238 0.011 1.86±0.06 重度 0.448 0.061 0.891** −26.646 14.972 −0.598 0.040 1.84±0.06 说明:间伐后第1年为2018年2月至2019年1月,间伐后第2年为2019年2月至2020年1月,总表示2018年2月到2020年1月总计。**表示相关极显著(P<0.01)。
下载: 导出CSV
表 4 土壤呼吸速率与环境因子的相关性
Table 4. Correlation between soil respiration rate and environmental factors
间伐处理 水溶性有机碳 易氧化有机碳 微生物生物量碳 土壤温度 土壤含水率 对照 0.838** 0.903** 0.860** 0.944** 0.201 中度 0.895** 0.850** 0.915** 0.957** 0.028 重度 0.864** 0.897** 0.881** 0.960** 0.014 总计 0.874** 0.885** 0.880** 0.924** 0.106 说明:双尾检验。**表示极显著相关(P<0.01)。
下载: 导出CSV
表 5 土壤呼吸速率与环境因子的多元回归模型
Table 5. Multiple regression model of soil respiration rate with environmental factors
间伐处理 模型 n F R2 P 对照 Rs=0.947xT 24 192.818 0.893 <0.001 Rs=0.689 xT +0.291 xPOXC 24 113.839 0.908 <0.001 Rs =0.890 xT +0.486 xPOXC +0.406 xMBC 24 92.408 0.923 <0.001 中度 Rs =0.967 xT 24 321.305 0.933 <0.001 Rs =0.724 xT +0.293 xWSOC 24 268.603 0.959 <0.001 重度 Rs =0.960 xT 24 261.820 0.919 <0.001 Rs =0.681 xT +0.343 xPOXC 24 267.618 0.959 <0.001 总计 Rs =0.935 xT 72 489.252 0.875 <0.001 Rs =0.630 xT +0.383 xPOXC 72 447.716 0.928 <0.001 Rs =0.580 xT +0.278 xPOXC +0.167 xWSOC 72 322.352 0.934 <0.001 说明:Rs为土壤呼吸速率;xT为土壤温度;xPOXC为土壤易氧化有机碳;xWSOC为土壤水溶性有机碳;xMBC为土壤微生物生物量碳。
下载: 导出CSV
-
[1] LAL R. Soil carbon sequestration impacts on global climate change and food security [J]. Science, 2004, 304(5677): 1623 − 1627. [2] BOND-LAMBERTY B, BAILEY V, CHEN Min, et al. Globally rising soil heterotrophic respiration over recent decades [J]. Nature, 2018, 560(7716): 80 − 83. [3] CAO Mingkui, WOODWARD F I. Net primary and ecosystem production and carbon stocks of terrestrial ecosystems and their responses to climate change [J]. Global Change Biology, 1998, 4(2): 185 − 198. [4] 孟春, 王立海, 沈微. 择伐对小兴安岭针阔叶混交林土壤呼吸的影响[J]. 应用生态学报, 2008, 19(4): 729 − 734. MENG Chun, WANG Lihai, SHEN Wei. Effects of selective cutting on soil respiration in conifer/broad-leaved mixed forests in Xiao-xing’anling [J]. Chinese Journal of Applied Ecology, 2008, 19(4): 729 − 734. [5] XU Ming, SHANG Hua. Contribution of soil respiration to the global carbon equation [J]. Journal of Plant Physiology, 2016, 203: 16 − 28. [6] DANG Peng, GAO Yang, LIU Jinjing, et al. Effects of thinning intensity on understory vegetation and soil microbial communities of a mature Chinese pine plantation in the Loess Plateau [J]. Science of the Total Environment, 2018, 630: 171 − 180. [7] 张骏, 葛滢, 江波, 等. 浙江省杉木生态公益林碳储量效益分析[J]. 林业科学, 2010, 46(6): 22 − 26. ZHANG Jun, GE Ying, JIANG Bo, et al. Carbon storage efficiency of Cunninghamia lanceolata ecological service forest in Zhejiang [J]. Scientia Silvae Sinicae, 2010, 46(6): 22 − 26. [8] 李斌, 方晰, 项文化, 等. 湖南省杉木林植被碳贮量、碳密度及碳吸存潜力[J]. 林业科学, 2013, 49(3): 25 − 32. LI Bin, FANG Xi, XIANG Wenhua, et al. Carbon storage, carbon density and carbon sequestration potential of Cunninghamia lanceolata plantation in Hunan Province [J]. Scientia Silvae Sinicae, 2013, 49(3): 25 − 32. [9] ZHANG Hui, ZHOU Guomo, WANG Yixiang, et al. Thinning and species mixing in Chinese fir monocultures improve carbon sequestration in subtropical China [J]. European Journal of Forest Research, 2019, 138(3): 433 − 443. [10] 王兵, 魏文俊, 李少宁, 等. 中国杉木林生态系统碳储量研究[J]. 中山大学学报(自然科学版), 2008, 47(2): 93 − 98. WANG Bin, WEI Wenjun, LI Shaoning, et al. Carbon storage of Chinese fir forest ecosystem in China [J]. Acta Scientiarum Naturalium Universitatis Sunyatseni, 2008, 47(2): 93 − 98. [11] 邸富宏. 中国南方杉木人工林碳动态模拟研究[J]. 西北农林科技大学学报(自然科学版), 2016, 44(8): 127 − 134, 142. DI Fuhong. Simulation of carbon dynamics of Chinese fir plantation in southern China [J]. Journal of Northwest A&F University (Natural Science Edition), 2016, 44(8): 127 − 134, 142. [12] 田甜, 白彦锋, 张旭东, 等. 杉木人工林地力衰退的原因及对策研究[J]. 林业科技通讯, 2019, 23(4): 6 − 10. TIAN Tian, BAI Yanfeng, ZHANG Xudong, et al. The reason and countermeasure of the soil degradation of Chinese fir plantation [J]. Forest Science and Technology, 2019, 23(4): 6 − 10. [13] TIAN Dalun, YAN Wende, FANG Xi, et al. Influence of thinning on soil CO2 efflux in Chinese fir plantations [J]. Pedosphere, 2009, 19(3): 273 − 280. [14] LI Renshan, ZHENG Wenhui, YANG Qingpeng, et al. The response of soil respiration to thinning was not affected by understory removal in a Chinese fir (Cunninghamia lanceolata) plantation [J]. Geoderma, 2019, 353: 47 − 54. [15] WANG Dong, CHEN Xinli, CHEN H Y H, et al. Contrasting effects of thinning on soil CO2 emission and above-and belowground carbon regime under a subtropical Chinese fir plantation [J]. Science of the Total Environment, 2019, 690: 361 − 369. [16] 丁驰, 雷梅, 甘子莹, 等. 间伐和施肥对杉木人工林土壤温室气体排放的影响[J]. 生态学杂志, 2022, 41(6): 1056 − 1065. DING Chi, LEI Mei, GAN Ziying, et al. Effects of thinning and fertilization on GHGs emissions in Chinese fir plantation soil [J]. Chinese Journal of Ecology, 2022, 41(6): 1056 − 1065. [17] WU Jiasen, JIANG Peikun, CHANG S X, et al. Dissolved soil organic carbon and nitrogen were affected by conversion of native forests to plantations in subtropical China [J]. Canadian Journal of Soil Science, 2010, 90(1): 27 − 36. [18] WEIL R R, ISLAM K R, STINE M A, et al. Estimating active carbon for soil quality assessment: a simplified method for laboratory and field use [J]. American Journal of Alternative Agriculture, 2003, 18(1): 3 − 17. [19] XIONG Qinli, PAN Kaiwen, ZHANG Lin, et al. Warming and nitrogen deposition are interactive in shaping surface soil microbial communities near the alpine timberline zone on the eastern Qinghai-Tibet Plateau, southwestern China [J]. Applied Soil Ecology, 2016, 101: 72 − 83. [20] 鲍士旦. 土壤农化分析[M]. 北京: 中国农业出版社, 2000. BAO Shidan. Soil Agro-chemistrical Analysis [M]. Beijing: China Agricultural Press, 2000. [21] 朱文见, 张慧, 王懿祥. 采伐对森林土壤呼吸影响的研究进展[J]. 浙江农林大学学报, 2021, 38(5): 1000 − 1011. ZHU Wenjian, ZHANG Hui, WANG Yixiang. Research progress on effects of cutting on forest soil respiration [J]. Journal of Zhejiang A&F University, 2021, 38(5): 1000 − 1011. [22] 雷蕾, 肖文发, 曾立雄, 等. 马尾松林土壤呼吸组分对不同营林措施的响应[J]. 生态学报, 2016, 36(17): 5360 − 5370. LEI Lei, XIAO Wenfa, ZENG Lixiong, et al. Responses of soil respiration and its components to forest management in Pinus massioniana stand [J]. Acta Ecologica Sinica, 2016, 36(17): 5360 − 5370. [23] DOUKALIANOU F, RADOGLOU K, AGNELLI A E, et al. Annual greenhouse-gas emissions from forest soil of a peri-urban conifer forest in Greece under different thinning intensities and their climate-change mitigation potential [J]. Forest Science, 2019, 65(4): 387 − 400. [24] TERAMOTO M, LIANG Naishen, TAKAHASHI Y, et al. Enhanced understory carbon flux components and robustness of net CO2 exchange after thinning in a larch forest in central Japan [J]. Agricultural and Forest Meteorology, 2019, 274: 106 − 117. [25] FANG Shenzuo, LIN Da, TIAN Ye, et al. Thinning intensity affects soil-atmosphere fluxes of greenhouse gases and soil nitrogen mineralization in a lowland poplar plantation [J/OL]. Forests, 2016, 7(7): 141[2022-11-01]. doi: 10.3390/f7070141. [26] ZHAO Bo, CAO Jing, GENG Yan, et al. Inconsistent responses of soil respiration and its components to thinning intensity in a Pinus tabuliformis plantation in northern China [J]. Agricultural and Forest Meteorology, 2019, 265: 370 − 380. [27] EPRON D, NGAO J, GRANIER A. Interannual variation of soil respiration in a beech forest ecosystem over a six-year study [J]. Annals of Forest Science, 2004, 61(6): 499 − 505. [28] PANG Xueyong, HU Bin, BAO Weikai, et al. Effect of thinning-induced gap size on soil CO2 efflux in a reforested spruce forest in the eastern Tibetan Plateau [J]. Agriculture of Forest Meteorology, 2016, 220: 1 − 9. [29] 张庆晓, 陈珺, 朱向涛, 等. 杉木林土壤温室气体排放对毛竹入侵及采伐的短期响应[J]. 浙江农林大学学报, 2021, 38(4): 703 − 711. ZHANG Qingxiao, CHEN Jun, ZHU Xiangtao, et al. On the short-term response of soil greenhouse gas emissions in Cunninghamia lanceolata forest to the expansion and eradication of Phyllostachys edulis [J]. Journal of Zhejiang A&F University, 2021, 38(4): 703 − 711. [30] LUAN Junwei, LIU Shirong, ZHU Xueling, et al. Roles of biotic and abiotic vatiables in determing spatial variation of soil respiration in sencondary oak and planted pine forests [J]. Soil Biology and Biochemistry, 2012, 44(1): 143 − 150. [31] 左强, 何怀江, 张春雨, 等. 采伐对蛟河阔叶红松混交林土壤呼吸的影响[J]. 北京林业大学学报, 2016, 38(4): 71 − 76. ZUO Qiang, HE Huaijiang, ZHANG Chunyu, et al. Effect of cutting on soil respiration in a mixed broadleaf-Korean pine forest in western foothill of Changbai Mountain, northeast China [J]. Journal of Beijing Forestry University, 2016, 38(4): 71 − 76. [32] SIMONIN K, KOLB T, MONTES-HELU M, et al. The influence of thinning on components of stand water balance in a ponderosa pine forest stand during and after extreme drought [J]. Agriculture and Forest Meteorology, 2007, 143(3/4): 266 − 276. [33] 褚金翔, 张小全. 川西亚高山林区三种土地利用方式下土壤呼吸动态及组分区分[J]. 生态学报, 2006, 26(6): 1693 − 1700. ZHU Jinxiang, ZHANG Xiaoquan. Dynamic and fractionalization of soil respiration under three different land use/covers in the subalpine region of western Sichuan Province, China [J]. Acta Ecologica Sinica, 2006, 26(6): 1693 − 1700. [34] 张睿, 白杨, 刘娟, 等. 亚热带天然阔叶林转换为杉木人工林对土壤呼吸的影响[J]. 应用生态学报, 2015, 26(10): 2946 − 2952. ZHANG Rui, BAI Yang, LIU Juan, et al. Effects of conversion of natural broad-leaved forest to Chinese fir plantation on soil respiration in subtropical China [J]. Chinese Journal of Applied Ecology, 2015, 26(10): 2946 − 2952. [35] 赵吉霞, 王邵军, 陈奇伯, 等. 滇中云南松林土壤呼吸季节动态及其影响因子研究[J]. 西北林学院学报, 2015, 30(3): 8 − 13, 20. ZHAO Jixia, WANG Shaojun, CHEN Qibo, et al. Soil respiration and its affecting factors of Pinus yunnanensis in the middle regions of Yunnan [J]. Journal of Northwest Forestry University, 2015, 30(3): 8 − 13, 20. [36] IQBAL J, HU Ronggui, DU Lijun, et al. Differences in soil CO2 flux between different land use types in mid-subtropical China [J]. Soil Biology and Biochemistry, 2008, 40(9): 2324 − 2333. [37] 王晓荣, 雷蕾, 曾立雄, 等. 抚育间伐对马尾松林土壤活性有机碳的短期影响[J]. 生态学杂志, 2021, 40(4): 1049 − 1061. WANG Xiaorong, LEI Lei, ZENG Lixiong, et al. Short-term effects of tending thinning on soil labile organic carbon in Pinus massoniana stands [J]. Chinese Journal of Ecology, 2021, 40(4): 1049 − 1061. [38] WANG Huimei, LIU Wei, WANG Wenjie, et al. Influence of long-term thinning on the biomass carbon and soil respiration in a larch (Larix gmelinii) forest in northeastern China [J/OL]. The Scientific World Journal, 2013, 2013: 865645[2022-11-01]. doi: 10.1155/2013/865645. [39] MICHALZIK B, KALBITZ K, PARK J H, et al. Fluxes and concentrations of dissolved organic carbon and nitrogen: a synthesis for temperate forests [J]. Biogeochemistry, 2001, 52(2): 173 − 205. [40] YANG Yusheng, CHEN Guangshui, GUO Jianfen, et al. Soil respiration and carbon balance in a subtropical native forest and two managed plantations [J]. Plant Ecology, 2007, 193: 71 − 84. [41] 刘可, 韩海荣, 康峰峰, 等. 山西太岳山油松人工林生长季土壤呼吸对择伐强度的响应[J]. 生态学杂志, 2013, 32(12): 3173 − 3181. LIU Ke, HAN Hairong, KANG Fengfeng, et al. Responses of soil respiration to selective cutting intensity in Pinus tabulaeformis plantation in Taiyue Mountain of China during growing season [J]. Chinese Journal of Ecology, 2013, 32(12): 3173 − 3181. [42] 巫志龙, 周成军, 周新年, 等. 不同强度采伐5年后杉阔混交人工林土壤呼吸速率差异[J]. 林业科学, 2019, 55(6): 142 − 149. WU Zhilong, ZHOU Chengjun, ZHOU Xinnian, et al. Difference in soil respiration rates of the mixed plantations of Cunninghamia lanceolata and broadleaved tree 5 year after harvesting at different intensities [J]. Scientia Silvae Sinicae, 2019, 55(6): 142 − 149. [43] TU Lihua, HU Tingxing, ZHANG Jian, et al. Nitrogen addition stimulates different components of soil respiration in a subtropical bamboo ecosystem [J]. Soil Biology and Biochemistry, 2013, 58: 255 − 264. [44] SULLIVAN B W, KOLB T E, HART S C, et al. Thinning reduces soil carbon dioxide but not methane flux from southwestern USA Ponderosa pine forests [J]. Forest Ecology and Management, 2008, 255(12): 4047 − 4055. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20220704
 点击查看大图
点击查看大图
计量
- 文章访问数: 1471
- HTML全文浏览量: 317
- PDF下载量: 26
- 被引次数: 0
