




-
薄壳山核桃Carya illinoinensis[1]又称美国山核桃、长山核桃,是胡桃科Juglandaceae山核桃属Carya高大落叶乔木,原产于北美洲东南部[2]。薄壳山核桃果仁含有丰富的油脂,含油率为70%~75%[3],油脂成分中的不饱和脂肪酸占总脂肪酸含量的90%以上,是不饱和脂肪酸占比最高的坚果之一[4]。薄壳山核桃还富含抗氧化成分,如酚类物质、单宁、黄酮类、维生素E和植物甾醇等[5-6],这些物质对预防心脑血管疾病有一定的作用[7]。薄壳山核桃树干通直,质地坚韧,抗击力强,是一种优良木材;其树形挺拔、抗逆性强,可作为园林绿化的观赏树种。薄壳山核桃适应性强,目前已被世界多国引种栽培[8]。中国在19世纪初就已开始陆续引进薄壳山核桃,目前已在20多个省(区、市)栽培[9]。近些年安徽、浙江、江苏等地的栽培已开始规模化和产业化。薄壳山核桃的产量也开始上升,经济效益增加。MaxEnt模型(最大熵模型)可以根据已知的物种分布点和气候变量,对物种潜在的分布区进行预测。该模型具有运行时间短,操作简单,精确度高等优点,尤其在物种分布点较少的情况下可对其展开精准预测[10-11],因此得到广泛应用[12-13]。MaxEnt模型被用于大量物种的潜在分布区预测,如经济植物潜在种植区的预测[14-16]、珍稀物种潜在生境的预测[17-19]、入侵物种的防控[20-21]和病虫害的防治[22-24]等方面。张日清等[25]根据薄壳山核桃在北美洲的生态宽度和前期中国引种效果,划分了薄壳山核桃的适生区范围;程晋昕等[26]根据北美产地气候数据与MaxEnt模型,对薄壳山核桃在云南的适生区进行了划分。鉴于薄壳山核桃寿命长的特点,预测当前和未来在中国的潜在种植区,有利于薄壳山核桃在中国的引种栽培。本研究以薄壳山核桃为研究对象,基于北美栽培区的分布点和全球的环境气候数据,利用MaxEnt模型预测薄壳山核桃当前和未来3种气候情景下在中国的潜在适生区,为薄壳山核桃长期的引种栽培提供理论参考。
-
从薄壳山核桃在北美洲的分布数据中,共获取274条记录点,作为MaxEnt软件预测位点,数据信息来源于全球生物多样性信息机构(GBIF,
)。为避免过拟合,以15 000 m为半径,设置圆形缓冲区,每个缓冲区内保留1个样本点[27]。最后共得到152个薄壳山核桃在北美洲的坐标数据,将其存为*.CSV格式以备用。 -
19种生物气候变量数据(表1)来源于世界气候数据网站
(http://www.worldclim.org/ ),空间分辨率为2.5 min,这些气候变量(bio)与生物的生存相关,由月均气温和降水量产生,反映了地区的气温、降水等特点[28]。数据包括现代气候数据(1970—2000年)、未来气候数据(2041—2060年和2061—2080年)。本研究选取第6次国际耦合模式比较计划(CMIP6)中的未来气候数据,模式为BCC-CSM2-MR。研究未来气候数据中选择了3种不同气候情景:SSP2-4.5(ssp245)、SSP3-7.0(ssp370)和SSP5-8.5(ssp585),分别代表了中等强迫情景(2100年辐射强迫稳定在约4.5 W·m−2)、中等至高强迫情景(2100年辐射强迫稳定在约7.0 W·m−2)和高强迫情景(2100年辐射强迫稳定在约8.5 W·m−2),辐射强迫数值越大表示碳排放程度越强,未来环境温度越高[29]。表 1 气候变量
Table 1. Climate variables
气候变量 描述 单位 气候变量 描述 单位 bio1 年均气温 ℃ bio11 最冷季平均气温 ℃ bio2 昼夜温差月均值 ℃ bio12 年降水量 mm bio3 等温性 bio13 最湿月降水量 mm bio4 气温的季节性 bio14 最干月降水量 mm bio5 最暖月最高气温 ℃ bio15 季节性降水 bio6 最冷月最低气温 ℃ bio16 最湿季降水量 mm bio7 年均气温范围 ℃ bio17 最干季降水量 mm bio8 最湿季平均气温 ℃ bio18 最暖季降水量 mm bio9 最干季平均气温 ℃ bio19 最冷季降水量 mm bio10 最暖季平均气温 ℃ 说明:等温性为昼夜温差月均值除以年均气温范围;年均气温范围为最暖月最高气温与最冷月最低气温的差值 -
气候变量与薄壳山核桃分布点的过强相关性,会对预测结果产生过拟合作用。为避免这一现象,提取19个生物气候变量与152个薄壳山核桃分布位点的数据,用SPSS 20.0进行Pearson相关性分析。若2个气候变量之间的相关性绝对值|r|>0.8,则剔除在预测中贡献率较低的气候变量[30]。最后得到8个气候变量:年均气温(bio1)、昼夜温差月均值(bio2)、气温的季节性(bio4)、最干季平均气温(bio9)、最暖季平均气温(bio10)、年降水量(bio12)、最干月降水量(bio14)和最暖季降水量(bio18),用于最终的预测。
-
将最后筛选的8个气候变量和样点分布数据加载到MaxEnt 3.4.1软件中,测试数据集设为25%,训练数据集为75%,输出分布值的形式选择Logistic,其他参数默认。
MaxEnt模型预测的结果为受试者工作特征曲线(ROC),ROC曲线下的面积(AUC)取值为(0, 1]。AUC越接近1,模型预测的结果越准确,当AUC>0.9时,表示预测结果很精确[31]。
-
根据MaxEnt模型预测的结果,利用ArcGIS 10.4.1软件掩膜提取功能提取薄壳山核桃在中国的适生范围。再利用ArcGIS的重分类功能,将MaxEnt模型预测的薄壳山核桃在中国实际栽培区域的适生概率进行适生等级划分。根据适生概率(P),将预测的薄壳山核桃适生区划分为3类:非适生区(P<0.08)、潜在低适生区(0.08<P<0.25)、潜在高适生区(P>0.25)。
-
从预测精度ROC曲线(图1)可知:训练集的AUC为0.987,检验集AUC为0.985。AUC均大于0.9,表示预测结果非常好,模型对薄壳山核桃适生区预测的准确度很高。
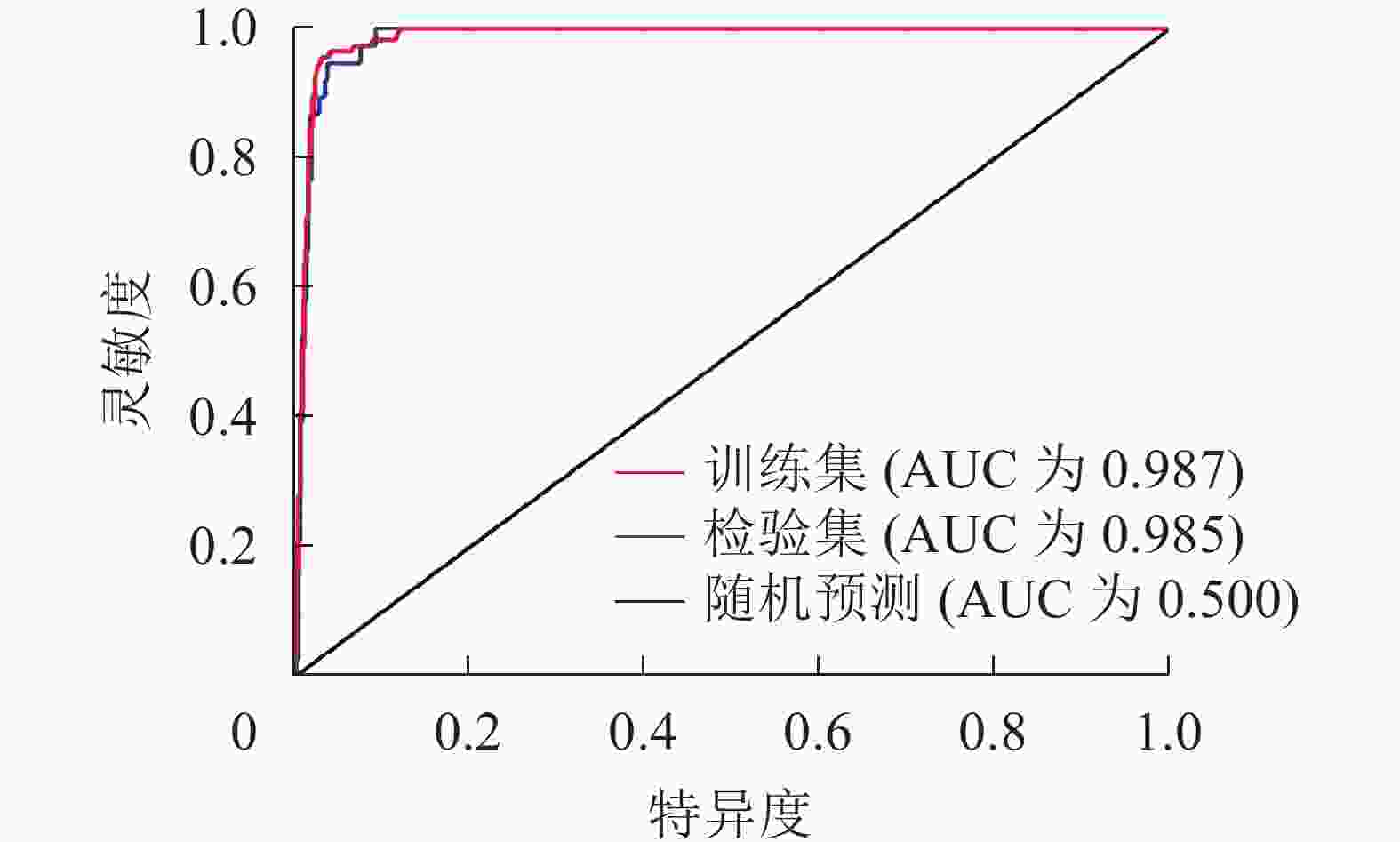
图 1 MaxEnt预测结果的AUC值
Figure 1. Forecast results of the AUC value by MaxEnt model
-
从表2可知:最终预测的8个气候变量中,贡献率前4的气候变量为最干月降水量(bio14)、昼夜温差月均值(bio2)、最暖季平均气温(bio10)和气温的季节性(bio4),累积贡献率达83.2%。最干月降水量的贡献率最大,达33.8%,表明最干月降水量是影响薄壳山核桃分布的重要气候变量。其次是昼夜温差月均值,占了21.8%。最暖季平均气温和气温的季节性贡献率都超过了10%,分别为17.3%和10.4%。剩余的4个气候变量贡献率总和为16.8%。
表 2 气候变量的贡献率
Table 2. Contribution of climate variables
气候变量 贡献率/% 累积贡献/% 气候变量 贡献率/% 累积贡献/% bio14 33.8 33.8 bio18 7.1 90.3 bio2 21.8 55.6 bio1 5.5 95.8 bio10 17.3 72.9 bio12 3.8 99.6 bio4 10.3 83.2 bio9 0.4 100.0 -
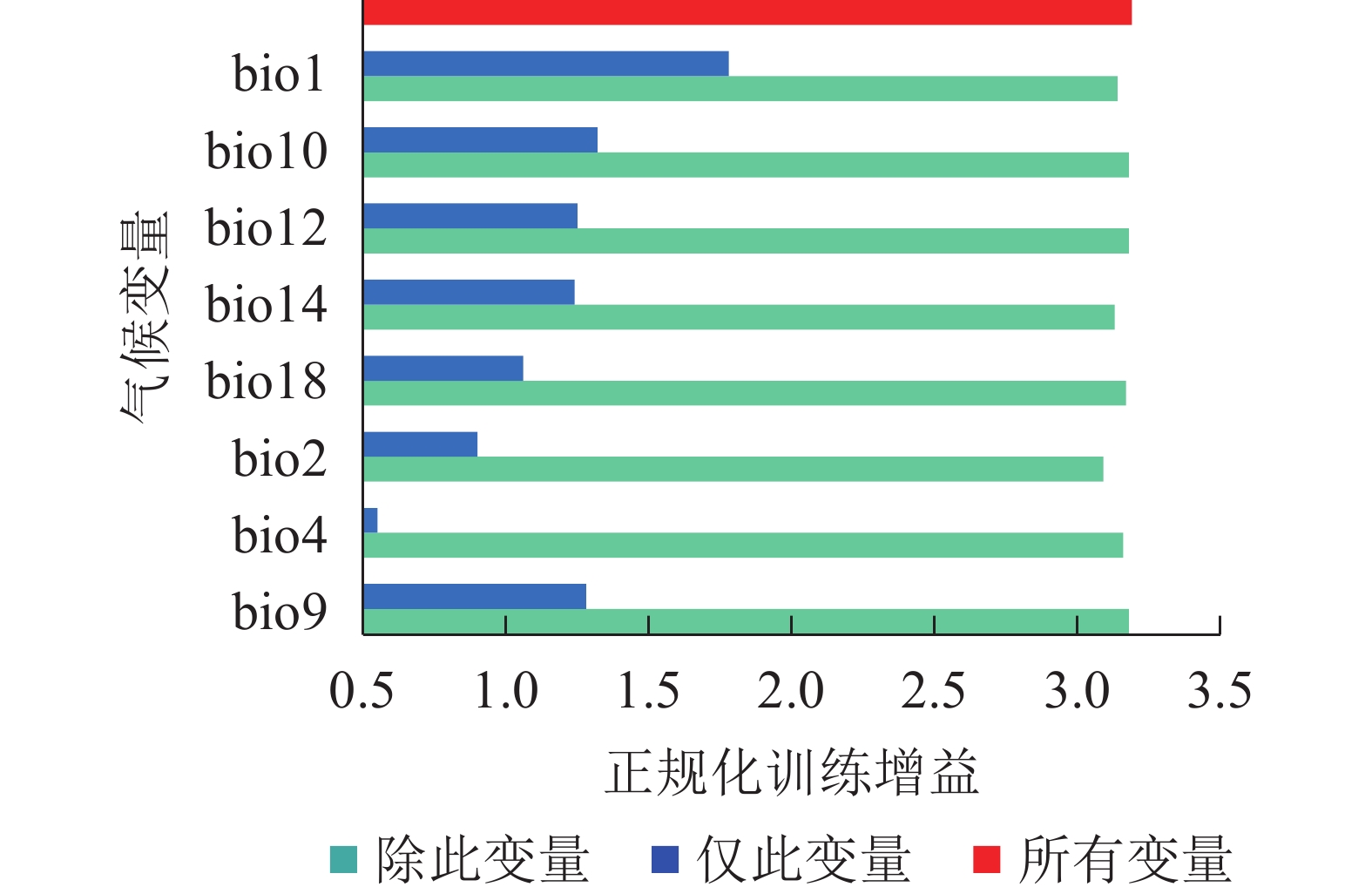
从图2可见:在刀切法检验中,变量的得分越高,对正规化训练增益影响越大。年均气温(bio1)的训练得分最高,超过1.7,说明其对薄壳山核桃的潜在分布影响最为重要。最暖季平均气温(bio10)、年降水量(bio12)、最干月降水量(bio14)和最干季平均气温(bio9)的训练得分超过1.2,说明它们对薄壳山核桃的分布较为重要。综上,年均气温(bio1)、最暖季平均气温(bio10)、年降水量(bio12)、最干月降水量(bio14)和最干季平均气温(bio9)是影响薄壳山核桃分布的关键气候变量。
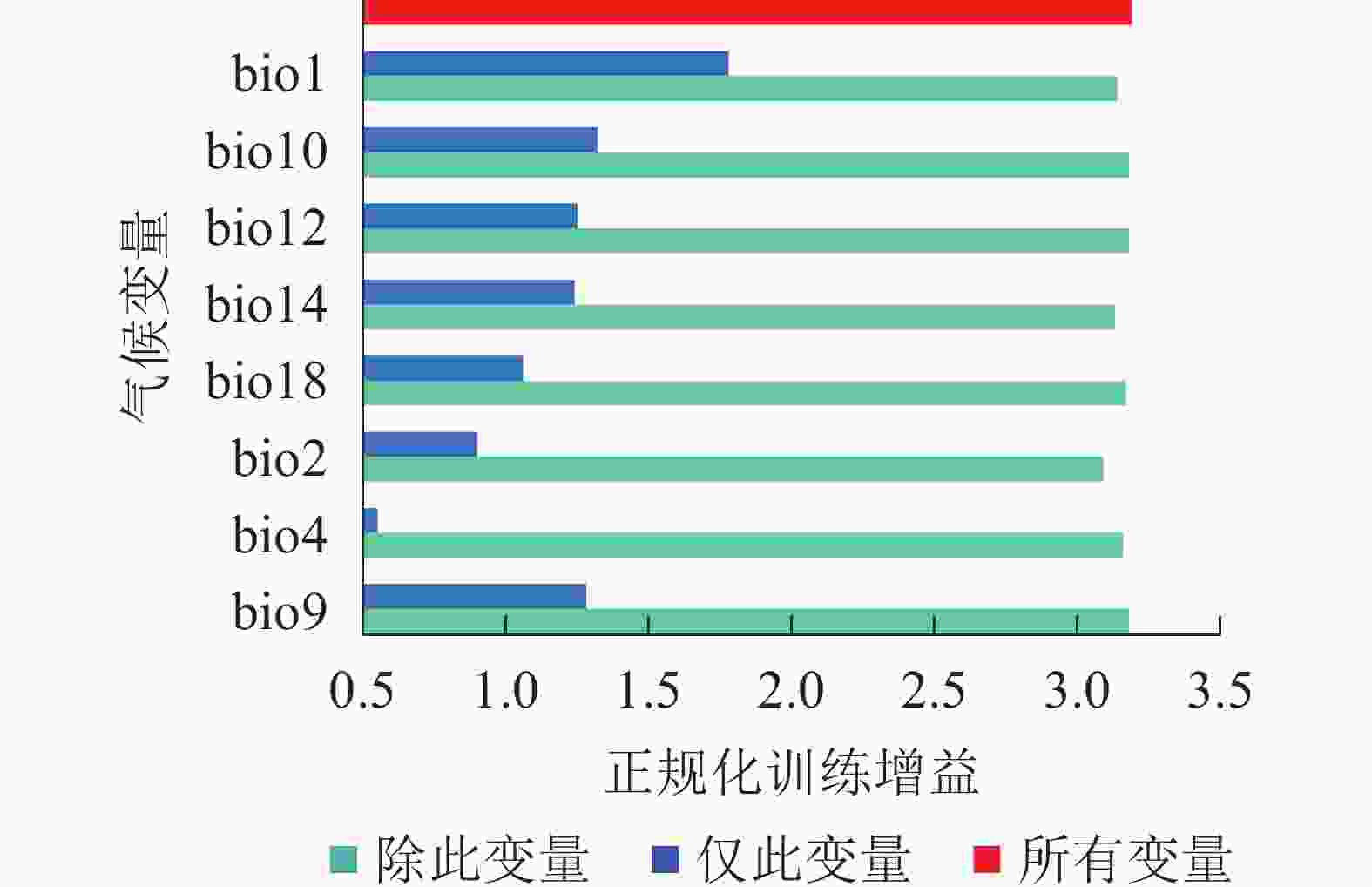
图 2 刀切法检验气候变量的测试增益结果
Figure 2. Jackknife test to examine training gain result of climate variables
-
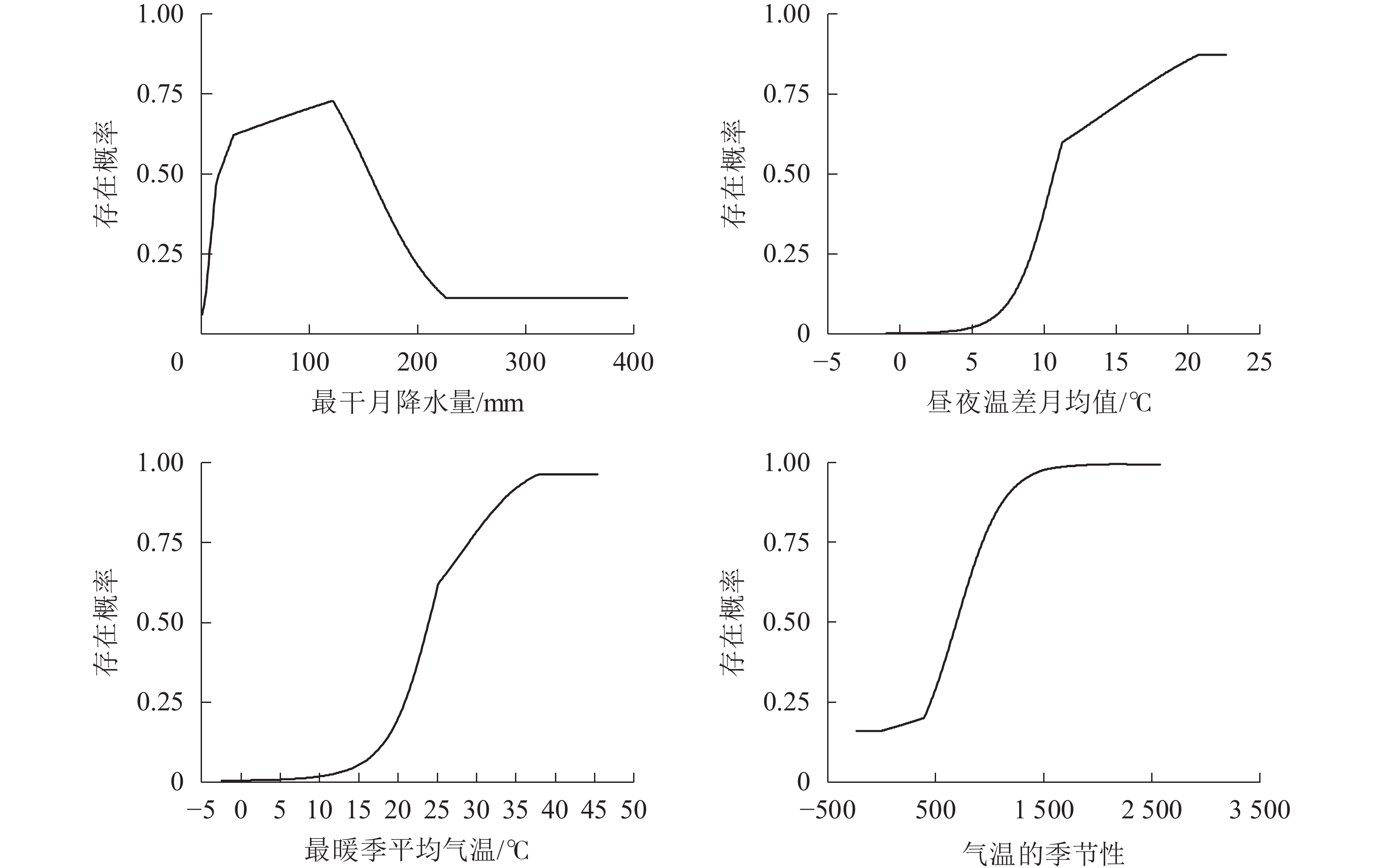
选取对模型预测贡献率排名前4的气候变量对薄壳山核桃的适宜性气候进行分析。一般认为,当存在概率大于0.5时[32],其对应的气候变量数值更利于薄壳山核桃的生长。最干月降水量(bio14)为15.5~156.0 mm时,较适宜薄壳山核桃生长(图3)。当最干月降水量低于15.5 mm时,存在概率急剧下降;当最干月降水量大于220.0 mm时,存在概率处于较低水平,说明最干月降水量过低过高都不利于薄壳山核桃生长。当昼夜温差月均值(bio2)大于10.5 ℃,最暖季平均气温(bio10)大于23.8 ℃,气温的季节性(bio4)大于687时,有利于薄壳山核桃的生长。
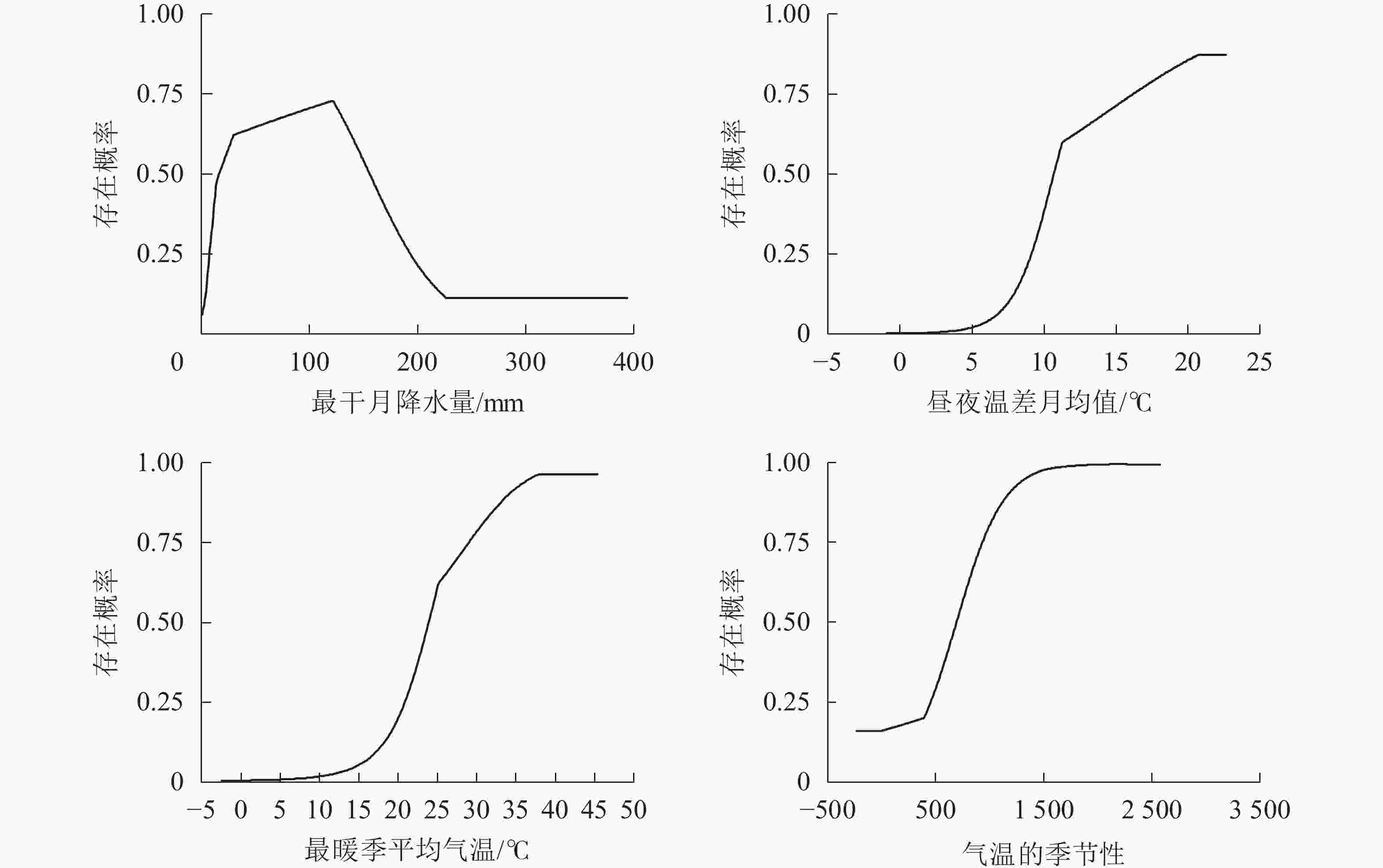
图 3 薄壳山核桃存在概率对主要气候变量的响应曲线
Figure 3. Response curves of existence probability of C. illinoinensis to main climate variables
-
当前气候环境下,薄壳山核桃适生分布区的总面积为891 936 km2,其中潜在高适生区面积为55 142 km2,潜在低适生区面积为836 794 km2。大部分适生区位于中国东部地区和中部地区,主要集中在湖南、江西、安徽、浙江的北部和西部、福建西北部、湖北东部、河南南部和江苏中西部地区。在广东北部、广西东北部、山东南部和云南中西部有少量潜在低适生区。潜在高适生区在江西、河南西部、湖南东部和安徽北部有少量存在。
-
不同年代不同碳排放的情景下,薄壳山核桃在中国的潜在高适生区面积相对于当前适生区有较大幅度的增加,潜在低适生区面积增加不多,甚至在有些情景下有所下降(表3)。
表 3 不同气候情景下薄壳山核桃在中国的适生区面积
Table 3. Suitable area of C. illinoinensis in China under different climate change scenarios
气候情景 起止年份 潜在高适生区面积/
km2潜在低适生区面积/
km2适生区总面积/km2
(约占中国陆地面积的比例/%)当前 1970—2000 55 142 836 794 891 936(9.29) 中等强迫情景(ssp245) 2041—2060 265 155 906 770 1 171 925(12.21) 2061—2080 591 863 827 055 1 418 918(14.78) 中等至高强迫情景(ssp370) 2041—2060 581 777 798 618 1 380 395(14.38) 2061—2080 837 574 866 531 1 704 105(17.75) 高强迫情景(ssp585) 2041—2060 571 657 842 062 1 413 719(14.73) 2061—2080 846 533 852 165 1 698 698(17.69) 未来不同碳排放情景下,薄壳山核桃的适生区地理位置变化不大,主要还是集中在中国的中东部地区。随着碳排放程度增加,适生区有向北方扩展的趋势,部分已延伸至河北南部、辽宁东部和吉林南部。潜在高适生区在福建、江西、浙江、湖南、湖北、安徽、河南、江苏和山东等省份内部扩张。河北、陕西和山西有少量潜在低适生区增加。云南的潜在低适生区主要还是集中在中西部。
-
本研究预测结果表明:当前环境下薄壳山核桃的适生区主要集中在中国的东部和中部地区,这与现实情况基本相符。但预测分布区未涵盖四川、重庆等地,可能是由于预测所用的环境因素过少,对预测产生了一定影响。四川等地也可以栽种薄壳山核桃,其适应性也较好[33]。云南的适生区相对较少,主要集中在中西部地区,这也与其实际的种植区域相符[26]。广东和广西的北部只有少量适生区,而海南没有适生区分布,这是因为薄壳山核桃需要一定的低温促进花芽分化,温度过高也会使其死亡[25]。北京、辽宁等地也栽种过薄壳山核桃,由于积温不足等原因导致其不结果实,但可以作为绿化树栽植。山东南部有结果大树,北部结果不良或品质不佳[25]。这与预测的薄壳山核桃适生区南北界限也较为符合。中国中东部地区适生区的预测也是较为准确的,近几年江西、安徽等地种植的薄壳山核桃产量开始逐年上升。
-
结合刀切法和模型预测中的贡献率可挖掘限制薄壳山核桃生长的气候变量。本研究中,最干月降水量(bio14)、昼夜温差月均值(bio2)、气温的季节性(bio4)、年均气温(bio1)、最暖季平均气温(bio10)、年降水量(bio12)和最干季平均气温(bio9)为主要气候变量,最佳阈值范围分别为15.5~156 mm、>10.5 ℃(在一定范围内)、>687(在一定范围内)、11.3~23 ℃、>23.8 ℃(在一定范围内)、>665 mm(在一定范围内)、−26.5~23.5 ℃。其中最干月降水量、昼夜温差月均值、最暖季平均气温和气温的季节性预测贡献率占80%以上。年均气温和年降水量的适宜范围与张日清等[9]的研究相符。本研究以气候变量为参考数据,未将土壤、地形等因素考虑在内,存在一定的局限性[34]。
-
2041—2060年,3种气候情景下的薄壳山核桃适生区均有大量的内扩和轻微的北扩趋势。相较于当前气候情景,2041—2060年,3种气候情景下,原来存在于中国中东部地区的大量潜在低适生区转变为潜在高适生区。其中,中等至高等强迫情景下和高等强迫情景下,潜在低适生区转变为潜在高适生区的面积较大;2061—2080年,3种气候情景下,大量潜在低适生区也转变为潜在高适生区,潜在高适生区由江西、湖南东部、湖北东部向北延伸至安徽、河南等省份。可见未来碳排放增强的情景下,薄壳山核桃的适生区有向北方扩展的趋势,适生区最北延伸到了吉林南部和辽宁东部。赤桉Eucalyptus camaldulensis、千金榆Carpinus cordata和白栎Quercus fabri等物种在未来碳排放增强的情景下,适生区都有向北扩展的趋势,这与气温上升有一定的关系[34-36]。中国中东部地区可以参考本研究结果的适生区范围,在适宜区域种植薄壳山核桃。
-
利用MaxEnt模型预测薄壳山核桃在中国的适生区有较高精度。在当前情景下,薄壳山核桃的适生区集中在中国中东部地区;随着未来气温的升高,薄壳山核桃的潜在高适生区在中国相应的中东部地区内部扩张,潜在适生区存在向北扩张的趋势,中国中东部地区比较适宜栽种薄壳山核桃。
Prediction on the potential planting area of Carya illinoinensis in China based on MaxEnt model
-
摘要:
目的 预测当前和未来中国适宜种植薄壳山核桃Carya illinoinensis的地区,对薄壳山核桃的引种栽培提供参考。 方法 基于北美薄壳山核桃栽培区的分布数据和世界环境气候数据,利用MaxEnt模型和ArcGIS预测薄壳山核桃当前以及未来不同气候情景下在中国的适生区范围。 结果 ①训练集和检验集的AUC值分别为0.987和0.985,表明预测结果精度高。②刀切法检验结果和贡献率结果表明:年均气温、最暖季平均气温、年降水量、最干月降水量、最干季平均气温、昼夜温差月均值和气温的季节性是薄壳山核桃生长的关键气候变量。③预测薄壳山核桃当前在中国的适生区主要集中在中东部省份,包括江西、湖南、湖北、安徽、浙江、江苏和河南;在未来碳排放增强的气候情景下,薄壳山核桃在中国中东部省份的潜在高适生区的面积大幅增加,适生范围有向北移动的趋势,少量延伸到了辽宁东部和吉林南部。 结论 利用MaxEnt模型预测的薄壳山核桃适生区主要集中在中国的中东部省份;随着未来气温的升高,薄壳山核桃的潜在高适生区在中国相应的中东部地区内扩张。图3表3参36 Abstract:Objective The suitable areas for planting Carya illinoinensis in China at present and in the future are predicted, which can provide reference for the cultivation of C. illinoinensis. Method With the employment of MaxEnt maximum entropy model and ArcGIS, this study is aimed to make predictions of the suitable areas for planting C. illinoinensis in China at present and in the future in different climate scenarios based on distribution data of C. illinoinensis cultivated areas in North America and the global environmental climate variables so as to provide reference for the introduction and cultivation of C. illinoinensis. Result (1) The area under the curve (AUC) of the training data and the testing data were 0.987 and 0.985, indicating high accuracy of the prediction results. (2) The jackknife test results and contribution rate results showed that the annual mean temperature, the mean temperature of the warmest season, the annual precipitation, the precipitation of the driest month, the mean temperature of the driest season, the mean diurnal range and the temperature seasonality were the key climatic variables for the growth of C. illinoinensis. (3) It is predicted that the suitable planting areas of C. illinoinensis in China at present are mainly distributed in Jiangxi, Hunan, Hubei, Anhui, Zhejiang, Jiangsu and Henan whereas under a warmer climate scenario in the future, the highly suitable planting areas of C. illinoinensis in China will increase significantly, and show a tendency of moving northward to cover a some limited areas in eastern Liaoning and Southern Jilin. Conclusion The predictions made of the suitable planting area of C. illinoinensis in China using MaxEnt model were featured with high accuracy: currently, the suitable planting areas of C. illinoinensis are mainly distributed in central and eastern China while in the future, the potential highly suitable planting areas for C. illinoinensis in central and eastern China will expand inward with the increase of temperature and the potential suitable planting areas tend to expand northward. [Ch, 3 fig. 3 tab. 36 ref.] -
Key words:
- Carya illinoinensis /
- MaxEnt model /
- jackknife test /
- potential planting area /
- climate variables
-
图 2 刀切法检验气候变量的测试增益结果
Figure 2 Jackknife test to examine training gain result of climate variables
图 3 薄壳山核桃存在概率对主要气候变量的响应曲线
Figure 3 Response curves of existence probability of C. illinoinensis to main climate variables
表 1 气候变量
Table 1. Climate variables
气候变量 描述 单位 气候变量 描述 单位 bio1 年均气温 ℃ bio11 最冷季平均气温 ℃ bio2 昼夜温差月均值 ℃ bio12 年降水量 mm bio3 等温性 bio13 最湿月降水量 mm bio4 气温的季节性 bio14 最干月降水量 mm bio5 最暖月最高气温 ℃ bio15 季节性降水 bio6 最冷月最低气温 ℃ bio16 最湿季降水量 mm bio7 年均气温范围 ℃ bio17 最干季降水量 mm bio8 最湿季平均气温 ℃ bio18 最暖季降水量 mm bio9 最干季平均气温 ℃ bio19 最冷季降水量 mm bio10 最暖季平均气温 ℃ 说明:等温性为昼夜温差月均值除以年均气温范围;年均气温范围为最暖月最高气温与最冷月最低气温的差值  下载: 导出CSV
下载: 导出CSV
表 2 气候变量的贡献率
Table 2. Contribution of climate variables
气候变量 贡献率/% 累积贡献/% 气候变量 贡献率/% 累积贡献/% bio14 33.8 33.8 bio18 7.1 90.3 bio2 21.8 55.6 bio1 5.5 95.8 bio10 17.3 72.9 bio12 3.8 99.6 bio4 10.3 83.2 bio9 0.4 100.0
下载: 导出CSV
表 3 不同气候情景下薄壳山核桃在中国的适生区面积
Table 3. Suitable area of C. illinoinensis in China under different climate change scenarios
气候情景 起止年份 潜在高适生区面积/
km2潜在低适生区面积/
km2适生区总面积/km2
(约占中国陆地面积的比例/%)当前 1970—2000 55 142 836 794 891 936(9.29) 中等强迫情景(ssp245) 2041—2060 265 155 906 770 1 171 925(12.21) 2061—2080 591 863 827 055 1 418 918(14.78) 中等至高强迫情景(ssp370) 2041—2060 581 777 798 618 1 380 395(14.38) 2061—2080 837 574 866 531 1 704 105(17.75) 高强迫情景(ssp585) 2041—2060 571 657 842 062 1 413 719(14.73) 2061—2080 846 533 852 165 1 698 698(17.69)
下载: 导出CSV
-
[1] 中国科学院中国植物志编辑委员会. 中国植物志[M]. 北京: 科学出版社, 2004. Editorial Committee of Flora of China, Chinese Academy of Sciences. Flora of China[M]. Beijing: Science Press, 2004. [2] HALL G D. Pecan food potential in prehistoric North America [J]. Econ Bot, 2000, 54(1): 103 − 112. [3] WAKELING L T, MASON R L, D’ARCY B R, et al. Composition of pecan cultivars Wichita and Western Schley [Carya illinoinensis (Wangenh.)K. Koch] grown in Australia [J]. J Agric Food Chem, 2001, 49(3): 1277 − 1281. [4] YANG Jun. Brazil nut and associated health benefits: a review [J]. LWT-Food Sci Technol, 2009, 42(10): 1573 − 1580. [5] BOLLING B W, CHEN C Y O, MCKAY D L, et al. Tree nut phytochemicals: composition, antioxidant capacity, bioactivity, impact factors. A systematic review of almonds, Brazils, cashews, hazelnuts, macadamias, pecans, pine nuts, pistachios and walnuts [J]. Nutr Res Rev, 2011, 24(2): 244 − 275. [6] 罗会婷, 贾晓东, 翟敏, 等. 薄壳山核桃营养成分的研究进展[J]. 中国农学通报, 2017, 33(8): 39 − 46. LUO Huiting, JIA Xiaodong, ZHAI Min, et al. A review of nutritional components in Carya illinoinensis [J]. Chin Agric Sci Bull, 2017, 33(8): 39 − 46. [7] MORGAN W A, CLAYSHULTE B J. Pecans lower low density lipoprotein cholesterol in people with normal lipid levels [J]. J Am Dietetic Association, 2000, 100(3): 312 − 318. [8] ZHANG Rui, PENG Fangren, LI Yongrong. Pecan production in China [J]. Sci Hortic, 2015, 197(10): 719 − 727. [9] 张日清, 吕芳德, 何方. 美国山核桃引种栽培区划研究(Ⅰ)原生境与新生境自然条件比较[J]. 中南林学院学报, 2001, 21(2): 1 − 5. ZHANG Riqing, LÜ Fangde, HE Fang. Cultural regionalization for the introduced pecan tree in China (Ⅰ) comparative study of the site conditions between USA and China [J]. J Cent South For Univ, 2001, 21(2): 1 − 5. [10] HERNANDEZ P A, GRAHAM C H, MASTER L L, et al. The effect of sample size and species characteristics on performance of different species distribution modeling methods [J]. Ecography, 2006, 29(5): 773 − 785. [11] 陈新美, 雷渊才, 张雄清, 等. 样本量对MaxEnt模型预测物种分布精度和稳定性的影响[J]. 林业科学, 2012, 48(1): 53 − 59. CHEN Xinmei, LEI Yuancai, ZHANG Xiongqing, et al. Effects of sample sizes on accuracy and stability of maximum entropy model in predicting species distribution [J]. Sci Silv Sin, 2012, 48(1): 53 − 59. [12] 胡秀, 吴福川, 郭微, 等. 基于MaxEnt生态学模型的檀香在中国的潜在种植区预测[J]. 林业科学, 2014, 50(5): 27 − 33. HU Xiu, WU Fuchuan, GUO Wei, et al. Identification of potential cultivation region for Santalum album in China by the MaxEnt ecologic niche model [J]. Sci Silv Sin, 2014, 50(5): 27 − 33. [13] 李宏群, 刘晓莉, 汪建华, 等. 未来气候变化对重庆榨菜种植适宜区的影响[J]. 应用生态学报, 2018, 29(8): 2651 − 2657. LI Hongqun, LIU Xiaoli, WANG Jianhua, et al. Influence of climate change on the suitable ranges for planting pickled mustard tuber in Chongqing [J]. Chin J Appl Ecol, 2018, 29(8): 2651 − 2657. [14] 张梅, 禄彩丽, 魏喜喜, 等. 基于MaxEnt模型新疆枣潜在适生区预测[J]. 经济林研究, 2020, 38(1): 152 − 161. ZHANG Mei, LU Caili, WEI Xixi, et al. Potential suitable area forecast of jujube in Xinjiang based on MaxEnt model [J]. Nonwood For Res, 2020, 38(1): 152 − 161. [15] 王书越, 潘少安, 王明睿, 等. 基于MaxEnt模型评估刺五加在东北地区的空间分布[J]. 生态学报, 2019, 39(9): 3277 − 3286. WANG Shuyue, PAN Shaoan, WANG Mingrui, et al. Assessing the geographic distribution of Acanthopanax senticosus in northeastern China based on the MaxEnt model [J]. Acta Ecol Sin, 2019, 39(9): 3277 − 3286. [16] 张童, 黄治昊, 彭杨靖, 等. 基于Maxent模型的软枣猕猴桃在中国潜在适生区预测[J]. 生态学报, 2020, 40(14): 4921 − 4928. ZHANG Tong, HUANG Zhihao, PENG Yangjing, et al. Prediction of potential suitable areas of Actinidia arguta in China based on Maxent model [J]. Acta Ecol Sin, 2020, 40(14): 4921 − 4928. [17] MATYUKHINA D S, MIQUELLE D G, MURZIN A A, et al. Assessing the influence of environmental parameters on Amur tiger distribution in the Russian far east using a MaxEnt modeling approach [J]. Achiev Life Sci, 2014, 8(2): 95 − 100. [18] 王雨生, 王召海, 邢汉发, 等. 基于MaxEnt模型的珙桐在中国潜在适生区预测[J]. 生态学杂志, 2019, 38(4): 1230 − 1237. WANG Yusheng, WANG Zhaohai, XING Hanfa, et al. Prediction of potential suitable distribution of Davidia involucrata Baill in China based on MaxEnt [J]. Chin J Ecol, 2019, 38(4): 1230 − 1237. [19] 崔绍朋, 罗晓, 李春旺, 等. 基于MaxEnt模型预测白唇鹿的潜在分布区[J]. 生物多样性, 2018, 26(2): 171 − 176. CUI Shaopeng, LUO Xiao, LI Chunwang, et al. Predicting the potential distribution of white-lipped deer using the MaxEnt model [J]. Biodiversity Sci, 2018, 26(2): 171 − 176. [20] 喜超, 木霖, 李胜, 等. 基于MaxEnt和ArcGIS预测大薸在云南的潜在适生区[J]. 云南农业大学学报(自然科学), 2018, 33(1): 7 − 16. XI Chao, MU Lin, LI Sheng, et al. MaxEnt modeling and ArcGIS for predicting the potential distribution of Pistia stratiotes L. in Yunnan Province [J]. J Yunnan Agric Univ Nat Sci, 2018, 33(1): 7 − 16. [21] 李丽鹤, 刘会玉, 林振山, 等. 基于MaxEnt和ZONATION的加拿大一枝黄花入侵重点监控区确定[J]. 生态学报, 2017, 37(9): 3124 − 3132. LI Lihe, LIU Huiyu, LIN Zhenshan, et al. Identifying priority areas for monitoring the invasion of Solidago canadensis based on MaxEnt and ZONATION [J]. Acta Ecol Sin, 2017, 37(9): 3124 − 3132. [22] 王茹琳, 李庆, 封传红, 等. 基于MaxEnt的西藏飞蝗在中国的适生区预测[J]. 生态学报, 2017, 37(24): 8556 − 8566. WANG Rulin, LI Qing, FENG Chuanhong, et al. Predicting potential ecological distribution of Locusta migratoria tibetensis in China using MaxEnt ecological niche modeling [J]. Acta Ecol Sin, 2017, 37(24): 8556 − 8566. [23] 王晓玮, 任雪燕, 梁英梅. 基于MaxEnt模型的松针红斑病在中国的潜在分布区及适生性预测分析[J]. 林业科学, 2019, 55(4): 160 − 170. WANG Xiaowei, REN Xueyan, LIANG Yingmei, et al. MaxEnt-based prediction of potential geographic distribution and habitat suitability analysis for Dothistroma pini in China [J]. Sci Silv Sin, 2019, 55(4): 160 − 170. [24] 崔骁芃, 石娟, 王海香, 等. 基于MaxEnt模型的红脂大小蠹在中国适生区的预测[J]. 植物保护学报, 2019, 46(4): 925 − 926. CUI Xiaopeng, SHI Juan, WANG Haixiang, et al. Predicting potential geographical distribution of red turpentine beetle Dendroctonus valens in China based on MaxEnt model [J]. J Plant Prot, 2019, 46(4): 925 − 926. [25] 张日清, 吕芳德, 何方, 等. 美国山核桃引种栽培区划研究(Ⅲ)区划结果与分区描述[J]. 中南林学院学报, 2002, 22(3): 14 − 19. ZHANG Riqing, LÜ Fangde, HE Fang, et al. Cultural regionalization for the introduced pecan tree in China (Ⅲ) region designation and description [J]. J Cent South For Univ, 2002, 22(3): 14 − 19. [26] 程晋昕, 段长春, 闫生杰. 基于MaxEnt模型的薄壳山核桃气候适宜性区划[J]. 应用气象学报, 2020, 31(5): 631 − 640. CHENG Jinxin, DUAN Changchun, YAN Shengjie. Climate suitability regionalization of pecan based on MaxEnt model [J]. J Appl Meteorol Sci, 2020, 31(5): 631 − 640. [27] 郭燕青, 史梦竹, 李建宇, 等. 基于Maxent模型的假臭草潜在分布区预测[J]. 热带亚热带植物学报, 2019, 27(3): 250 − 260. GUO Yanqing, SHI Mengzhu, LI Jianyu, et al. Prediction of potential distribution area of Praxelis clematidea based on Maxent model [J]. J Trop Subtrop Bot, 2019, 27(3): 250 − 260. [28] HIJMANS R J, CAMERON S E, PARRA J L, et al. Very high resolution interpolated climate surfaces for global land areas [J]. Int J Climatol, 2005, 25(15): 1965 − 1978. [29] 张丽霞, 陈晓龙, 辛晓歌. CMIP6情景模式比较计划(ScenarioMIP)概况与评述[J]. 气候变化研究进展, 2019, 15(5): 519 − 525. ZHANG Lixia, CHEN Xiaolong, XIN Xiaoge. Short commentary on CMIP6 Scenario Model Intercomparison Project (ScenarioMIP) [J]. Clim Change Res, 2019, 15(5): 519 − 525. [30] 董波, 李建伟, 林杰, 等. 基于MaxEnt模型预测漆树在中国的潜在适生区研究[J]. 西南林业大学学报(自然科学), 2020, 40(2): 79 − 85. DONG Bo, LI Jianwei, LIN Jie, et al. Predication of potential distribution of Toxicodendron vernicifluum in China based on MaxEnt model [J]. J Southwest For Univ Nat Sci, 2020, 40(2): 79 − 85. [31] 王运生, 谢丙炎, 万方浩, 等. ROC曲线分析在评价入侵物种分布模型中的应用[J]. 生物多样性, 2007, 15(4): 365 − 372. WANG Yunsheng, XIE Bingyan, WAN Fanghao, et al. Application of ROC curve analysis in evaluating the performance of alien species’ potential distribution models [J]. Biodiversity Sci, 2007, 15(4): 365 − 372. [32] 邱浩杰, 孙杰杰, 徐达, 等. 基于MaxEnt模型预测鹅掌楸在中国的潜在分布区[J]. 浙江农林大学学报, 2020, 37(1): 1 − 8. QIU Haojie, SUN Jiejie, XU Da, et al. MaxEnt model-based prediction of potential distribution of Liriodendron chinense in China [J]. J Zhejiang A&F Univ, 2020, 37(1): 1 − 8. [33] 张日清, 吕芳德, 何方, 等. 美国山核桃引种栽培区划研究(Ⅱ)前期引种效果[J]. 中南林学院学报, 2002, 22(2): 17 − 20. ZHANG Riqing, LÜ Fangde, HE Fang, et al. Cultural regionalization for the introduced pecan tree in China (Ⅱ) survey and assessment of previous introduction results [J]. J Cent South For Univ, 2002, 22(2): 17 − 20. [34] 欧阳林男, 陈少雄, 刘学锋, 等. 赤桉在中国的适生地理区域及其对气候变化的响应[J]. 林业科学, 2019, 55(12): 1 − 11. OUYANG Linnan, CHEN Shaoxiong, LIU Xuefeng, et al. Suitable geographic range for Eucalyptus camaldulensis in China and its response to climate change [J]. Sci Silv Sin, 2019, 55(12): 1 − 11. [35] 赵儒楠, 何倩倩, 褚晓洁, 等. 气候变化下千金榆在我国潜在分布区预测[J]. 应用生态学报, 2019, 30(11): 3833 − 3843. ZHAO Runan, HE Qianqian, CHU Xiaojie, et al. Prediction of potential distribution of Carpinus cordata in China under climate change [J]. Chin J Appl Ecol, 2019, 30(11): 3833 − 3843. [36] 李璇, 李垚, 方炎明. 基于优化的Maxent模型预测白栎在中国的潜在分布区[J]. 林业科学, 2018, 54(8): 153 − 164. LI Xuan, LI Yao, FANG Yanming. Prediction of potential suitable distribution areas of Quercus fabri in China based on an optimized Maxent model [J]. Sci Silv Sin, 2018, 54(8): 153 − 164. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20210106
 点击查看大图
点击查看大图

计量
- 文章访问数: 2120
- HTML全文浏览量: 499
- PDF下载量: 59
- 被引次数: 0
