




 下载:
下载:


-
三江源素有“中华水塔”[1]之称,在涵养水源、保护生物多样性、固定碳素等方面有重要的生态价值[2]。然而近年来受全球气候变化和人类过度活动等影响,三江源高寒草甸普遍退化,甚至部分优良草地逐步退化为次生裸地或“黑土滩”[3],引发了物种多样性减少、水源涵养功能下降及毒杂草入侵等一系列生态问题[4],严重制约当地畜牧业可持续发展[2]。因此,解析三江源高寒草甸恢复限制因子及研发生态恢复技术是目前亟待解决的关键问题[5]。
草地退化是植被、土壤以及能流共同退化的生态逆向演替过程结果[6]。土壤代谢物是土壤中植物根系分泌、微生物自身内源性代谢物释放及对环境物质的代谢分解产生的重要小分子物质,是植物-土壤-微生物共同作用的结果[7],在植物生长、营养平衡、物质交换和能量转化、生物化学循环等过程起关键作用[8],是评价土壤质量的重要指标,也能直接反映土壤退化状况[9]。因此,研究土壤代谢物可为明确土壤-植物-微生物交互反馈作用的重要因素和信号分子提供物质基础[10]。土壤退化是草地生态系统退化的核心,整合土壤非生物因素(理化指标、酶活性、土壤代谢物等)和生物因素(土壤微生物群落组成及功能)研究,可以更加全面地了解草地退化程度[11−12]。
现有研究主要关注退化草地植物群落及群落特征[13−14]、土壤理化因子[2, 6]、土壤养分[15]、酶活性[16]、微生物多样性以及部分指标相关性[17]等,发现随着草地退化的加剧,土壤抗侵蚀能力、有机质、全氮及速效钾含量呈先增加后降低的趋势,土壤酶活性(土壤脲酶活性)、植被生物量及土壤微生物数量(真菌数量)显著下降;同时,退化通过地上生物量、土壤含水量、pH、总有机碳、全氮、铵态氮、速效磷和全钾含量及有效氮磷比等环境因子显著驱动土壤微生物格局和功能改变[18−20]。然而,草地生态系统尤其退化高寒草甸土壤系统的代谢组学研究相对匮乏。马源[10]研究发现:祁连山东缘退化显著影响了优势植物分泌的代谢物种类和含量,酚类和有机酸等化合物为适应生存环境的变化而释放的主要化学物质。白炜等[21]发现:增温显著改变高寒沼泽草甸根际土壤蔗糖酶和纤维素酶活性,改变了根际土壤代谢物组成及网络结构,且与酶活性高度相关。因此,研究退化草地土壤代谢产物特征及其与环境因子的相关性,可为高寒草甸退化的关键过程的解析及草地土壤环境的信息与物质互换协同机制研究提供理论依据[12]。
非靶向代谢组学(NTM)通过对小分子量代谢产物进行定性与定量分析,直接准确地映射土壤及相关生物体的生理状态,是研究生物学样本的相似性、聚类及差异的重要方法。超高效液相色谱与高分辨率质谱联用技术(UHPLC-LC-MS/MS)因分离性强、灵敏度高,广泛用于分析植物体内及根际土壤代谢产物并预测其可能的代谢途径和机制[22]。近年来土壤代谢组学在土壤微生态研究中的应用日趋广泛,通过识别代谢途径中的生化中间产物,提高对土壤生物过程的理解及预测能力[23],已成为当前研究的热点。张小芳等[24]研究表明:混播草地能刺激植物分泌更多的有益土壤代谢物(如海藻糖和缬氨酸),通过招募类诺卡氏重塑细菌群落并促进生态系统健康;程锦春等[25]研究表明:青藏高原药用植物红景天Rhodiola rosea、珠芽蓼Polygonum viviparum和瑞香狼毒Stellera chamaejasme的根际土壤标志代谢物与其药效成分密切相关,其代谢途径是生物合成活性次生代谢产物以适应高寒生态环境的重要基础。
为了系统阐释不同退化程度的高寒草甸土壤代谢产物差异特征,厘清土壤代谢产物与土壤环境非生物因子的相关性,本研究聚焦青海省果洛藏族自治州玛沁县三江源国家公园不同退化程度的高寒草甸,解析土壤代谢物的组成和差异代谢产物特征,探明与退化相关的标志性土壤代谢产物及其关键代谢富集通路,并揭示影响土壤代谢物的关键环境驱动因子。
-
研究区位于青海省果洛藏族自治州玛沁县大武镇(34°27′N,100°13′E),海拔为3 740.0 m,主要土壤类型为高山草甸土。属于典型的高原大陆性气候,昼夜温差大,年均气温−3.9 ℃,年降水量513.2~542.9 mm,多集中在6—9月,年蒸发量为2 471.6 mm,牧草生长时间为156.0 d,全年无绝对无霜期[6]。当地草地类型为高寒嵩草草甸,研究区以小嵩草Kobresia pygmaea、矮嵩草Kobresia humilis、针茅Stipa sp.为建群种,同时伴有大量鹅绒委陵菜Potentilla anserina、细叶亚菊Ajania tenuifolia和铁棒锤 Aconitum pendulum等毒杂草[26]。根据天然草地退化等分级指标(GB19377—2003)和文献[27]的方法,在植物生长旺盛期根据优势植物、群落、毒杂草、盖度等数据,将高寒草甸分为未退化草甸(none degradation meadow,NDM)、轻度退化草甸(light degradation meadow,LDM)、中度退化草甸(moderate degradation meadow,MDM)以及重度退化草甸(severe degradation meadow,SDM) [28](表1)。
表 1 样地基本情况
Table 1. Characteristics of basic information
样地 海拔/m 经纬度 优势物种 盖度/% 高度/cm 地上生物量/
(g·m−2)地下生物量/
(g·m−2)未退化草甸
(NDM)3 940 34°21′23″N
100°29′13″E线叶嵩草、 高山嵩草、
矮嵩草、干生苔草91.89±1.69 a 21.35±1.32 a 717.59±88.14 b 3 051.60±236.31 a 轻度退化草甸
(LDM)3 960 34°21′20″N
100°28′47″E线叶嵩草, 高山嵩草、
异针茅、珠芽蓼76.50±4.67 b 18.63±2.36 b 887.31±20.13 a 2 278.52±114.11 b 中度退化草甸
(MDM)3 950 34°21′39″N
100°27′40″E线叶嵩草、高山嵩草、矮嵩草、
鹅绒委陵菜、黄帚橐吾63.48±7.05 c 17.32±2.59 b 551.29±30.34 c 1 620.87±78.38 c 重度退化草甸
(SDM)3 950 34°21′10″N
100°28′48″E线叶嵩草、高山嵩草、矮嵩草、
黄帚橐吾、细叶亚菊、铁棒锤30.50±9.88 d 3.16±0.08 c 448.54±84.62 c 1 261.17±25.74 d 说明:线叶嵩草Koberesia capillifolia,高山嵩草 Kobreaia pygmaea,干生苔草Carex aridula,异针茅Stipa aliena,珠芽蓼Polygonum viviparum,黄帚橐吾 Ligularia virgaurea。 -
于2020年8月中上旬植物生长旺盛期,按随机取样法进行采样。每个退化区域设3个30 m×30 m随机采样区,每个采样区域内随机设置5个1 m×1 m重复样方。在样方内记录植物种类、物种数、群落总盖度和高度,采用刈割法测定植被群落地上部生物量。去除草地表面凋落物层后,每个小样方分别挖取1个0.15 m×0.15 m×0.20 m (长×宽×高)的土块,采用抖落法[7]收集土壤,将每个退化样地中5个土壤样本均匀混合为1个复合土样,共计12个土样。将每个土样分为3份:一份测定土壤含水量;一份土样过1.00和0.25 mm土筛后进行土壤理化性质及酶活性分析;另一份分别取5 g左右的新鲜土壤样品,液氮速冻15 min,−80 ℃保存,用于土壤代谢物检测分析。
-
土壤理化性质和酶活性的测定参照JIN等[27]和景媛媛等[29]的测定方法。土壤pH采用电位法,土壤含水量(SMC)采用烘干法,土壤有机质(SOM)采用重铬酸钾外加热法,土壤全氮(TN)采用凯氏定氮法,全磷(TP)和速效磷(AP)采用钼锑抗显色法,全钾(TK)和速效钾(AK)采用火焰光度法,土壤硝态氮(NN)采用氯化钾浸提-靛酚蓝比色法。土壤脲酶(UE)活性采用苯酚钠-次氯酸钠比色法,中性磷酸酶(NP)采用磷酸苯二钠比色法,蔗糖酶(SC)采用3,5-二硝基水杨酸显色法。
-
称取不同样本新鲜土壤各0.50 g,土壤样品前处理的方法参考文献[25]。另外,每个样本分别移取20 μL上清液,混合后作为质控样品(QC)进样时在样本队列的前中后插入[30−31]。
-
采用 Nexera UPLC 超高效液相串联系统,联用 QExactive高分辨质谱仪对样品成分进行分析鉴定。ACQUITY UPLC BEH C18色谱柱(100.0 mm×2.1 mm,1.7 μm)。流动相A为体积分数0.1%的甲酸,B为体积分数0.1%的甲酸乙腈。梯度洗脱:0.01 min,5% B;1.5 min,5% B;3 min,30% B;7 min,60% B;9 min,90%B;11 min,100%B;12 min,100%B;15 min,5%B,百分数皆为体积分数。自动进样器温度为4 ℃,柱温为45 ℃,体积流量为0.35 mL·min−1,进样量为10 μL。
-
采用正负离子扫描模式采集样品质谱信号,扫描的选择范围为60~900 m/z,正负离子电离均使用电喷雾离子源(ESI),正离子模式电压为3.5 kV,负离子模式电压为3.1 kV,毛细管温度为320 ℃。
-
采用UNIFI 1.8.1软件采集原始数据后经Progenesis QI v2.3 软件(Nonlinear Dynamics,Newcastle,英国)预处理。采用The Human Metabolome Database (HMDB)、Lipidmaps v2.3和METLIN数据库以及自建库对含有二级质谱数据的峰进行物质鉴定和注释。删除组内缺失值均>40%的离子峰,得到代谢物数据矩阵。采用SIMCA-P 14.1 软件进行多元统计分析,统计分析采用主成分分析(PCA)、偏最小二乘判别(PLS-DA)、正交偏最小二乘判别(OPLS-DA)[第1主成分的变量权重值(VIP>1)]和Student’s t检验(P<0.05)和变异倍数分析相结合的办法,来筛选差异代谢物。利用R软件对VIP前50的差异代谢物的表达量进行层次聚类分析,绘制火山图对P和fold change值进行可视化,以筛选显著差异代谢物。利用京都基因和基因组百科全书(KEGG) ID对显著性差异表达代谢物(P<0.05)进行通路富集分析,用R软件绘制气泡图以可视化显著富集的代谢通路[32]。采用Canoco 5.0对不同退化高寒草甸土壤样本标志代谢物与环境因子进行冗余分析(RDA),以揭示二者间的相关性。
-
由表2可见:三江源地区不同退化程度下高寒草甸土壤pH为6.39~7.34,随着高寒草甸退化程度增加而升高,MDM的土壤pH显著高于其他草甸(P<0.05)。相反,随着草甸退化程度的增加,SMC和SOM质量分数显著下降(P<0.05),其中SDM最低。TN、TP、NN和AP质量分数呈急剧下降后上升趋势,MDM中最低(P<0.05)。AK质量分数呈先升高后下降的趋势,在LDM中最高。同时,不同退化程度草甸下SC活性无显著差异,在MDM中活性最低。UE活性随着草甸退化的增加呈先升高后降低的趋势,在LDM中最高。NP活性呈先下降后升高的趋势,在MDM中活性最低。
表 2 样地土壤理化性质
Table 2. Soil physical and chemical characteristics of different degraded alpine meadows
样地 pH 土壤含水量
(SMC)/%有机质(OM)/
(g·kg−1)全氮(TN) /
(g·kg−1)全磷(TP) /
(g·kg−1)硝态氮(NN) /
(mg·kg−1)NDM 6.39±0.08 d 45.15±1.28 a 177.70±6.58 a 11.75±0.20 a 1.23±0.02 a 8.49±1.36 a LDM 6.84±0.03 c 41.37±2.13 a 154.22±0.24 b 9.79±0.03 b 0.92±0.02 b 6.90±0.44 b MDM 7.68±0.02 a 21.32±1.12 b 93.55±0.68 c 4.97±0.08 d 0.74±0.02 c 3.90±0.47 d SDM 7.34±0.03 b 21.16±0.98 b 84.07±1.52 c 5.46±0.10 c 0.81±0.02 b 5.01±0.31 c 样地 速效磷(AP)/
(mg·kg−1)速效钾(AK)/
(mg·kg−1)蔗糖酶(SC)/
(mg·g−1·d−1)脲酶(UE)/
(µg·g−1·d−1)中性磷酸酶(NP)/
(mg·g−1·d−1)NDM 0.57±0.03 a 168.91±3.86 b 130.53±2.27 a 1 142.39±147.17 c 3.42±0.06 a LDM 0.55±0.08 ab 208.37±3.72 a 128.27±2.38 a 1 758.94±60.26 a 3.22±0.99 ab MDM 0.31±0.01 ab 145.14±4.20 c 121.39±2.17 a 1 704.23±28.71 a 2.87±0.82 c SDM 0.36±0.06 b 138.37±6.83 d 126.19±2.13 a 1 414.22±155.38 b 3.16±0.43 b 说明:同列不同字母表示差异显著(P<0.05)。 -
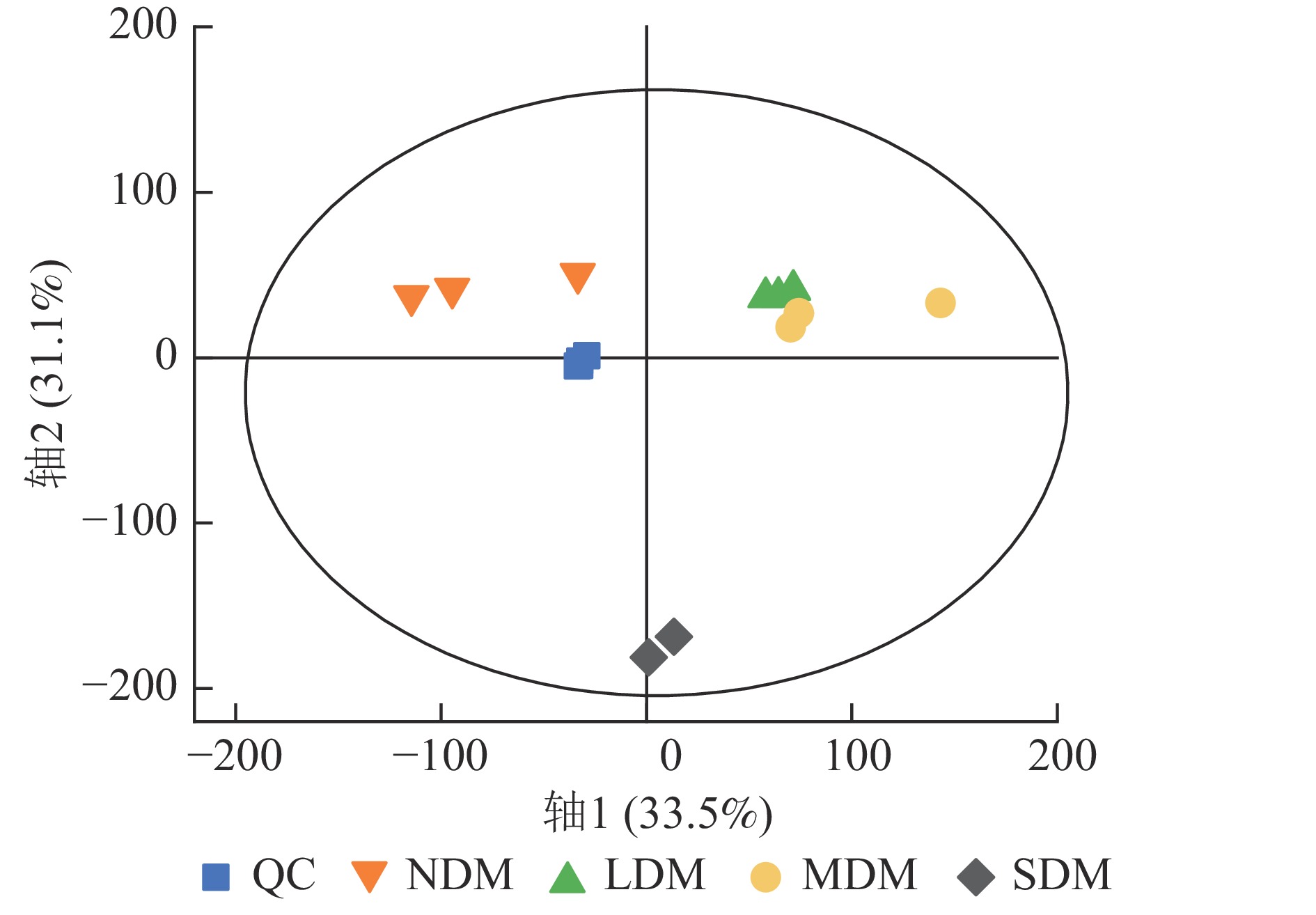
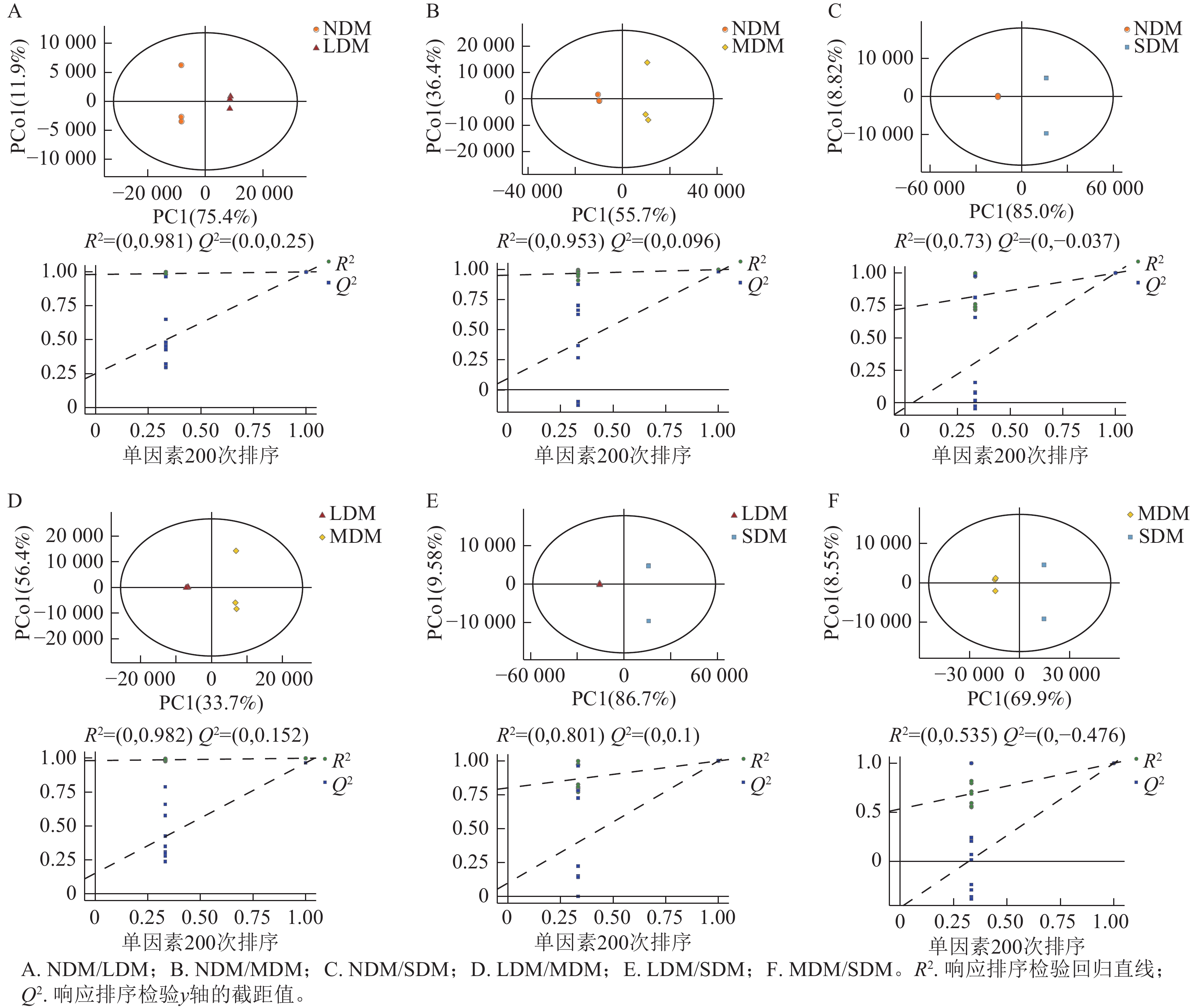
根据不同退化程度高寒草甸土壤样本的PCA结果可知:QC样本相对聚集(图1),表明本研究获得的代谢组学数据变异性小,数据具有较强的稳定性和重现性。同时,不同样本间明显分离,表明主成分分析对不同样本区分效果较好,不同退化高寒草甸土壤代谢物组成及含量存在明显差异。根据OPLS-DA模型的得分散点图(图2),表明组内差异较小。模型参数中R2Y、R2X、Q2均接近于1,说明分析数据稳定可靠。根据OPLS-DA置换检验随机模型(次数n=200),可知模型未发生过拟合(图2),R2Y均大于0.99,Q2均大于0.96,说明该模型可以很好地区分样品。
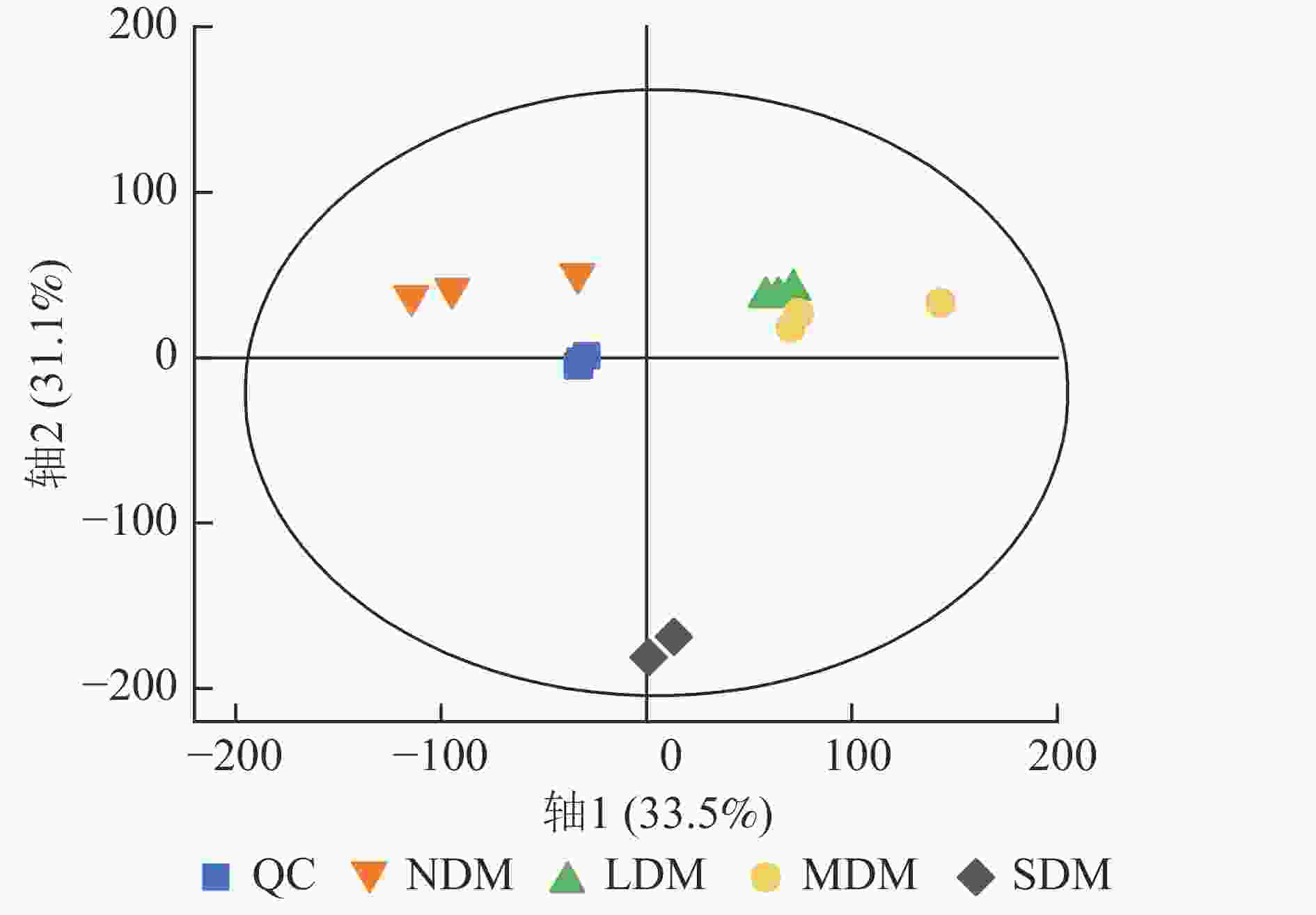
图 1 各组样本和QC样本PCA得分图
Figure 1. PCA scores plot of metabolites in all experimental samples and QC samples
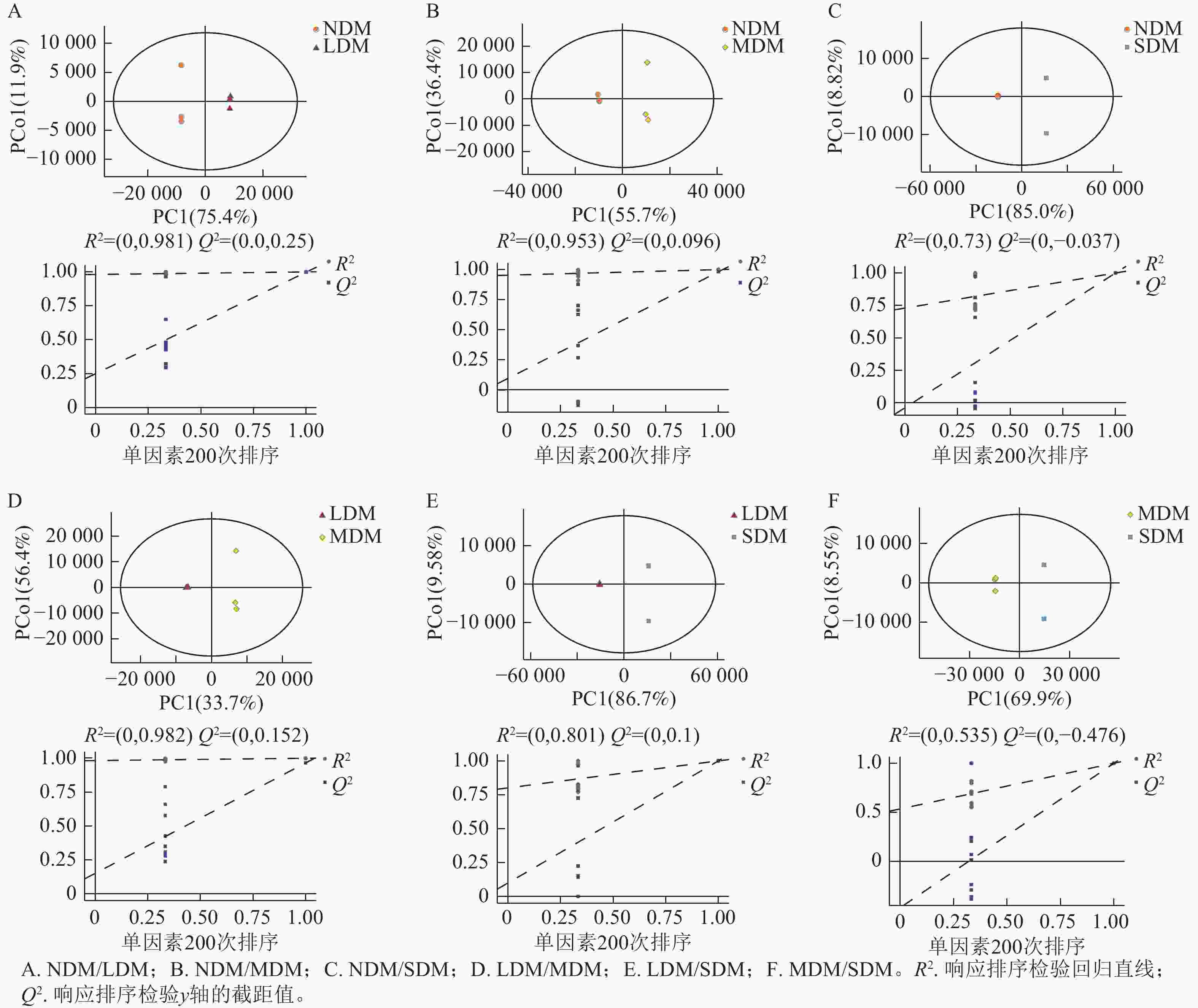
图 2 OPLS-DA得分图和置换检验
Figure 2. OPLS-DA score and displacement test
-
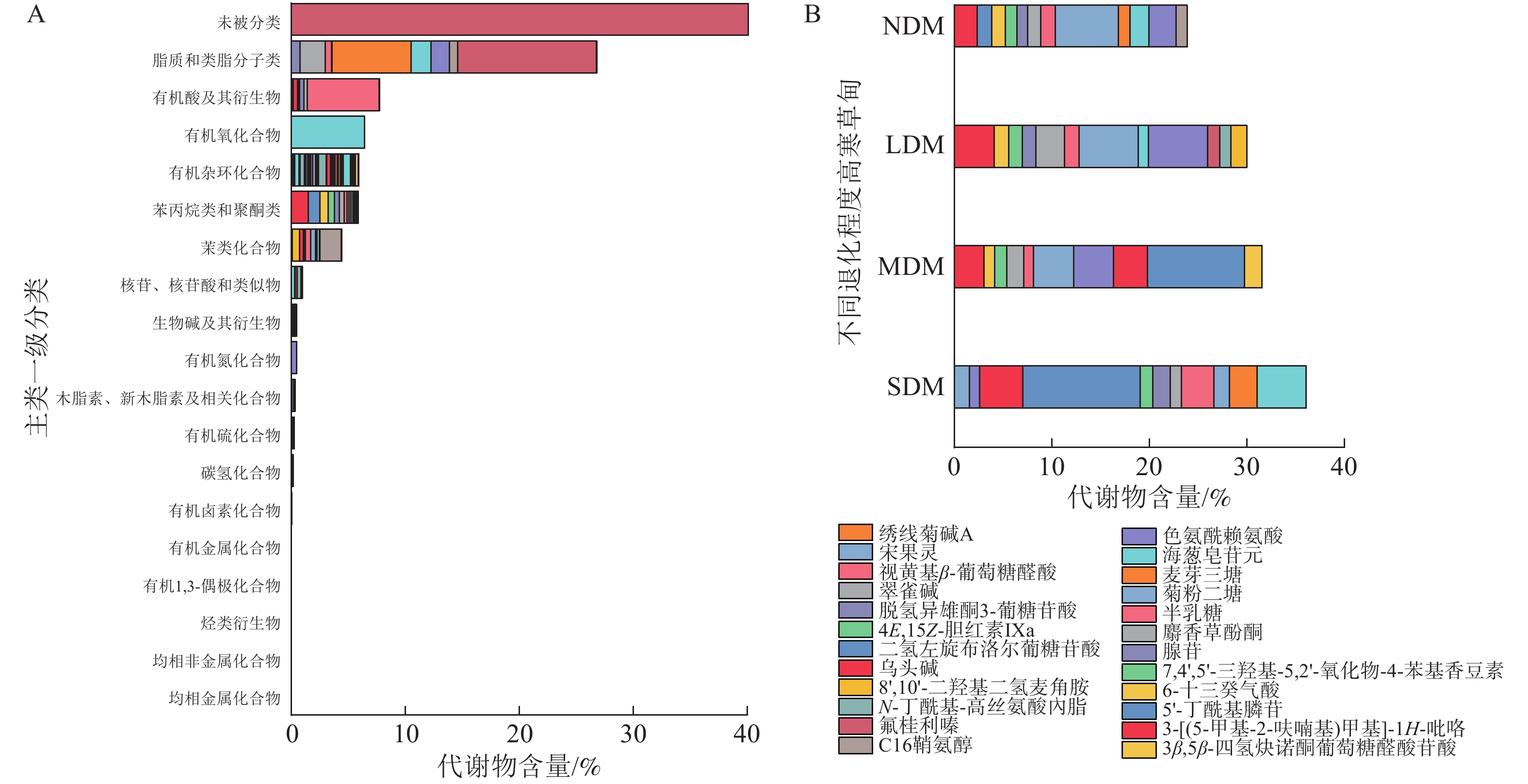
采用LC-MS测定,正离子模式下共鉴定出5 784种代谢物,负离子模式下2 667 种代谢物,根据 KEGG共鉴定出代谢物8 451种。代谢物主要包括脂质和类脂分子类(26.81%)、有机酸及其衍生物(7.73%)、有机氧化合物(6.44%)、有机杂环化合物(5.93%)、苯丙烷类和聚酮类(5.82%)和苯类化合物(4.45%),且代谢物中大部分为次生代谢物(图3A)。4个不同退化程度土壤中代谢物组成和含量存在显著差异(图3B)。NDM样地中代谢物共有 1 758 种,主要组成有菊粉二糖(6.46%)、海葱皂苷元(2.80%)和3-[(5-甲基-2-呋喃基)甲基]-1H-吡咯(2.37%);LDM样地中代谢物共有1 411种,主要组成有菊粉二糖(6.10%)、海葱皂苷元(6.03%)和3-[(5-甲基-2-呋喃基)甲基]-1H-吡咯(4.08%);MDM样地中代谢物共有1 327种,主要组成有乌头碱(9.95%)、海葱皂苷元(4.13%)和菊粉二糖(4.09%);SDM样地中代谢物共有1 224种,主要组成有乌头碱(12.06%)、绣线菊碱A (5.05%)和8',10'-二羟基二氢麦角胺(4.42%)。
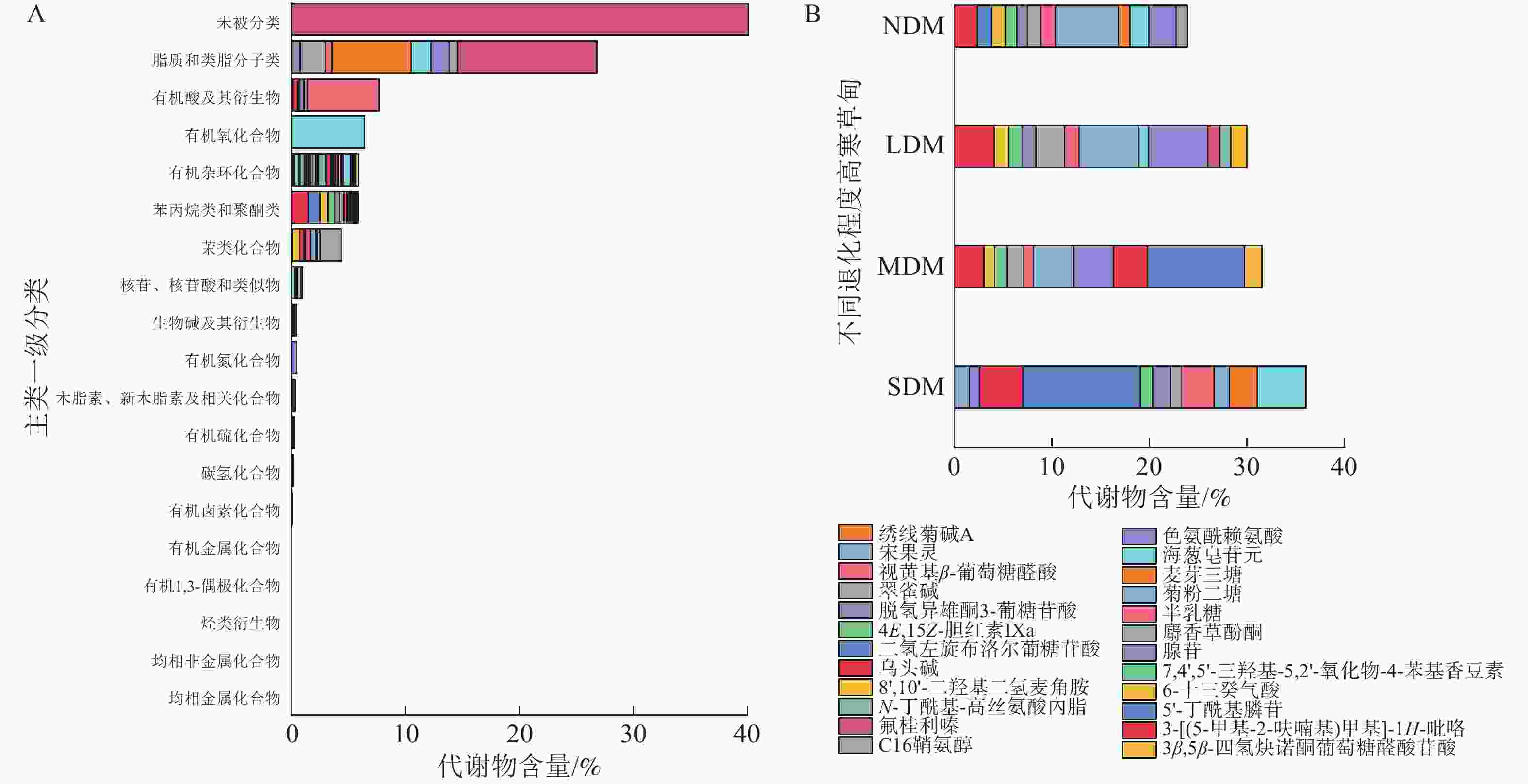
图 3 退化高寒草甸土壤样本中所有代谢物分类及占比(A)和不同程度退化高寒草甸土壤样本中质量分数大于1%的代谢物组成(B)
Figure 3. Classification and proportion of all metabolites in soil sample of the degraded alpine meadows (A) and classification of metabolites >1% in soil sample of different degraded alpine meadows (B)
-
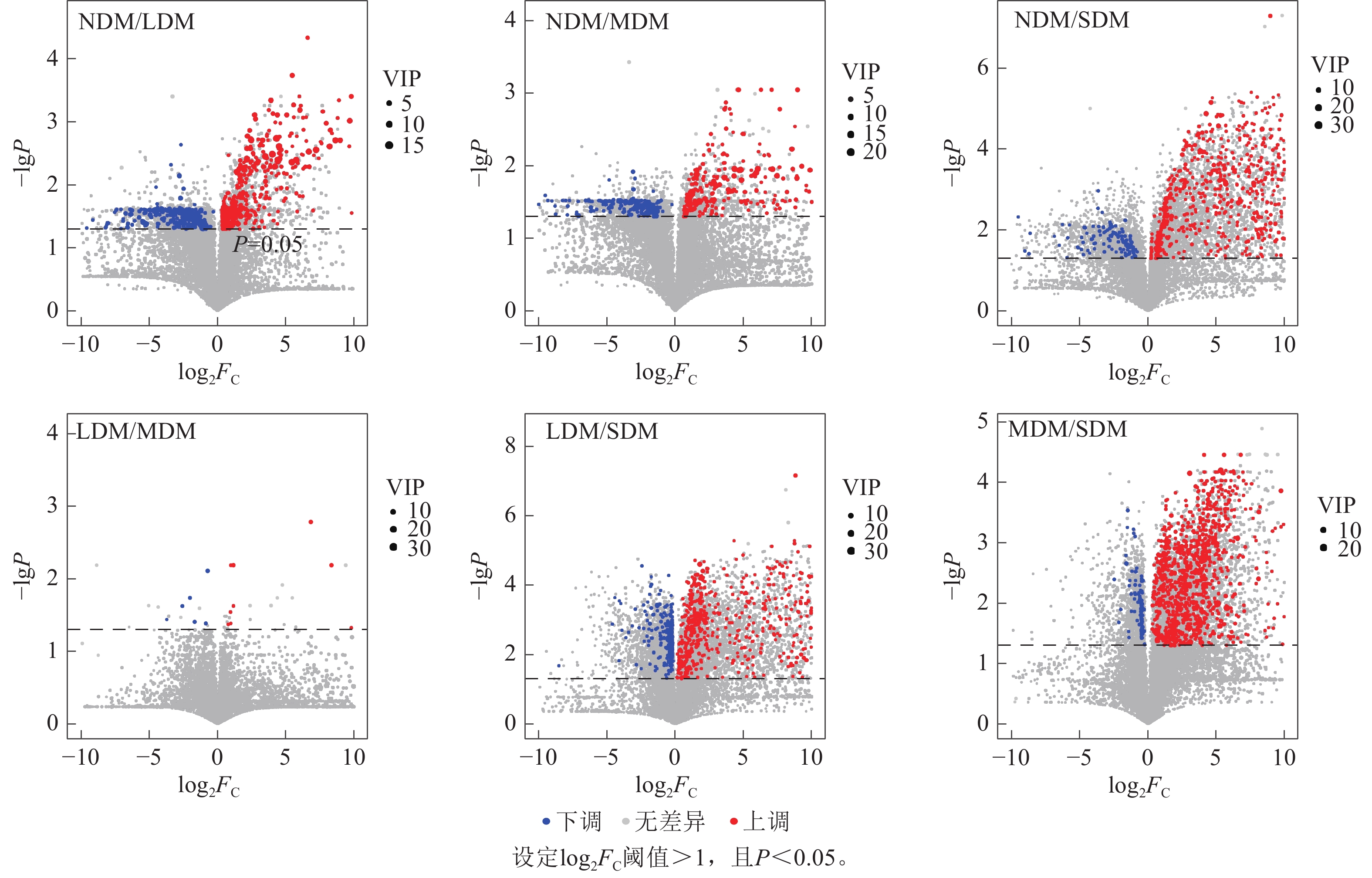
如图4所示:随三江源高寒草甸退化程度增加,各比较组间上调差异代谢物数量高于下调差异代谢物,尤其是SDM比NDM、LDM和MDM的土壤中差异代谢物种类多,其中,SDM和NDM中显著上调的代谢物数量比下调代谢物数量高450种,SDM和LDM中显著上调的代谢物数量比下调代谢物数量高429种,SDM和MDM显著上调的代谢物数量比下调代谢物数量高381种(
https://zlxb.zafu.edu.cn/fileZJNLDXXB/journal/article/file/03cdd59f-3922-4975-990a-bc367563243e.pdf )。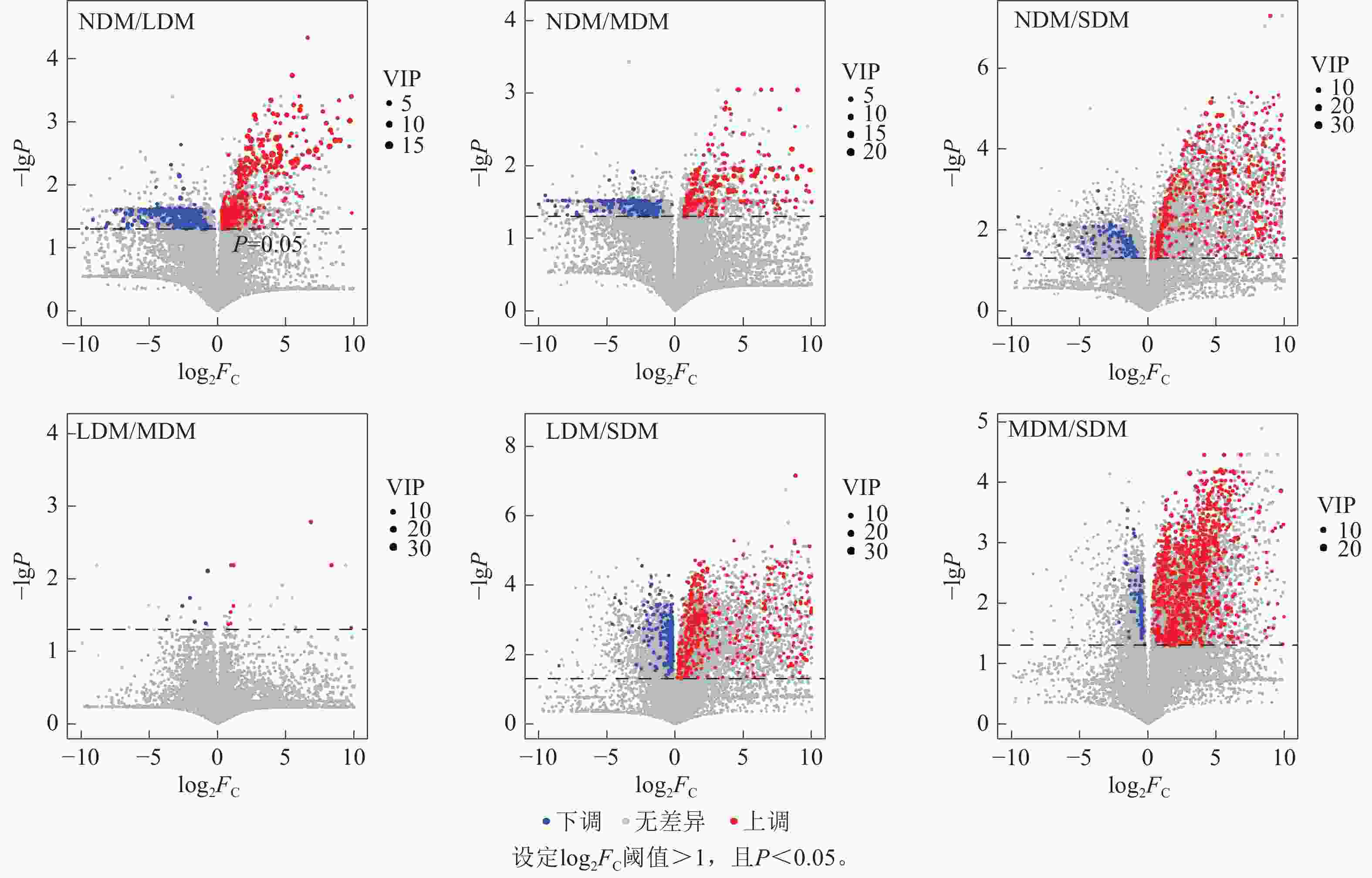
图 4 不同程度退化高寒草甸土壤差异代谢物火山图
Figure 4. Volcanic plots of differential metabolites in soil samples of 4 different degraded alpine meadows
-
为进一步确定不同退化高寒草甸土壤代谢物的差异,分别筛选了不同样本比较组表达量前20的差异代谢物(
https://zlxb.zafu.edu.cn/fileZJNLDXXB/journal/article/file/03cdd59f-3922-4975-990a-bc367563243e.pdf )。NDM中TOP20的代谢物中正离子模式下有14种(10种显著上调,4种显著下调),负离子模式下有6种(2种显著上调,4种显著下调),这些代谢物多属于甾醇脂类、吡咯类、嘌呤核苷类、羧酸及其衍生物和脂肪酰基类化合物;LDM中TOP20的代谢物中正离子模式下有16种(14种显著上调,2种显著下调),负离子模式下有4种(3种显著上调,1种显著下调),这些代谢物多属于羧酸及其衍生物、苯及其取代衍生物、吲哚及其衍生物;MDM中TOP20的代谢物均在正离子模式下,而且18种显著上调,2种显著下调,这些代谢物多属于羧酸及其衍生物、脂肪酰基、类固醇和类固醇衍生物、吲哚及其衍生物和甘油磷脂类化合物;SDM中TOP20的代谢物均在正离子模式下且均显著上调,这些代谢物多属于类固醇和类固醇衍生物、吡咯、丙烯醇脂质、羧酸及其衍生物和甾醇脂质类化合物。同时,对4种不同退化高寒草甸土壤样本6个比较组中TOP20的代谢物表达量变化(
https://zlxb.zafu.edu.cn/fileZJNLDXXB/journal/article/file/03cdd59f-3922-4975-990a-bc367563243e.pdf )分析表明:N-乙酰基-L-谷氨酸酯、3-[(5-甲基-2-呋喃基)甲基]-1H-吡咯、N-棕榈酰色胺、11-β-羟基雄甾酮-3-葡萄糖醛酸苷、水苏糖、C1鞘氨醇、4-丙酰基-HT2毒素和乌头碱的差异倍数随着退化程度的增加逐渐变大,可作为区分不同退化程度的标识物。 -
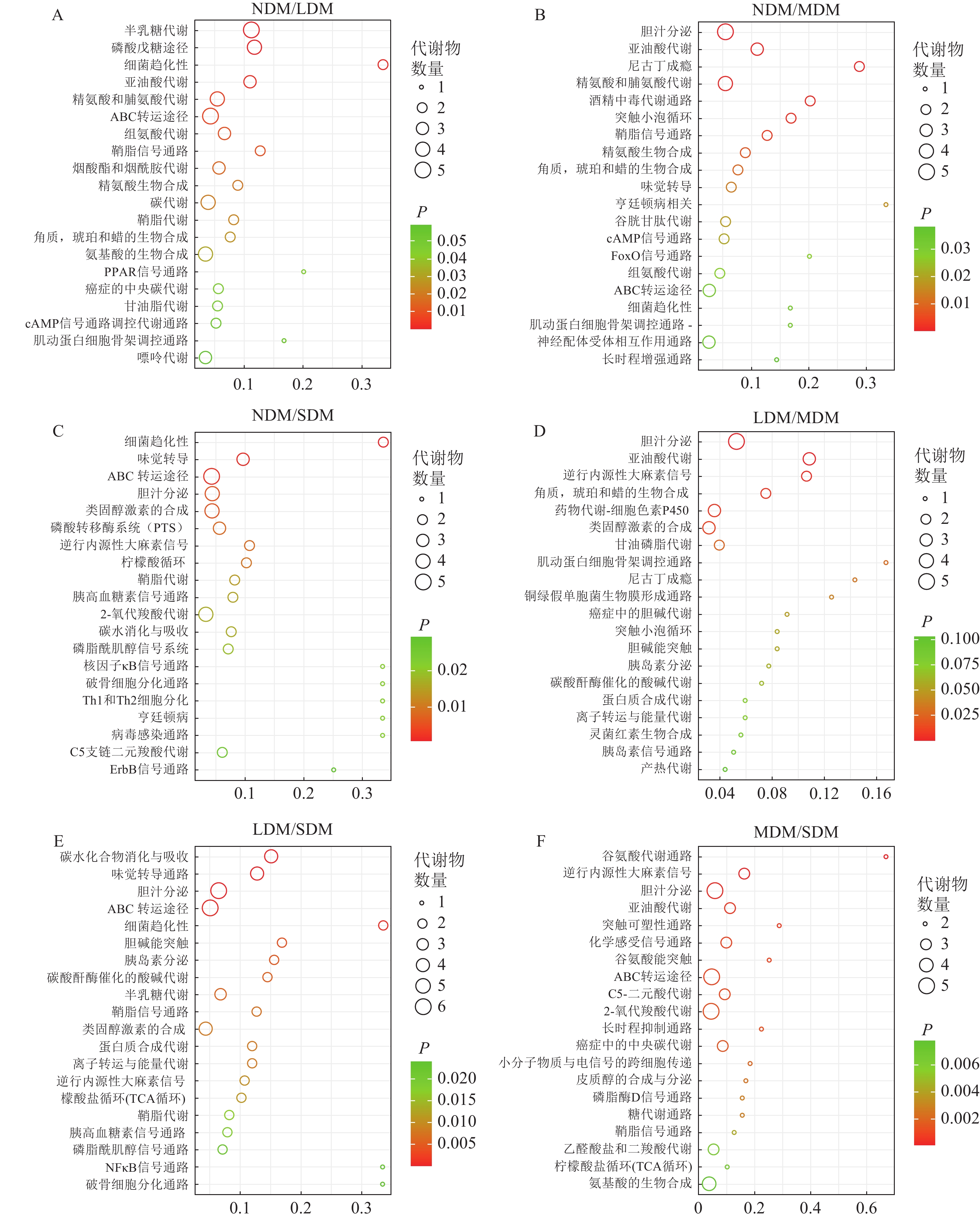
基于KEGG数据库的不同退化高寒草甸土壤样本差异代谢物通路富集分析,亚油酸代谢(linoleic acid metabolism)、ABC 转运途径(ABC transporters)、磷脂酶D信号(phospholipase D signaling pathway) 通路和鞘脂信号通路(sphingolipid signaling pathway)在6个比较组中均极显著富集(P<0.01),为Top20关键代谢通路(图5)。其中,NDM与LDM、MDM、SDM组间分别富集差异代谢通路17、29和43条,对应代谢物数目56、33和44 种,其中磷酸戊糖途径(pentose phosphate pathway) (图5A)、鞘脂信号通路代谢通路(图5B)、ABC 转运途径(图5C)均极显著富集(P<0.01);LDM 与MDM、SDM组间分别富集10和49条通路,对应代谢物数目28和49种,亚油酸代谢(图5D)和ABC 转运途径(图5E)极显著富集(P<0.01);MDM 与SDM 组中富集72条差异代谢通路,37 种代谢物,鞘脂信号通路极显著富集(图5F) (P<0.01)。
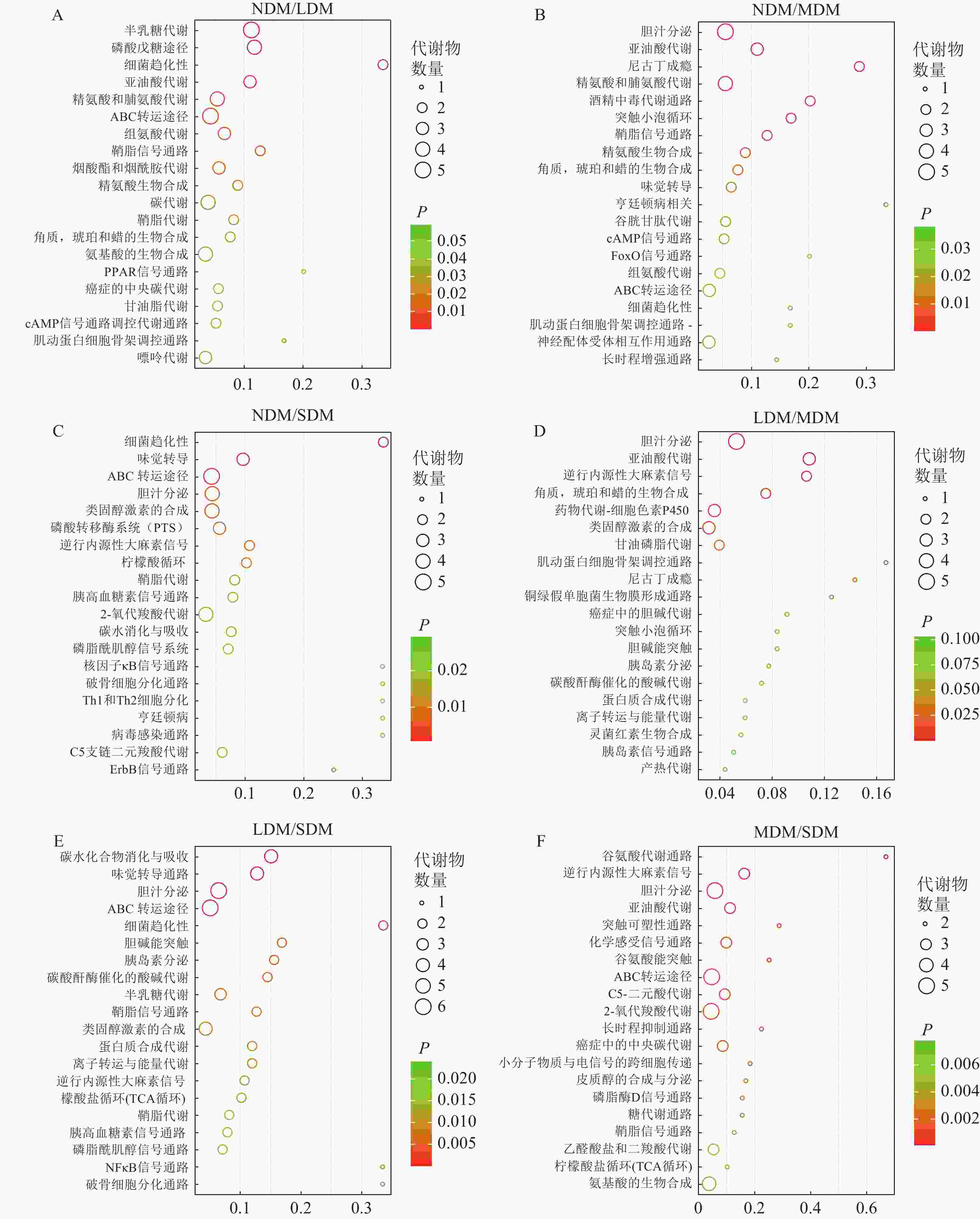
图 5 土壤样本差异代谢通路富集Top20气泡图
Figure 5. Bubble chart of KEGG enrichment analysis of the top 20 different metabolites in the soil samples
-
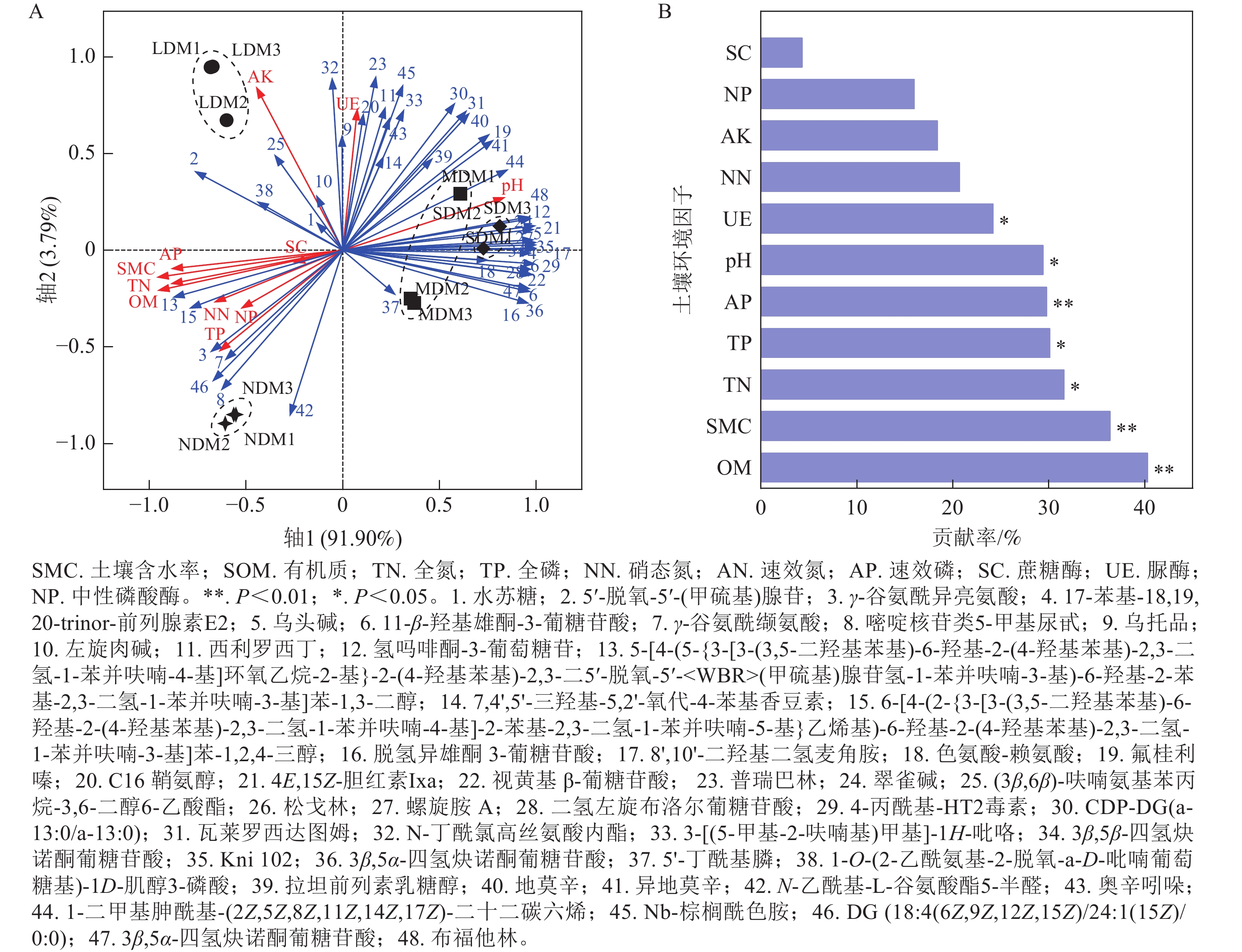
采用RDA分析解析了4种不同退化程度高寒草甸土壤样本前20差异代谢物与环境因子相关性。如图6A所示:第1、第2排序轴的解释量分别为91.90%和3.79%。由图6B可知:对土壤差异代谢物变化影响显著的土壤环境因子指标为SOM (P=0.002)、SMC (P=0.004)、TN (P=0.01)、TP (P=0.014)、AP (P=0.006)、pH (P=0.014)和UE (P=0.03)。NDM中大部分差异代谢物与SOM、SMC、TN、TP、AP呈显著正相关(P<0.05),与pH和UE呈显著负相关(P<0.05); LDM中,脂质和类脂质分子32对AK和UE呈显著正相关(P<0.05);在MDM和SDM中,脂质和类脂质分子4、16、29和44,有机酸及其衍生物23和41以及生物碱及其衍生物12与pH和UE具有显著正相关性(P<0.05),与SOM、SMC、TN呈显著负相关(P<0.05)。结果表明高寒草甸退化可能导致土壤代谢物组成受到SOM、SMC、TN、TP、AP、pH和UE的显著影响,SOM和SMC对代谢物的影响最为显著。
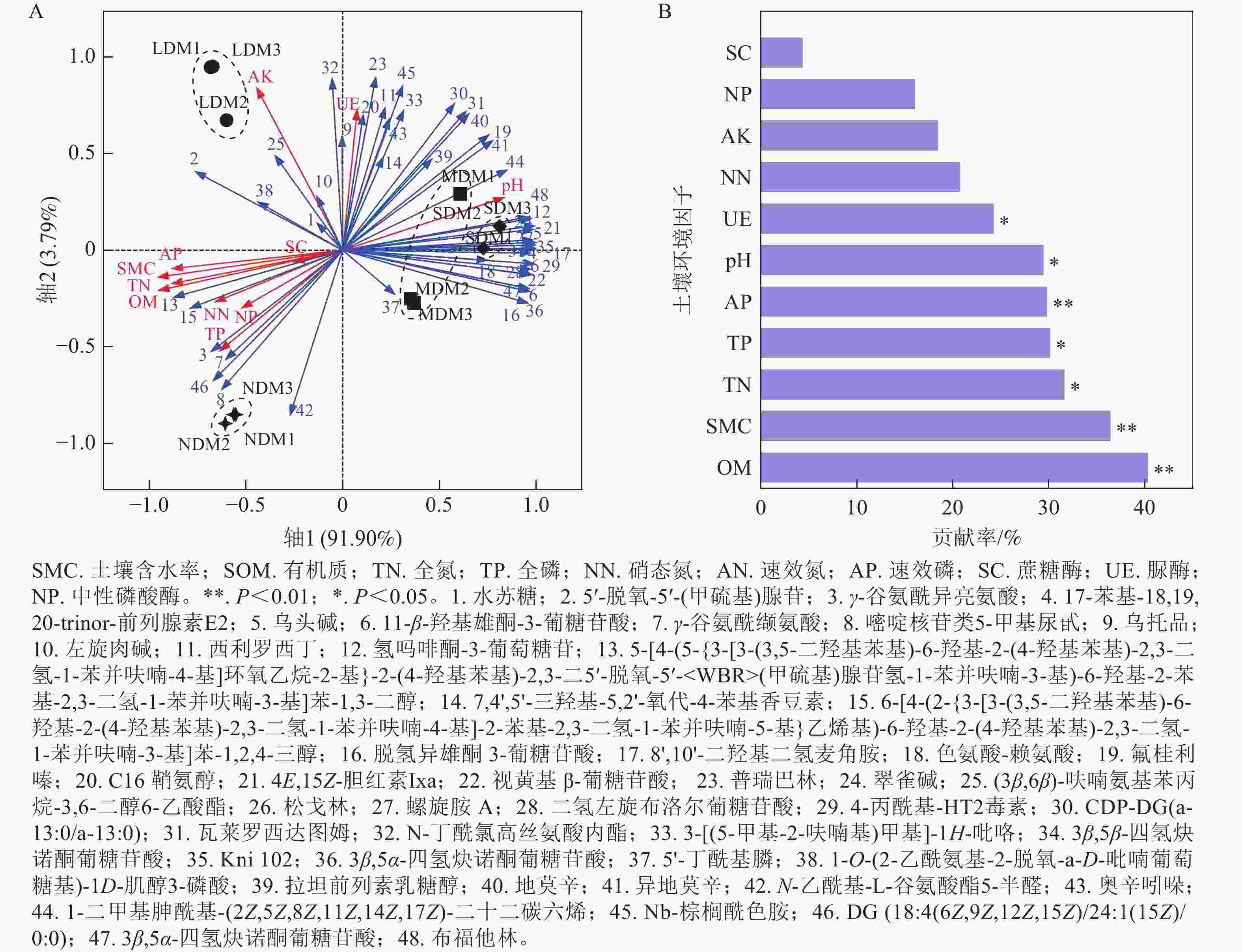
图 6 4种不同程度退化高寒草甸土壤差异代谢物与土壤环境因子的 RDA分析
Figure 6. RDA analysis of differential metabolites in soil samples of 4 different degraded alpine meadows and environmental factors
-
土壤代谢物成分高度复杂且具有较高的生物利用度,是土壤-植物-微生物生态系统间物质交换和能量流动的基础,在调控微生物营养网络、调节土壤化学、土壤微生物基因表达及植物生长发育等方面发挥着改进作用[33−34]。本研究发现:三江源退化高寒草甸土壤代谢产物丰富,以脂质和类脂分子为主。这2类物质持续累积可抑制植物生长信号通路并破坏土壤团聚体结构[35]。不同退化样地间代谢物组成差异显著,中度和重度退化草甸中苯丙烷类、聚酮类等具有化感潜力的化合物特异性富集,可能在退化高寒草甸群落种间竞争和演替中的具有重要作用[36]。已有研究表明:退化通过改变植物根系分泌物[10, 37]和微生物群落[17]结构的共同驱动土壤代谢物变化。高寒草甸主要土壤代谢物为菊粉二糖和海葱皂苷元,是典型的应激代谢物[27],且随退化加剧而减少,其衰减扰动了土壤主要养分循环和能量流动[38]。由此可见,退化高寒草甸土壤代谢物的组成和分布特征与植被群落及土壤微生态环境密切相关[39]。
本研究发现:不同退化程度高寒草甸土壤代谢物显著分离,其中重度退化草甸与其他组间的显著差异代谢物数量较多,表明代谢调控网络更为复杂。各比较组中上调差异代谢物的优势富集使其可作为退化的关键标志物。本研究鉴定出差异标志性代谢物主要涉及的羧酸及其衍生物、脂肪酰基和鞘脂类等化合物不仅直接参与碳磷氮循环[40],还为土壤微生物和植物提供碳源[41]。随着退化的加剧,羧酸及其衍生物、吡咯类化合物及吲哚及其衍生物普遍呈上调趋势,这些代谢物累积可能是土壤环境恶化[40]、植被根系分泌释放[37]、微生物的选择性利用[42]共同作用的结果。在中度和重度退化草甸中,有机氧化合物(水苏糖)和鞘脂类化合物(C16鞘氨醇)显著上调。水苏糖作为一种能量储存和胁迫响应分子有助于维持植物的正常生理功能[43];C16鞘氨醇则通过诱导系统抗性和次生代谢产物拮抗病原菌发挥生防作用[44]。因此,退化过程中特定化合物释放及协同作用共同增强了植物的抗逆能力。此外,重度退化草甸中戊二烯醇脂质化合物(4-丙酰基-HT2毒素)和有机氮化合物(乌头碱)也显著累积,前者为镰刀菌属Fusarium的主要代谢产物,可能与土壤中病原菌增加有关[45];而乌头碱属于高寒草甸毒杂草的潜在化感物质,可显著抑制优质牧草生长[46],进而驱动植被演替。由此可见,高寒草甸不同退化程度土壤样本中标志性代谢物特征性变化可为高寒草甸退化早期诊断提供参考依据。
在不同退化程度的高寒草甸土壤中,差异代谢物显著富集于亚油酸代谢、ABC 转运途径、磷脂酶D信号通路和鞘脂信号通路,其中磷脂酶D信号通路的富集程度最高。该通路中显著上调的肌醇1,4,5-三磷酸和L-谷氨酸促进了磷脂酶D物质的积累,在糖代谢和信号转导中扮演着重要角色,通过提高有机物质水解来响应退化[47]。亚油酸代谢通路的途径参与免疫调节、细胞信号传导等生理功能,同时在土壤有机碳循环起着关键作用[48]。鞘脂信号通路是激活鞘质醇激酶产生鞘烷基-1-磷酸(S1P),在低温胁迫下增加细胞膜的流动性和稳定性,从而提升植物的抗逆性[49]。ABC转运途径则参与无机离子、聚糖与胆固醇等多种代谢产物的跨膜转运,对土壤有机物的吸收、分解与转化具有关键影响[50−51]。因此,差异代谢物的富集通路及功能分析为揭示高寒草甸退化过程中土壤环境变化与功能响应奠定了基础。
退化高寒草甸土壤代谢产物的变化与土壤环境因子密切相关[52]。本研究发现:随着退化加剧,SOM、SMC、TN、TP、AP和UE活性降低,而pH升高。这一变化可归因于植物群落盖度、地上生物量和土壤含水量导致土壤中凋落物归还量下降,进而降低了土壤碳氮磷的循环效率[8];同时,土壤代谢物如有机酸和胺类代谢物通过络合金属离子或调节pH,进一步影响土壤养分有效性和酶活性。RDA分析结果表明:未退化草甸中γ-谷氨酰异亮氨酸和γ-谷氨酰缬氨酸等羧酸及其衍生物和2-芳基苯并呋喃类黄酮类化合物与SOM、SMC、TN、TP呈显著正相关。这些代谢物参与根际碳循环和难溶性磷活化,为植物提供碳源和速效磷[53−54],同时,无机养分富集导致初级和次级代谢以及资源流动和土壤功能的变化[44]。中度和重度退化草甸中类固醇类、脂肪酰基、生物碱和多种羧酸类化合物与pH和UE呈极显著正相关,研究表明退化导致土壤养分损耗,这些代谢物激活了碳、氮、磷 循环相关的根际过程(碳沉积、微生物碳转化、氮素矿化等),从而调控土壤微生态环境[44]。同时,pH增加导致代谢物迁移[55],SOM是影响可溶性代谢物含量的主要因子[23],共同导致重度退化草甸中代谢物种类和表达量显著上调,其中羧酸及其衍生物受土壤理化性质和酶活性影响最为显著,可作为退化过程中最敏感的“先驱代谢物”。
-
三江源不同退化程度高寒草甸土壤代谢物以脂质、有机酸及其衍生物、有机氧化合物、苯丙烷类和聚酮类等次生代谢物为主,其种类和含量因退化程度不同呈现显著差异。其中,羧酸及其衍生物、脂肪酰基、吡咯、吲哚及其衍生物、有机氮化合物、有机氧化合物、戊二烯醇脂质和鞘脂类化合物为差异标志性代谢物。重度退化草甸与未退化、轻度退化和中度退化草甸的显著差异代谢物种类和数量均高于其他比较组,说明重度退化草甸中代谢通路更为复杂。而且,各比较组中上调差异代谢物普遍高于下调代谢物,揭示特定代谢物的正向富集可作为退化的关键标志物。差异代谢物主要富集于亚油酸代谢、ABC 转运途径、磷脂酶D信号通路和鞘脂信号通路等关键途径,这些通路与植被群落和土壤微生态环境的变化密切相关。SOM、SMC、TN、TP、AP、pH和UE等土壤环境因子与土壤代谢物具有显著相关性,推测是驱动不同退化高寒草甸土壤差异代谢物变化的关键环境因子。
全文HTML
1.1. 研究区概况
1.2. 样品采集
1.3. 土壤理化性质分析及土壤酶活性测定
1.4. 代谢组样品前处理
1.5. 色谱条件
1.6. 质谱条件
1.7. 数据处理
2.1. 不同退化高寒草甸土壤理化性质和酶活性变化
2.2. 不同退化高寒草甸土壤样品PCA和OPLS-DA结果
2.3. 不同退化高寒草甸土壤样本代谢物组分分析
2.4. 不同退化高寒草甸土壤样本差异代谢物的火山图分析
2.5. 不同退化高寒草甸土壤样本标志代谢物筛选
2.6. 不同退化程度高寒草甸土壤样本差异代谢物通路富集分析
2.7. 不同退化程度高寒草甸土壤样本差异代谢物与理化性质及酶活性相关性分析
 2025-0227-附件材料.pdf
2025-0227-附件材料.pdf
|

|
