





-
枳椇Hovenia acerba是鼠李科Rhamnaceae枳椇属Hovenia高大落叶乔木,俗称拐枣。枳椇果实利用价值高,其果梗质量占果实总质量的90%。枳椇果梗肥大、肉质多汁、营养丰富,具很高的食用价值[1−6]。光叶毛果枳椇H. trichocarpa var. robusta为枳椇属的变种[7]。叶下面无毛或沿脉被疏柔毛,萼片和果实被锈色密绒毛。肥大果梗含丰富糖,可生食、酿酒、制醋和熬糖等[2, 8]。枳椇果梗是主要食用部位,其滋味主要由非挥发性代谢物主导,不同种枳椇果梗常呈不同风味[1]。卢孟飞等[1]通过比较主栽枳椇品种果梗的营养成分差异发现:不同枳椇品种果梗营养成分和内在品质存在显著差异。因此,为更好地利用枳椇不同种及品种,需对其开展代谢物分析和评价。
广泛靶向代谢组学是一种高通量、高精度和广覆盖的代谢产物检测方法,在茶树Camellia sinensis[9−12]、草莓Fragaria × ananassa[13]、玉米Zea mays[14]和黄纹早竹Phyllostachys violascens var. flavistriatus[15]等植物的化学成分和品质特征研究中应用广泛。PENG等[16]采用LC-ESI-QTOF-MS/MS技术对枳椇果梗进行测定,发现枳椇果梗的抗氧化活性与酚类、单宁酸总质量分数呈正相关,表明酚类物质可能是枳椇果梗抗氧化活性的主要成分,且首次鉴定出枳椇果梗提取物中含有低聚糖、有机酸、脂肪酸和黄酮类等影响风味的化合物。YANG等[17]通过定量分析发现:枳椇果肉的主要营养成分包括糖类、蛋白质、脂质和黄酮等化合物,且糖类十分丰富。
目前,对枳椇研究集中于果梗的代谢物提取和功能食品的开发利用等[17−18],但对枳椇不同种的全面分析报道较少。诸多研究表明:代谢组学可对植物各器官内含物质进行准确解析,并筛选不同种或品种的特征化合物。因此,本研究以枳椇和光叶毛果枳椇果梗为试验材料,通过广泛靶向代谢组学鉴定2种枳椇果梗的代谢物差异,为枳椇种特征化合物的提取与利用及优良品种的选育提供参考。
-
材料选自浙江省丽水市白云山的枳椇和光叶毛果枳椇。枳椇浆果状核果,近球形,无毛(图1A);光叶毛果枳椇浆果状核果,近球形,密被锈色或棕色绒毛(图1B)。2023年11月13日,在2种枳椇树的东、西、南、北4个方位,分别采集完全成熟的果梗,均匀混合后液氮速冻,−80 ℃保存。在液氮环境下用植物粉碎机粉碎,真空冷冻干燥后用于广泛靶向代谢组学测定。
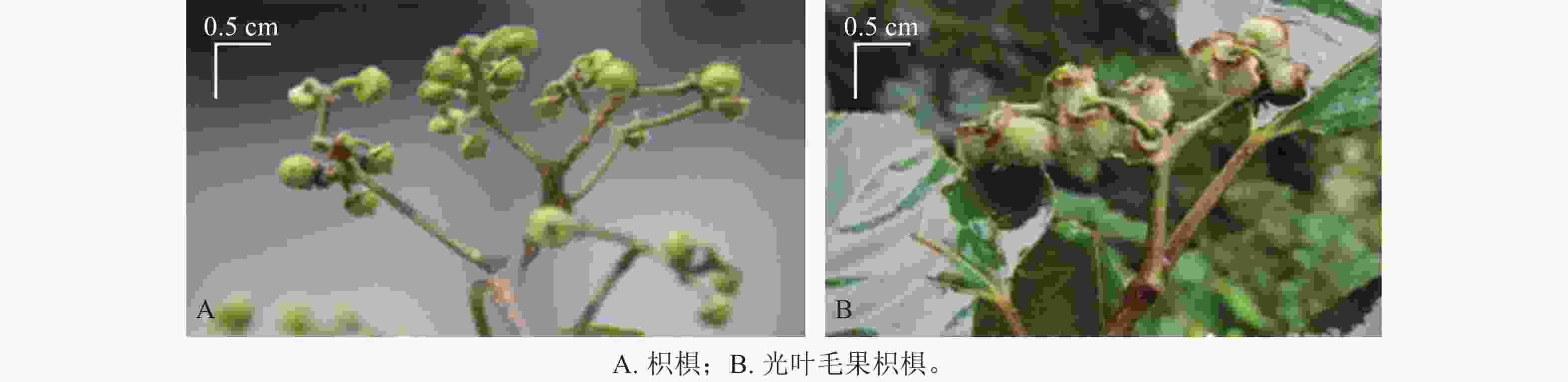
图 1 2种枳椇属植物的果实表型
Figure 1. Fruit phenotypes of two Hovenia species
-
称取50 mg样品粉末,溶解于
1200 μL提取液(−20 ℃预冷的体积分数为70%的甲醇水内标提取液);涡旋6次,每次间隔30 min,每次30 s;离心3 min (12 000 r·min−1,4 ℃),吸取上清液,用0.22 μm微孔滤膜过滤样品,保存于进样瓶,进行超高效液相色谱(UPLC,ExionLC™ AD)和串联质谱(MS/MS,QTRAP® 6500+)分析。色谱条件:Agilent SB-C18 1.8 µm,2.1 mm×100.0 mm,流速0.35 mL·min−1;柱温40 ℃;进样量2 μL。流动相条件:流动相A为超纯水(加入体积分数为0.1%的甲酸),流动相B为乙腈(加入体积分数为0.1%的甲酸)。
-
使用Analyst 1.6.3软件分析质谱数据中的代谢物,得到代谢物列表,包括代谢物名称和丰度。利用KEGG数据库(
https://www.metaboanalyst.ca )对鉴定到的代谢物进行功能和分类注释。使用SIMCA 14.1软件对代谢物进行主成分分析(PCA)和正交偏最小二乘判别分析(OPLS-DA)。其中OPLS-DA 的S-plot图用于识别和解释模型中重要变量的关系,位于右上角和左下角的变量表示其差异显著,红色点通常表示分组贡献值(VIP)≥1的变量,而绿色点表示VIP<1的变量。根据VIP和组间变化的显著性筛选得到差异代谢物。再应用排列检验(permutation test)验证OPLS-DA模型可靠性,通过随机打乱样本的分组标签,重新建立和评估模型,以此来确定模型可靠性,R2为模型的解释率,代表了模型的可解释性,即模型中有多少信息能被解释。其中:R2X表示模型对预测变量(X矩阵)的解释程度;R2Y表示模型对响应变量(Y矩阵)的解释程度;Q2表示模型的预测能力,Q2>0.5被认为是好模型。使用Excel 2019软件对数据进行标准化处理和t检验。接着通过t检验计算得到2组样品间的P值,筛选显著性差异代谢物。使用KEGG对差异代谢物进行通路分析,得到差异代谢通路的富集结果。在本研究中,VIP为变量重要性投影值,表示代谢物对分组的贡献。log2FC表征差异倍数,为每个代谢物在样本组与对照组中3个重复均值的比值,正值表示样本组中代谢物相对含量高于对照组,负值则相反。P值用于判断实验结果的显著性。数据标准化计算公式为:
$$ z=\frac{(x-\mu )}{\sigma }。 $$ 其中:x 是原始数据,μ 是数据的均值,σ 是数据的标准差。
-
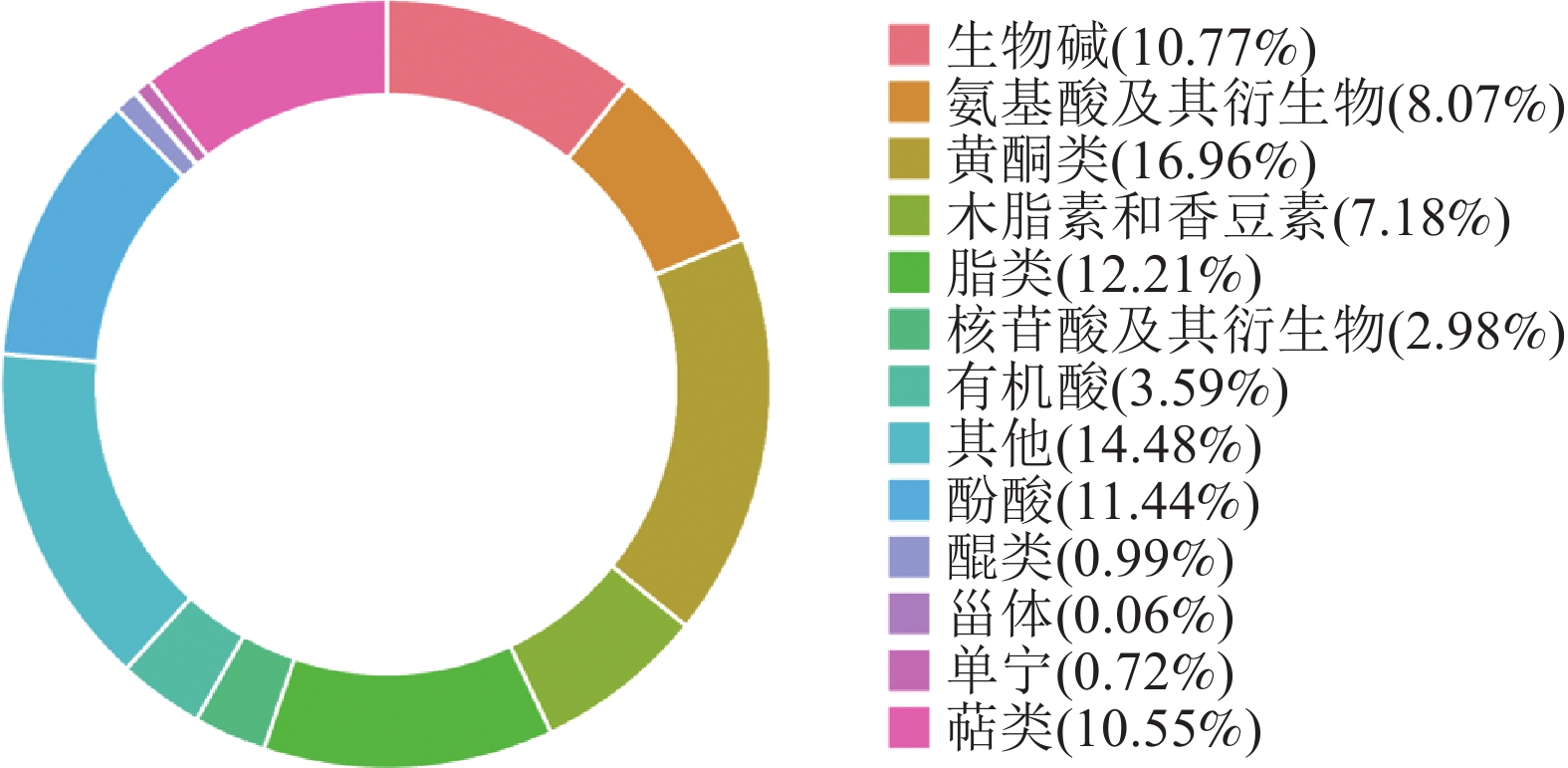
对2种枳椇果梗样品进行UPLC-MS/MS分析,共鉴定出13类1 810种代谢物(图2),包括黄酮类307种(16.96%)、生物碱195种(10.77%)、脂类221种(12.21%)、有机酸65种(3.59%)、萜类191种(10.55%)、单宁13种(0.72%)、甾体1种(0.06%)、醌类18种(0.99%)、酚酸类207种(11.44%)、氨基酸及其衍生物146种(8.07%)、核苷酸及其衍生物54种(2.98%)、木脂素和香豆素130种(7.18%)以及其他代谢物262种(14.48%)。
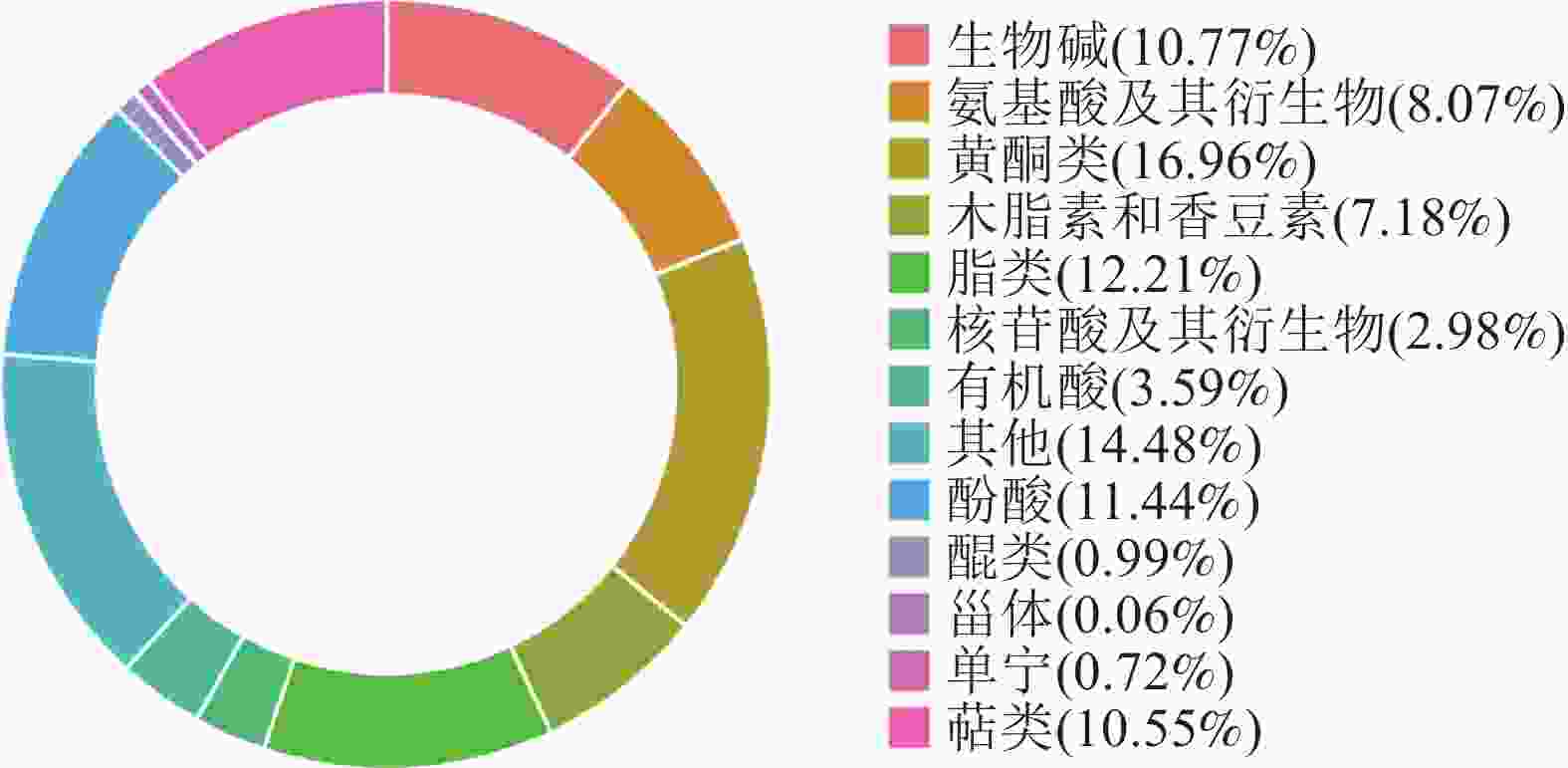
图 2 2种枳椇属植物果梗代谢物分类
Figure 2. Metabolite classification of fruit pedicels in two Hovenia species
-
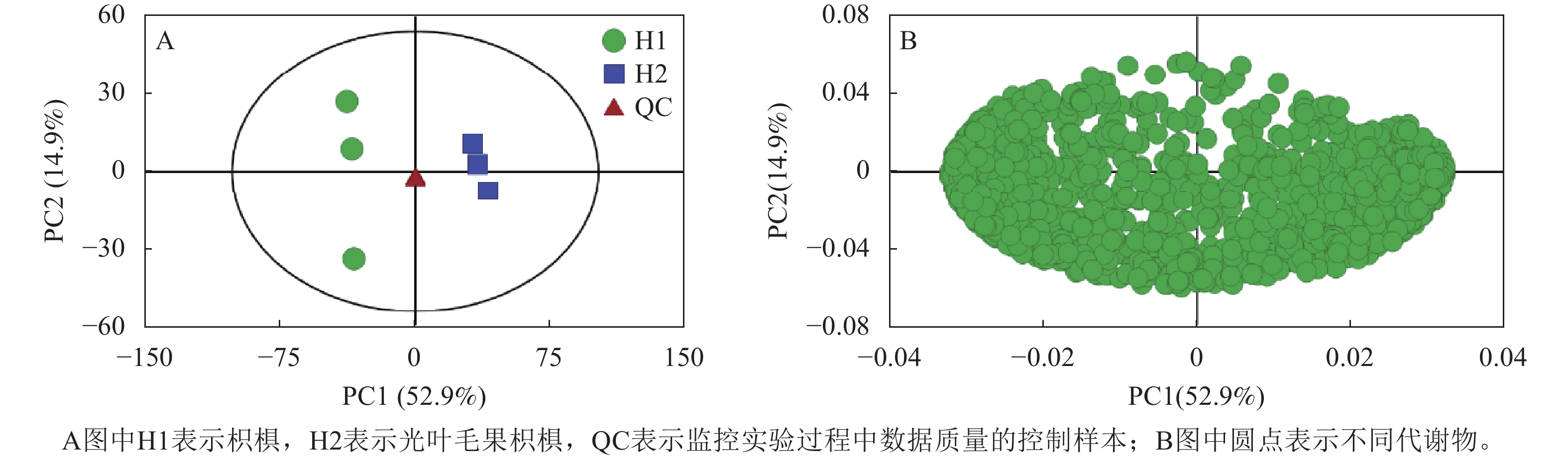
为揭示2种枳椇果梗的代谢物组成差异,采用多维模式识别方法对代谢组数据进行多元统计分析。主成分分析得分表明:2组样品的3个重复良好,代谢组学数据质量可靠(图3A)。PC1和PC2分别解释了总变量的52.9%和14.9%,累计贡献率达67.8%。来自2组的样品表现出明显分层,并在第一主成分上分离显著,表明2组样品的代谢物在组间差异显著。载荷分析确定各代谢物与2个种之间存在相关性(图3B)。
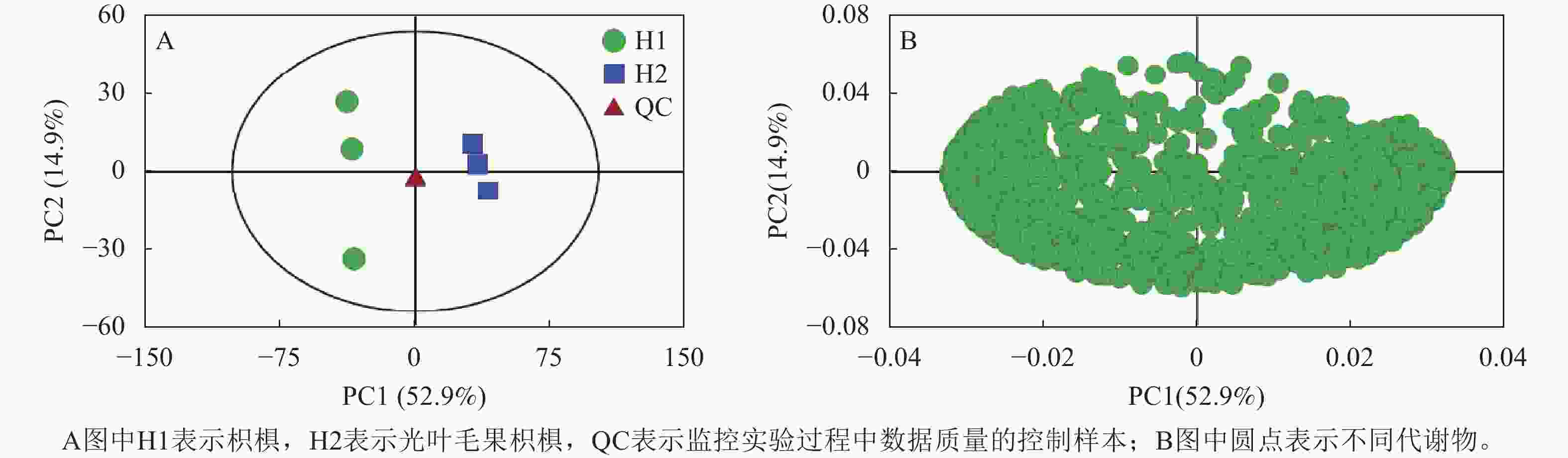
图 3 2种枳椇属植物果梗PCA得分图(A)和载荷图(B)
Figure 3. PCA score diagram (A) and loading diagram (B) of fruit pedicels in two Hovenia species
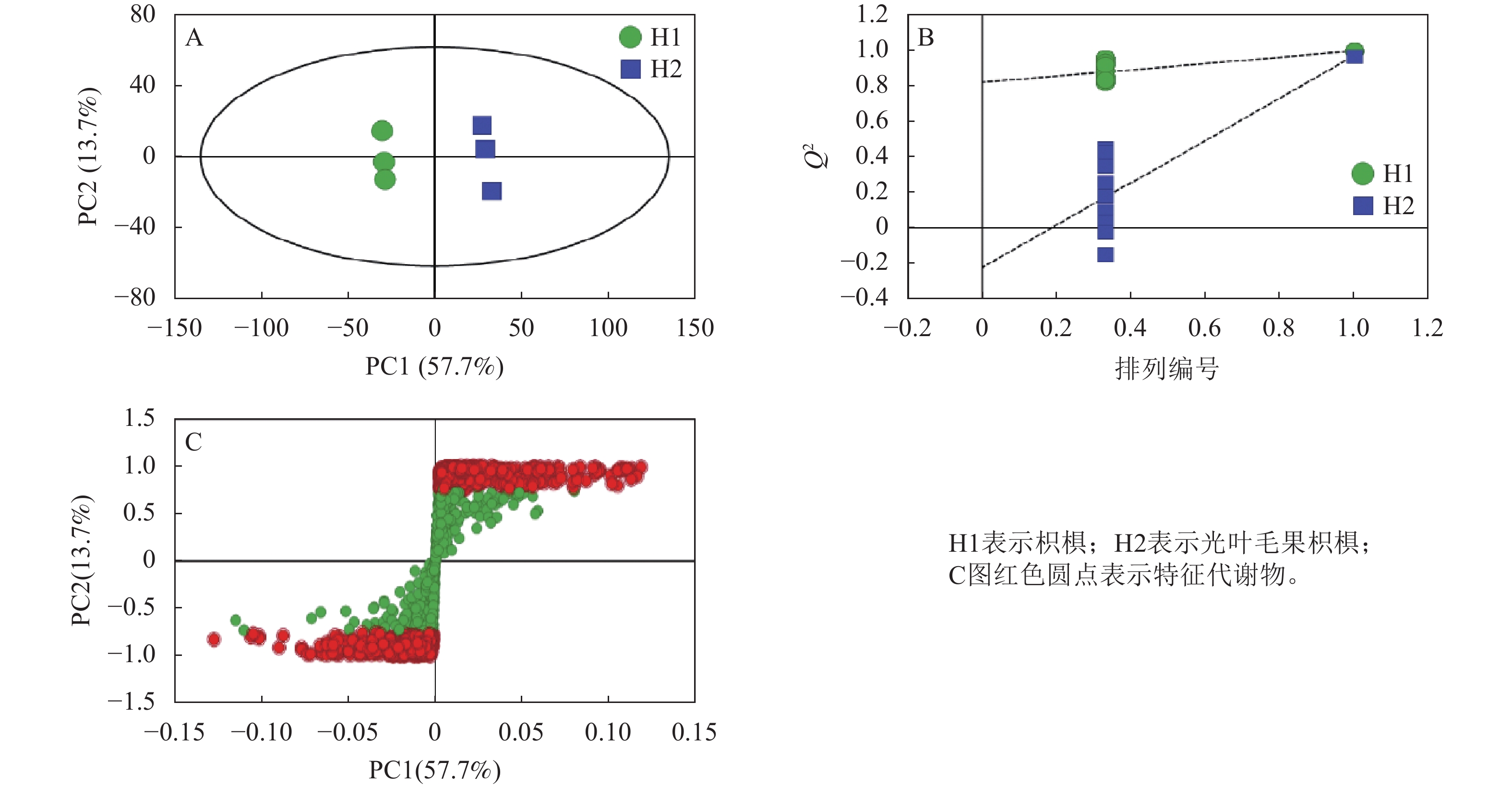
OPLS-DA分析表明:第一主成分和第二主成分的贡献率分别为57.7%和13.7%,累计贡献率为71.4%,可更好地解释总体变量(图4A)。R2X=0.775,R2Y=0.998,Q2=0.972,其中Q2>0.9表明模型可靠(图4B)。S-plot图表示主成分与代谢物的相关系数,可通过变量重要性投影值(VIP)筛选差异代谢物(图4C)。
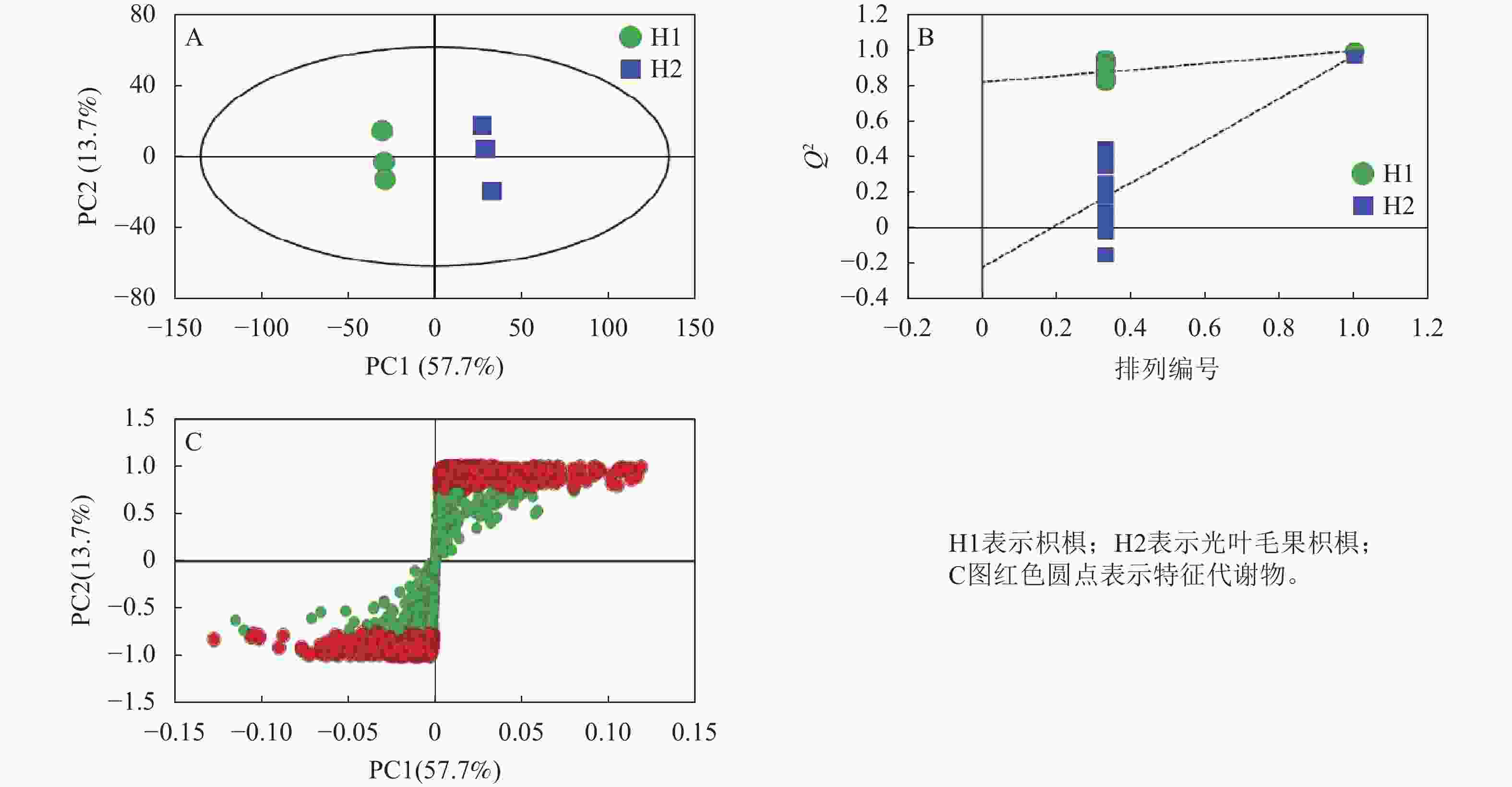
图 4 2种枳椇属植物果梗OPLS-DA图(A)、排序检验图(B)和S-plot图(C)
Figure 4. OPLS-DA score plot (A), ranked test plot (B) and S-plot (C) of fruit pedicels in two Hovenia species
-
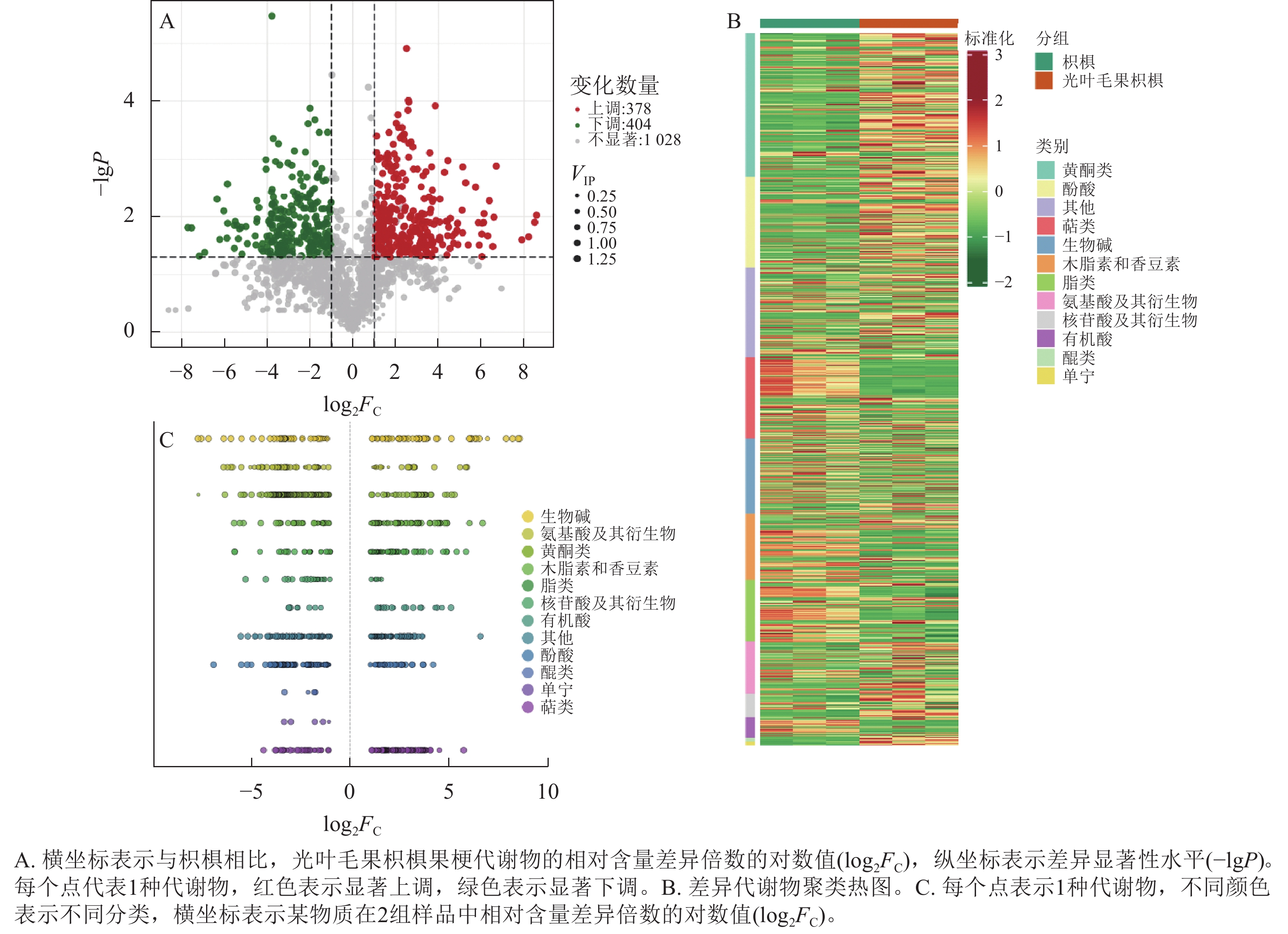
为研究枳椇和光叶毛果枳椇果梗的代谢物差异,以VIP>1、|log2FC|>1和P<0.05为标准筛选出782种差异代谢物。与枳椇果梗相比,378种代谢物在光叶毛果枳椇果梗中显著上调,404种代谢物显著下调(P<0.05,图5A)。为更好地了解差异代谢物种类,对筛选的差异代谢物进行标准化处理,并根据代谢物种类进行聚类分析(图5B)。结果表明:782种差异代谢物中,黄酮和酚酸类种类最多,分别为140和102种,其次为萜类95种、生物碱89种、木脂素和香豆素75种、脂类70种、氨基酸及其衍生物56种、核苷酸及其衍生物27种、有机酸22种、单宁4种、醌类4种和其他类98种。与枳椇果梗相比,单宁、醌类代谢物相对含量在光叶毛果枳椇果梗中显著下调(P<0.05,图5C)。
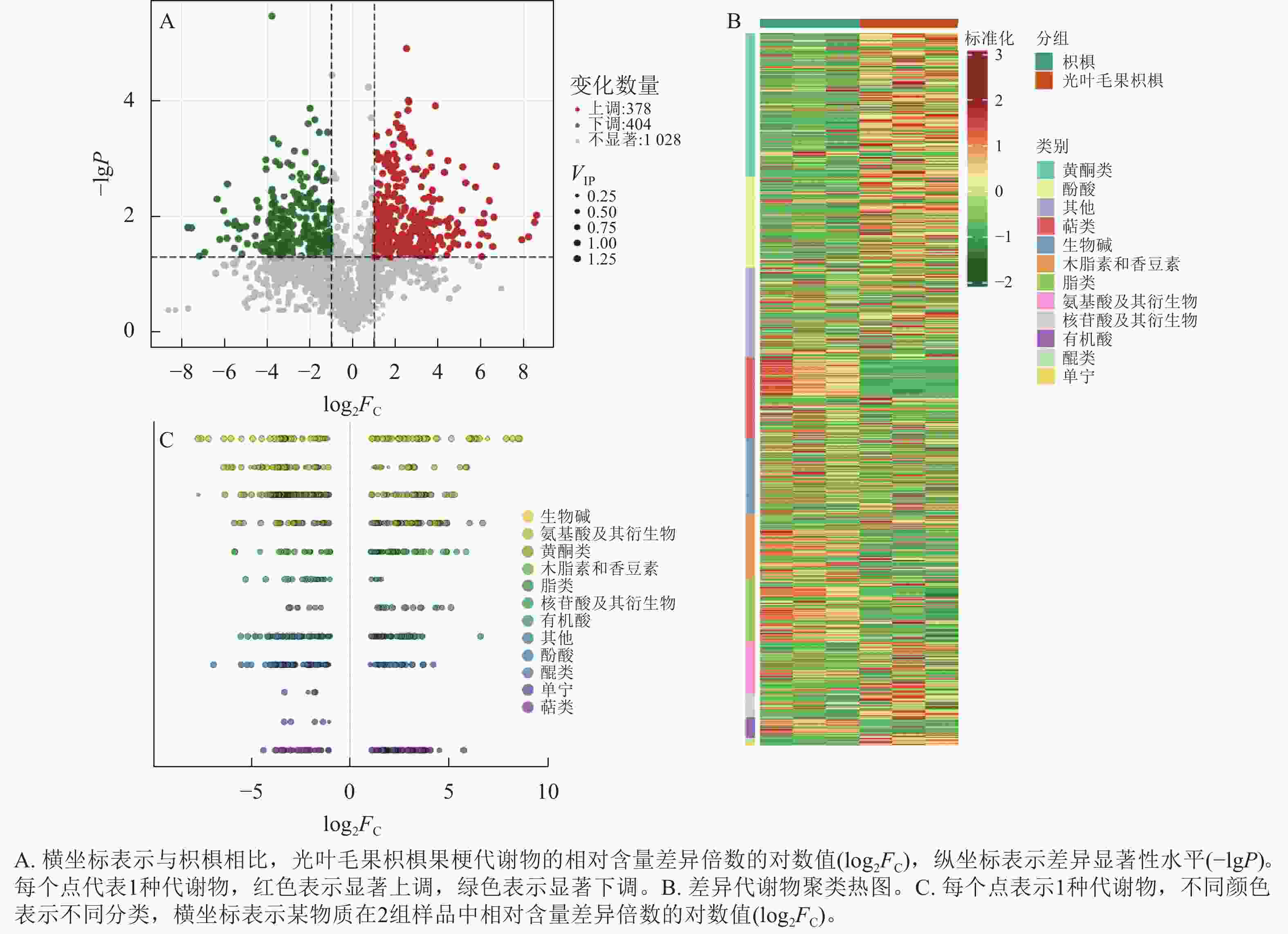
图 5 2种枳椇属植物果梗差异代谢物火山图(A)、差异代谢物聚类热图(B)和差异代谢物散点图(C)
Figure 5. Differential metabolite volcano plot (A), differential metabolite clustering heat map (B) and differential metabolite scatter plot (C) of fruit pedicels in two Hovenia species
-
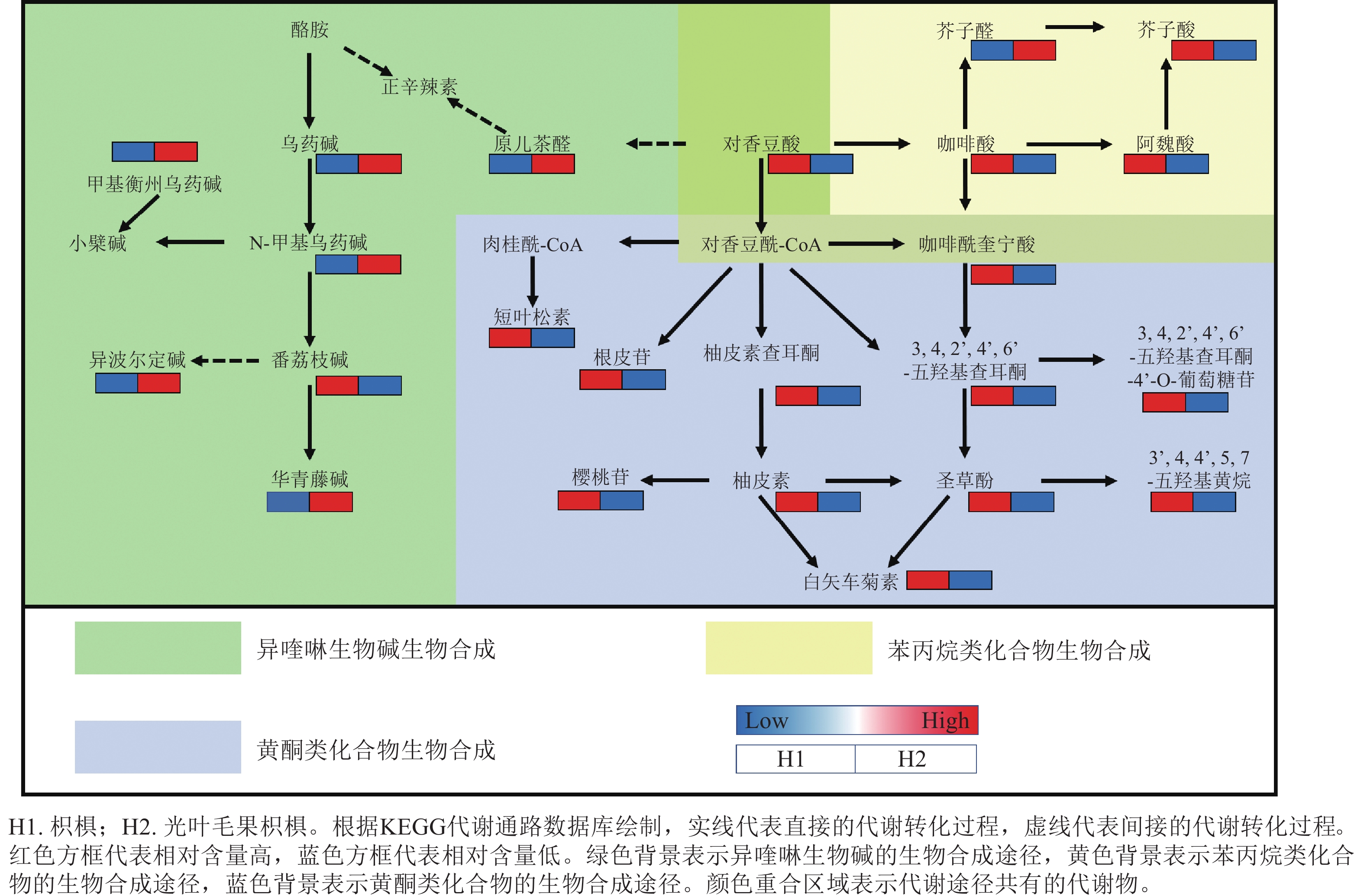
为进一步预测差异代谢物的主要生物学功能,将枳椇和光叶毛果枳椇果梗的差异代谢物注释到KEGG数据库,差异代谢物的代谢通路富集分析筛选到3条差异显著的代谢通路(图6),每条通路中富集的代谢物如表1所示。差异显著的代谢物主要富集在苯丙烷类化合物、黄酮类化合物和异喹啉生物碱的生物合成途径(图7)。在这3条代谢通路中,黄酮类化合物生物合成途径富集的差异代谢物在枳椇果梗中的相对含量显著高于光叶毛果枳椇果梗(P<0.05);在苯丙烷类化合物生物合成途径中,除了芥子醛外,其他富集的差异代谢物在光叶毛果枳椇果梗中的相对含量均显著高于枳椇果梗(P<0.05);在异喹啉生物碱生物合成途径中,除了番荔枝碱外,其他富集的差异代谢物在光叶毛果枳椇果梗中的相对含量均显著高于枳椇果梗(P<0.05)。
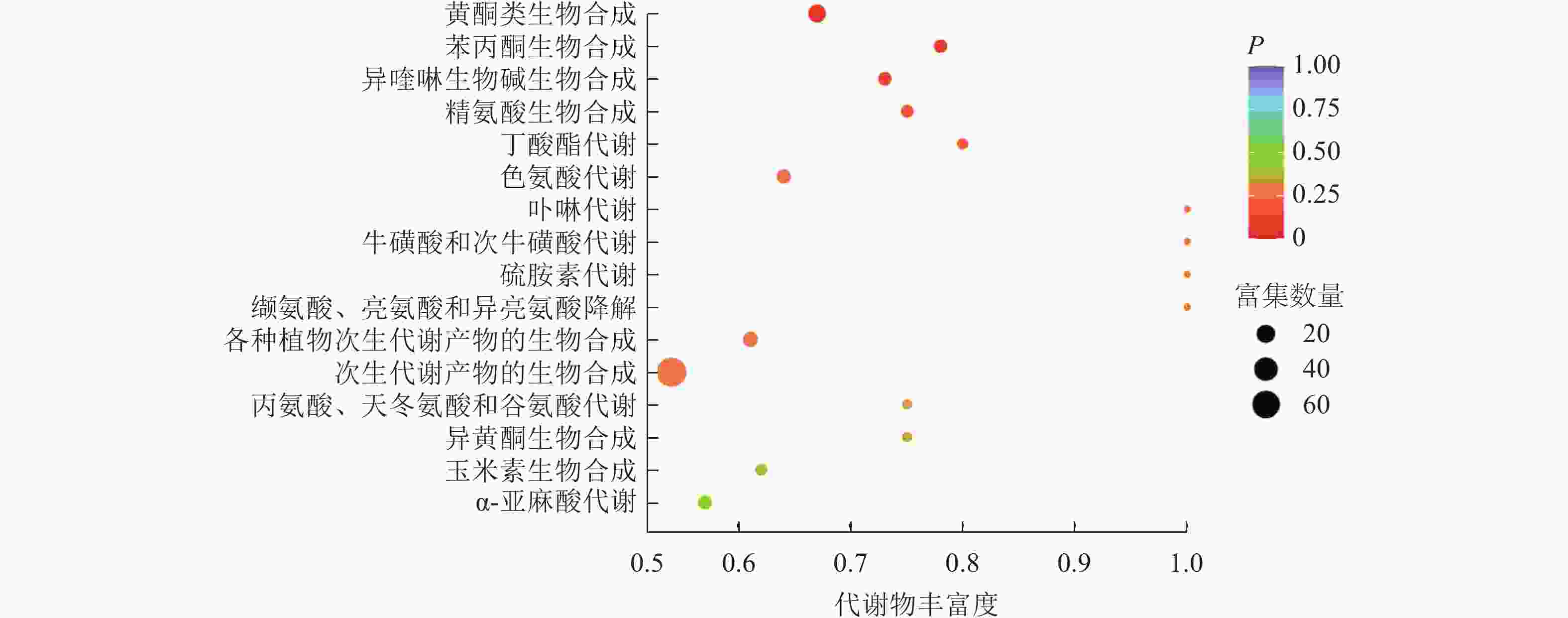
图 6 2种枳椇属植物果梗差异代谢物的富集通路气泡图
Figure 6. Enrichment pathway bubble diagram of fruit pedicels in two Hovenia species
表 1 2种枳椇属植物果梗差异代谢物显著富集通路
Table 1. Metabolites significantly enriched pathways of fruit pedicels in two Hovenia species
通路 通路名称 富集的代谢物名称 Map00940 苯丙烷类化合物生物合成 咖啡酰奎宁酸、对香豆酸、芥子醛、芥子酸、咖啡酸、阿魏酸 Map00941 黄酮类化合物生物合成 咖啡酰奎宁酸、柚皮素查耳酮、柚皮素、3, 4, 2’, 4’, 6’-五羟基查耳酮-4’-O-葡萄糖苷、樱桃苷、
3, 4, 2’, 4’, 6’-五羟基查耳酮、3’, 4, 4’, 5, 7-五羟基黄烷、短叶松素、圣草酚、根皮苷、
白矢车菊素Map00950 异喹啉生物碱生物合成 甲基衡州乌药碱、N-甲基乌药碱、异波尔定碱、华青藤碱、乌药碱、原儿茶醛、
对香豆酸、番荔枝碱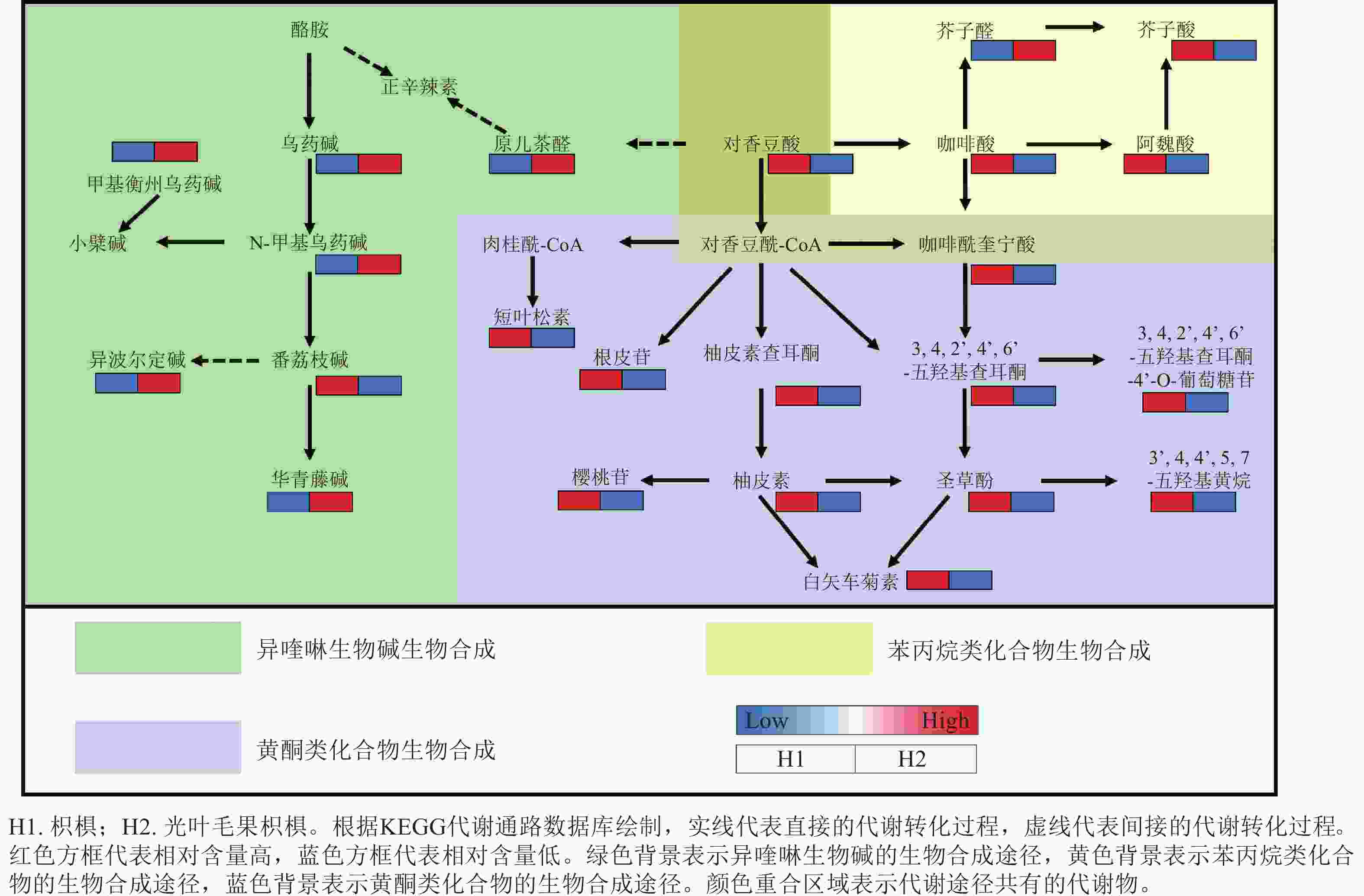
图 7 2种枳椇属植物果梗代谢差异中苯丙烷类化合物、黄酮类化合物和异喹啉生物碱的生物合成通路及相对含量差异
Figure 7. Biosynthesis pathways and contents of phenylpropanoid, flavonoid and isoquinoline alkaloid in fruit pedicels from two Hovenia species
-
本研究对枳椇果梗和光叶毛果枳椇果梗进行广泛靶向代谢组学分析,以探究2种枳椇属植物果梗的代谢物差异,共鉴定出1 810种代谢物,其中枳椇果梗中化合物数量最多的三类分别是黄酮类、脂类和酚酸类化合物。KEGG代谢通路分析发现:2种枳椇果梗差异代谢物苯丙烷类化合物、黄酮类化合物和异喹啉生物碱的生物合成通路富集程度最高。前人研究表明:乌药碱、甲基衡州乌药碱和N-甲基乌药碱等生物碱化合物具有显著镇痛效果[19−20],在临床上被用于缓解疼痛,且具有抗炎、抗高血脂和抗肿瘤等多种药理作用[21]。柚皮素、查尔酮等黄酮类化合物亦具有多种生物活性和药理作用,如较强抗氧化性和抗菌性[22−25],且查尔酮等黄酮类化合物还具有高甜度和低热量[26],可作为食品甜味剂。本研究发现:乌药碱、甲基衡州乌药碱和N-甲基乌药碱等生物碱化合物在光叶毛果枳椇果梗中显著上调,而查耳酮、柚皮素和2’, 3, 4, 4’, 6’-五羟基查耳酮等黄酮化合物在枳椇果梗中显著上调。因此,2种枳椇属植物果梗在潜在应用领域可能存在差异,需进行分类加工利用。
目前,枳椇的应用不再局限于果汁、果酒类产品[27],更多聚焦于果梗中多糖、黄酮和酚酸类化合物的提取和功能性研究[28−31]。例如,黄酮类化合物具有抗菌性[27],枳椇花粉提取物和茎、叶提取物可抑制粪肠球菌Enterococcus faecalis介导的炎症和丙酸杆菌 Cutibacterium acnes引起的痤疮[32−33]。本研究发现:枳椇果梗黄酮类化合物相对含量比光叶毛果枳椇果梗更高,更适合用于提取黄酮类化合物。然而,目前对于枳椇果梗黄酮类化合物抗菌性的研究及其提取物在抗菌上的应用尚欠缺。单宁、醌类具多样生物活性,可用于抗菌和抗病毒作用[34−36],但也存在潜在危害,例如,较高水平的单宁可与消化酶结合,影响消化功能[37],且可与蛋白质形成不溶性复合物,可能会干扰蛋白质和其他营养元素的吸收[38]。部分醌类物质可能对肾脏、肝脏有损害[39],还被认为具有致癌作用[40−41]。因此,在开发利用枳椇产品时,需进行充分的安全性评价。
本研究表明,苯丙烷类化合物、黄酮类化合物和异喹啉生物碱的生物合成途径是2种枳椇果梗中差异最显著的3条代谢通路,且多种中间产物也存在显著差异。这些差异代谢物与果梗感官差异的关系还需更多试验解析。
-
同一生态条件下,2种枳椇果梗的代谢谱存在显著差异,特别是具有药用价值和高甜度的生物碱和黄酮类代谢产物相对含量。这可能与2种枳椇果梗感官质量差异的形成有一定关系。本研究结果可为2种枳椇属植物的特征化合物提取与利用及优良品种的选育提供理论支撑。
Comprehensive targeted metabolomic analysis of fruit stalks in two species of Hovenia plants
-
摘要:
目的 阐明枳椇Hovenia acerba和光叶毛果枳椇H. trichocarpa var. robusta果梗的代谢物差异,为枳椇属Hovenia植物的开发利用奠定基础。 方法 通过高效液相色谱-串联质谱(HPLC-MS/MS)的广泛靶向代谢组学方法,分析枳椇和光叶毛果枳椇果梗中代谢产物和代谢通路的差异。 结果 2种枳椇属植物果梗中共检测到1 810种代谢物,其中显著差异代谢物782种(P<0.05)。与枳椇果梗相比,光叶毛果枳椇果梗中上调差异代谢物有378种,下调差异代谢物有404种,且单宁、醌类代谢物相对含量均显著下调(P<0.05)。2种枳椇属植物果梗中差异代谢物主要富集在苯丙烷类化合物、黄酮类化合物和异喹啉生物碱的生物合成途径,其中极显著差异代谢物包含乌药碱、N-甲基乌药碱和甲基衡州乌药碱等生物碱物质以及柚皮素、柚皮素查尔酮和2’, 3, 4, 4’, 6’-五羟基查耳酮等黄酮类产物。 结论 同一生态条件下,2种枳椇果梗的代谢谱存在显著差异,特别是具有药用价值和高甜度的生物碱和黄酮类代谢产物相对含量存在显著差异。图7表1参41 Abstract:Objective This study aims to clarify the metabolite differences in the fruit stalks of Hovenia acerba and H. trichocarpavar var. robusta, and lays a foundation for the development and utilization of Hovenia plants. Method Through a comprehensive targeted metabolomics approach of high-performance liquid chromatography-tandem mass spectrometry (HPLC-MS/MS), the differences in metabolites and metabolic pathways in the fruit stalks of H. acerba and H. trichocarpa var. robusta were analyzed. Result A total of 1810 metabolites were detected in the fruit stalks of the two Hovenia species, of which 782 metabolites showed significant differences (P<0.05). Compared with the fruit stalks of H. acerba, there were 378 up-regulated differential metabolites and 404 down-regulated differential metabolites in the fruit stalks of H. trichocarpa var. robusta, and the relative contents of tannin and quinone metabolites were significantly down-regulated. The differential metabolites in the fruit stalks of the two Hovenia species were primarily enriched in the biosynthetic pathways of phenylalanine compounds, flavonoids, and isoquinoline alkaloids. Among them, the extremely significant differential metabolites included alkaloid substances such as ligustroside, N-methylligustroside and methylcoclaurine, as well as flavonoids like naringin, naringin chalcone, and 2’, 3, 4, 4’, 6’-pentahydroxy chalcone.Conclusion There are significant differences in the metabolic profiles of the fruit stalks of the two Hovenia species under the same ecological conditions, especially the relative content of alkaloids and flavonoids with medicinal value and high sweetness. [Ch, 7 fig. 1 tab. 41 ref.] -
Key words:
- Hovenia /
- comprehensive targeted metabolome /
- fruit stalk /
- alkaloid /
- flavonoid compound
-
图 2 2种枳椇属植物果梗代谢物分类
Figure 2 Metabolite classification of fruit pedicels in two Hovenia species
图 3 2种枳椇属植物果梗PCA得分图(A)和载荷图(B)
Figure 3 PCA score diagram (A) and loading diagram (B) of fruit pedicels in two Hovenia species
图 4 2种枳椇属植物果梗OPLS-DA图(A)、排序检验图(B)和S-plot图(C)
Figure 4 OPLS-DA score plot (A), ranked test plot (B) and S-plot (C) of fruit pedicels in two Hovenia species
图 5 2种枳椇属植物果梗差异代谢物火山图(A)、差异代谢物聚类热图(B)和差异代谢物散点图(C)
Figure 5 Differential metabolite volcano plot (A), differential metabolite clustering heat map (B) and differential metabolite scatter plot (C) of fruit pedicels in two Hovenia species
图 6 2种枳椇属植物果梗差异代谢物的富集通路气泡图
Figure 6 Enrichment pathway bubble diagram of fruit pedicels in two Hovenia species
图 7 2种枳椇属植物果梗代谢差异中苯丙烷类化合物、黄酮类化合物和异喹啉生物碱的生物合成通路及相对含量差异
Figure 7 Biosynthesis pathways and contents of phenylpropanoid, flavonoid and isoquinoline alkaloid in fruit pedicels from two Hovenia species
表 1 2种枳椇属植物果梗差异代谢物显著富集通路
Table 1. Metabolites significantly enriched pathways of fruit pedicels in two Hovenia species
通路 通路名称 富集的代谢物名称 Map00940 苯丙烷类化合物生物合成 咖啡酰奎宁酸、对香豆酸、芥子醛、芥子酸、咖啡酸、阿魏酸 Map00941 黄酮类化合物生物合成 咖啡酰奎宁酸、柚皮素查耳酮、柚皮素、3, 4, 2’, 4’, 6’-五羟基查耳酮-4’-O-葡萄糖苷、樱桃苷、
3, 4, 2’, 4’, 6’-五羟基查耳酮、3’, 4, 4’, 5, 7-五羟基黄烷、短叶松素、圣草酚、根皮苷、
白矢车菊素Map00950 异喹啉生物碱生物合成 甲基衡州乌药碱、N-甲基乌药碱、异波尔定碱、华青藤碱、乌药碱、原儿茶醛、
对香豆酸、番荔枝碱 下载: 导出CSV
下载: 导出CSV
-
[1] 卢孟飞, 陈瞳晖, 莫双凤, 等. 拐枣主栽品种果实的经济性状和营养成分分析及综合评价[J]. 经济林研究, 2024, 42 (2): 1−10. LU Mengfei, CHEN Tonghui, MO Shuangfeng, et al. Comprehensive evaluation and analysis on fruit economic characteristics and nutrient composition of Hovenia varieties [J]. Non-wood Forest Research, 2024, 42 (2): 112−121. [2] 向进乐, 杜琳, 朱文学, 等. 枳椇果梗营养功能与加工利用[J]. 中国林副特产, 2015(1): 98−101. XIANG Jinle, DU Lin, ZHU Wenxue, et al. Research progress in nutrition and utilization of Hovenia peduncles [J]. Forest By-Product and Speciality in China, 2015(1): 98−101. [3] 谢志民. 枳椇和枳椇子的本草考证[J]. 中药材, 1994, 17(6): 44−45, 56. XIE Zhimin. Herbological study of “Zhiju” and “Zhijuzi” [J]. Journal of Chinese Medicinal Materials, 1994, 17(6): 44−45, 56. [4] 陆石英, 覃志高. 药食两用枳椇的研究进展[J]. 食品安全质量检测学报, 2020, 11(6): 1865−1870. LU Shiying, QIN Zhigao. Research progress on Hovenia dulcis Lindl. for medicine and food [J]. Journal of Food Safety and Quality, 2020, 11(6): 1865−1870. [5] 徐方方, 刘博, 张晓琦. 枳椇属化学成分和药理活性的研究进展[J]. 中国中药杂志, 2020, 45(20): 4827−4835. XU Fangfang, LIU Bo, ZHANG Xiaoqi. Research progress on chemical constituents and pharmacological activities of Hovenia [J]. China Journal of Chinese Materia Medica, 2020, 45(20): 4827−4835. [6] WANG Mingchun, ZHU Peilei, JIANG Changxing, et al. Preliminary characterization, antioxidant activity in vitro and hepatoprotective effect on acute alcohol-induced liver injury in mice of polysaccharides from the peduncles of Hovenia dulcis [J]. Food and Chemical Toxicology, 2012, 50(9): 2964−2970. [7] 嵇扬, 陆红. 枳椇子研究进展[J]. 中草药, 2002, 33(9): 102−104. JI Yang, LU Hong. Research progress on Hovenia dulcis Thunb. [J]. Chinese Traditional and Herbal Drugs, 2002, 33(9): 102−104. [8] 时涛, 王晓玲, 陈振德, 等. 枳椇子化学成分及其药理活性研究进展[J]. 中药材, 2006, 29(5): 510−513. SHI Tao, WANG Xiaoling, CHEN Zhende, et al. Research progress of chemical constituents and pharmacological activities of Hovenia dulcis [J]. Journal of Chinese Medicinal Materials, 2006, 29(5): 510−513. [9] 朱婉, 吴颖, 黎晓湘, 等. 基于广泛靶向代谢组学结合高效液相色谱法分析‘紫娟’和‘迎霜’茶树花代谢物差异[J]. 浙江大学学报(农业与生命科学版), 2023, 49(6): 825−839. ZHU Wan, WU Ying, LI Xiaoxiang, et al. Analysis of differential metabolites between ‘Zijuan’ and ‘Yingshuang’ tea flowers based on widely targeted metabolomics combined with high performance liquid chromatography [J]. Journal of Zhejiang University (Agriculture & Life Sciences), 2023, 49(6): 825−839. [10] WEI Kailing, LIU Meiya, SHI Yifan, et al. Metabolomics reveal that the high application of phosphorus and potassium in tea plantation inhibited amino-acid accumulation but promoted metabolism of flavonoid [J/OL]. Agronomy, 2022, 12 (5): 1086[2024-06-05]. DOI: 10.3390/agronomy12051086. [11] ZHANG Hua, SONG Yakang, FAN Zhenlei, et al. Aluminum supplementation mediates the changes in tea plant growth and metabolism in response to calcium stress [J/OL]. International Journal of Molecular Sciences, 2024, 25 (1): 530[2024-06-05]. DOI: 10.3390/ijms25010530. [12] ZHANG Hua, LI Chunlei, WEI Kailing, et al. The reduction of tea quality caused by irrational phosphate application is associated with anthocyanin metabolism [J/OL]. Beverage Plant Research, 2023, 3 : 10[2024-06-05]. DOI: 10.48130/BPR-2023-0010. [13] AKHATOU I, SAYAGO A, GONZÁLEZ-DOMÍNGUEZ R, et al. Application of targeted metabolomics to investigate optimum growing conditions to enhance bioactive content of strawberry [J]. Journal of Agricultural and Food Chemistry, 2017, 65(43): 9559−9567. [14] YANG Ruichun, LI Yunfeng, ZHANG Yuanyuan, et al. Widely targeted metabolomics analysis reveals key quality-related metabolites in kernels of sweet corn [J/OL]. International Journal of Genomics, 2021, 2021 (1): 2654546[2024-06-05]. DOI: 10.1155/2021/2654546. [15] QIAN Qixia, ZHANG Huicong, ZHANG Pengwei, et al. Integrated transcriptomics and metabolomics provide new insights into the leaf coloration of a bamboo variant Phyllostachys violascens var. flavistriatus [J/OL]. Scientia Horticulturae, 2024, 334 : 113330[2024-06-05]. DOI: 10.1016/j.scienta.2024.113330. [16] PENG Han, DENG Zeyuan, CHEN Xuan, et al. Major chemical constituents and antioxidant activities of different extracts from the peduncles of Hovenia acerba Lindl. [J]. International Journal of Food Properties, 2018, 21(1): 2135−2155. [17] YANG Zhijian, CHEN Hui, LIN Chaimei, et al. Comprehensive evaluation of quality traits of Hovenia acerba germplasm resources in Fujian Province [J/OL]. Forests, 2023, 14 (2): 204[2024-06-05]. DOI: 10.3390/f14020204. [18] NIIYA M, SHIMATO Y, OHNO T, et al. Effects of Hovenia dulcis fruit and peduncle extract on alcohol metabolism [J/OL]. Journal of Ethnopharmacology, 2024, 321 : 117541[2024-06-05]. DOI: 10.1016/j.jep.2023.117541. [19] 杨舜伊, 袁纯红, 蒋高华, 等. 乌头属植物生物碱研究新进展[J]. 湖北农业科学, 2020, 59(23): 5−10. YANG Shunyi, YUAN Chunhong, JIANG Gaohua, et al. Recent advance of alkaloids in genus Aconitum [J]. Hubei Agricultural Sciences, 2020, 59(23): 5−10. [20] 黄茜, 孙明丽, 李腾飞, 等. 乌头生物碱镇痛作用及机制研究进展[J]. 神经药理学报, 2017, 7(3): 21−32. HUANG Qian, SUN Mingli, LI Tengfei, et al. Research progress on mechanisms underlying aconitines analgesia [J]. Journal of Neuropharmacology, 2017, 7(3): 21−32. [21] 邢梦雨, 田崇梅, 夏道宗. 乌药化学成分及药理作用研究进展[J]. 天然产物研究与开发, 2017, 29(12): 2147−2151. XING Mengyu, TIAN Chongmei, XIA Daozong. Review on chemical constituents and pharmacological effects of Lindera aggregata Kosterm [J]. Natural Product Research and Development, 2017, 29(12): 2147−2151. [22] DUDA-MADEJ A, STECKO J, SOBIERAJ J, et al. Naringenin and its derivatives: health-promoting phytobiotic against resistant bacteria and fungi in humans [J/OL]. Antibiotics, 2022, 11 (11): 1628[2023-06-05]. DOI: 10.3390/antibiotics11111628. [23] PEREIRA R, SILVA A M S, RIBEIRO D, et al. Bis-chalcones: a review of synthetic methodologies and anti-inflammatory effects [J/OL]. European Journal of Medicinal Chemistry, 2023, 252 : 115280[2024-06-05]. DOI: 10.1016/j.ejmech.2023.115280. [24] RAMMOHAN A, REDDY J S, SRAVYA G, et al. Chalcone synthesis, properties and medicinal applications: a review [J]. Environmental Chemistry Letters, 2020, 18(2): 433−458. [25] UÇAR K, GÖKTAŞ Z. Biological activities of naringenin: a narrative review based on in vitro and in vivo studies [J]. Nutrition Research, 2023, 119: 43−55. [26] 彭颖, 潘思轶, 张德新. 柚皮苷二氢查尔酮的制备及其呈甜机理研究[J]. 食品与发酵工业, 2022, 48(1): 21−28. PENG Ying, PAN Siyi, ZHANG Dexin. Preparation and the mechanism of sweet taste of naringin dihydrochalcone [J]. Food and Fermentation Industries, 2022, 48(1): 21−28. [27] 黎继烈. 枳椇果酒的酿制工艺[J]. 食品与发酵工业, 2002(12): 71−73. LI Jilie. The brewing process of Hovenia dulcis fruit wine [J]. Food and Fermentation Industries, 2002(12): 71−73. [28] 向进乐, 李志西, 甘峰, 等. 枳椇果梗不同类型酚酸含量及抗氧化活性[J]. 食品与发酵工业, 2011, 37(8): 106−111. XIANG Jinle, LI Zhixi, GAN Feng, et al. Determination of four different phenolic acids of Hovenia acerba fruit and their antioxidant activities [J]. Food and Fermentation Industries, 2011, 37(8): 106−111. [29] 张玉超, 张智淮, 刘良禹, 等. 枳椇果梗黄酮的提取及其抗氧化性[J]. 食品科技, 2024, 49(4): 251−260. ZHANG Yuchao, ZHANG Zhihuai, LIU Liangyu, et al. Extraction and antioxidant activity of flavone from Hovenia dulcis peduncle [J]. Food Science and Technology, 2024, 49(4): 251−260. [30] 刘旭东, 张玉超, 朱思洁, 等. 枳椇果梗多糖的提取工艺优化及其抗氧化性[J]. 食品工业科技, 2023, 44(11): 230−237. LIU Xudong, ZHANG Yuchao, ZHU Sijie, et al. Optimization of extraction process of polysaccharides from Hovenia dulcis fruit pedicels and its antioxidant activity [J]. Science and Technology of Food Industry, 2023, 44(11): 230−237. [31] 刘菁华, 骆洁雅, 郭鹏, 等. 枳椇子黄酮的含量测定及其抗氧化作用的分子机制[J]. 中国兽医杂志, 2024, 60(2): 120−125. LIU Jinghua, LUO Jieya, GUO Peng, et al. Content determination of flavonoids in Hovenia dulcis and its molecular mechanism of antioxidant effect [J]. Chinese Journal of Veterinary Medicine, 2024, 60(2): 120−125. [32] CHOI H A, AHN S O, LIM H D, et al. Growth suppression of a gingivitis and skin pathogen Cutibacterium (Propionibacterium) acnes by medicinal plant extracts [J/OL]. Antibiotics, 2021, 10 (9): 1092[2024-06-05]. DOI: 10.3390/antibiotics10091092. [33] YOU R, KWON O Y, WOO H J, et al. Hovenia monofloral honey can attenuate Enterococcus faecalis mediated biofilm formation and inflammation [J]. Food Science of Animal Resources, 2022, 42(1): 84−97. [34] 杜国成. 中药鞣质成分的药理作用探析[J]. 中国医药科学, 2011, 1 (16): 27, 33. DU Guocheng. Pharmacological effects analysis of tannins in traditional chinese medicine [J]. China Medicine and Pharmacy, 2011, 1 (16): 27, 33. [35] WANG Peng, WEI Jia, HUA Xin, et al. Plant anthraquinones: classification, distribution, biosynthesis, and regulation [J/OL]. Journal of Cellular Physiology, 2023, 239 : e31063[2024-06-05]. DOI: 10.1002/jcp.31063. [36] ZHAO Lihua, ZHENG Lin. A review on bioactive anthraquinone and derivatives as the regulators for ROS [J/OL]. Molecules, 2023, 28 (24): 8139[2024-06-05]. DOI: 10.3390/molecules28248139. [37] GRIFFITHS D W. Nutritional and Toxicological Significance of Enzyme Inhibitors in Foods [M]. Boston: Springer, 1986: 509−516. [38] van BUREN J P, ROBINSON W B. Formation of complexes between protein and tannic acid [J]. Journal of Agricultural and Food Chemistry, 1969, 17(4): 772−777. [39] SONG Shiming, GAO Yanxia, FENG Shuai, et al. Widespread occurrence of two typical N, N’-substituted p-phenylenediamines and their quinones in humans: association with oxidative stress and liver damage [J/OL]. Journal of Hazardous Materials, 2024, 468 : 133835[2024-06-05]. DOI: 10.1016/j.jhazmat.2024.133835. [40] CAVALIERI E, ROGAN E. The 3, 4-quinones of estrone and estradiol are the initiators of cancer whereas resveratrol and N-acetylcysteine are the preventers [J/OL]. International Journal of Molecular Sciences, 2021, 22 (15): 8238[2024-06-05]. DOI: 10.3390/ijms22158238. [41] ZAMORA R, HIDALGO F J. Formation of naphthoquinones and anthraquinones by carbonyl-hydroquinone/benzoquinone reactions: a potential route for the origin of 9, 10-anthraquinone in tea [J/OL]. Food Chemistry, 2021, 354 : 129530[2024-06-05]. DOI: 10.1016/j.foodchem.2021.129530. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20240429
 点击查看大图
点击查看大图

计量
- 文章访问数: 2765
- HTML全文浏览量: 767
- PDF下载量: 30
- 被引次数: 0
