






-
香榧Torreya grandis ‘Merrillii’是中国特有的珍贵多用途经济干果树种,其发源地和主产区在浙江省会稽山区[1]。香榧营养丰富,风味独特,为干果中的珍品[2]。由于资源少,产品供不应求,成为目前经济价值和栽培效益最高的树种之一。矿质营养对植物生长发育、果实产量的形成和品质的改善有重大影响[3-4],营养与施肥作为生产上的调控手段,已成为改善果树产量和品质的主要途径之一[5-6]。叶片元素分析作为对植物体进行矿质营养研究的一种基本手段[7-8],也是一种较为成熟的果树营养诊断法[9],目前,对香榧叶片营养及林地土壤养分变化已有研究[10-12],但针对香榧种子速生期和充实期(5-9月),香榧叶片及种子中矿质元素动态变化的研究尚无报道,生产管理中的施肥环节也缺乏理论基础,普遍存在盲目施肥的现象。本研究通过测定香榧种子速生期和充实期内叶片及种子中矿物质元素的动态变化,了解香榧种子发育期内矿质元素变化情况,为香榧科学施肥提供理论依据。
-
试验于2012年在香榧主产区浙江省嵊州市谷来镇进行。谷来镇位于浙江省嵊州市西北部,是典型的山区小镇,全镇拥有735 hm2省级香榧森林食品基地,地理坐标为29°35′N,120°49′ E,亚热带海洋性季风气候,年平均气温为16.4 ℃,7月最热(平均所温为28.6 ℃),1月最冷(平均温度为4.2 ℃),年平均降水量为1 446.8 mm,年平均日照时数1 988 h,无霜期为235 d。土壤类型为不同母质的红黄壤,缓坡地带多为次生黄壤。林地肥料以复合肥为主,配施农家肥。研究的香榧树龄(2+10)a(砧木为2年生榧树,嫁接后生长10 a),树体生长正常,并已进入结果期。
-
试验采用随机区组设计,5株·小区-1,重复3次。从2012年5月15日开始,1个月采样1次,至2012年12月20日止,共取8次样。随机摘取香榧树冠中部东、南、西、北方向1年生枝条1~2个,采枝条4~8个·株-1,剪下枝条上的叶片装袋带回实验室。种子采集在种子完全从苞片先端伸出(5月中下旬)开始,15 d采集1次,共采集7次。
样品带回实验室后依次用自来水和蒸馏水冲洗干净,再用去离子水冲洗,并用滤纸干燥。105 ℃杀青30 min,并于65 ℃烘干至恒量。将样品粉碎并混合均匀后备用。待测样品用硫酸-过氧化氢(H2SO4-H2O2)联合消煮液进行消煮。氮用凯氏定氮法测定;磷用钼蓝比色法测定;钾、钙、镁、铜、锰、锌等用原子吸收分光光度法测定[13]。所得数据用SAS 9.2进行简单相关分析和单因素方差分析,采用最小显著差数法(LSD法)进行平均数间的多重比较,Excel 2007作图。
-
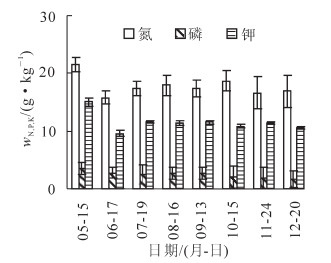
香榧生长期叶片中氮、磷、钾元素变化见图 1。3种元素均在5月份最高,然后呈现出不同的动态变化,其变化与香榧种子的生长发育(5-9月)及雌花芽分化(9月以后)的周期有关。氮元素在5-12月下降幅度为21.4%,而5-6月下降幅度为26.5%。这是因为6月香榧种子开始生长膨大,叶片中氮质量分数在种子膨大初期有所下降,供应给种子生长;7-8月种子充实后处于内部物质旺盛转化期,直至9月种子成熟及后续3个月,叶片中的氮质量分数相对稳定。磷元素从种子膨大初期至12月一直呈现下降趋势,总的下降幅度为52.4%,其中5-6月下降23.1%。种子充实及物质转化期至种子成熟,叶片中磷元素相对稳定,种子成熟后叶片中磷元素又呈现下降趋势,至12月降为最低点。叶片中钾在5-6月下降幅度为36.2%,6-7月叶片中的钾有所增加;7-12月,叶片钾有所波动,但相对稳定,变幅不大。从5月中旬至12月,叶片中的钾下降幅度为29.6%。9月种子采收后,氮元素变化不大,磷、钾元素均有下降,尤其是钾元素。在植物的幼嫩部分或旺盛生长部分,如芽、幼叶、根尖都含有大量钾,而雌球花冬季分化[2],叶片中流失的钾元素主要供应雌花芽的分化。

图 1 香榧叶片中氮、磷和钾元素的动态变化
Figure 1. Changes of N, P and K contents in leaves of Torreya grandis ‘Merrillii'
-
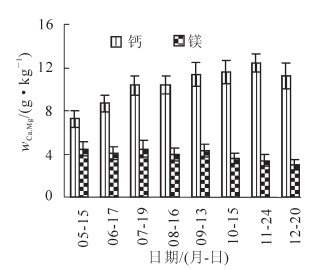
香榧生长期叶片中钙质量分数变化见图 2。因为钙是较难移动的元素[14],所以随着时间推移,钙质量分数不断升高。香榧叶片中的钙质量分数从2012年5-11月大致呈上升趋势,5月中旬叶片中钙处于整个年周期的最低水平;11-12月有小幅下降,此时是花芽分化期及根系旺盛生长期。叶片中镁质量分数在整个生长周期有小幅波动,总体呈下降趋势(图 2)。种子采收后是香榧的花芽分化期,9月种子采后1个月叶片中镁元素下降明显,随后缓慢下降,此时叶片中的镁有可能供应花芽分化。

图 2 香榧叶片中钙、锾元素的动态变化
Figure 2. Changes of Ca and Mg contents in leaves of Torreya grandis ‘Merrillii'
-
叶片中锰元素的变化(图 3),5月初较低(55.41 mg·kg-1),6月中旬开始快速上升,至11月中旬达到最大值(126.04 mg·kg-1),随后有所回落。叶片中铜元素的变化在种子体积速生期至种子发育成熟期总体上先呈现上升趋势,后又急剧下降,种子采摘后10-12月叶片中铜质量分数保持相对稳定。叶片中锌质量分数从5-6月,由26.02 mg·kg-1下降为17.15 mg·kg-1,降幅为34.1%,随后叶片中锌质量分数至10月中旬呈现上升趋势。

图 3 香榧叶片中锰、铜、锌元素的动态变化
Figure 3. Changes of Mn, Cu and Zn contents in leaves of Torreya grandis ‘Merrillii'
-
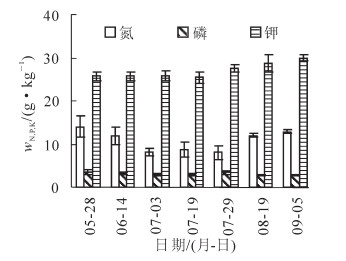
香榧种子大量元素中钾质量分数最高,其次为氮和磷(图 4)。钾元素在种子的速生期和充实期一直处于积累状态,而氮元素在速生期出现流失现象,后又不断积累,磷元素在种子的速生期和充实期始终处于一个较为平缓的过程,前后变化不明显。

图 4 香榧种子中氮、磷、钾元素的动态变化
Figure 4. Changes of N, P and K contents in Torreya grandis Torreya grandis ‘Merrillii' fruits
-
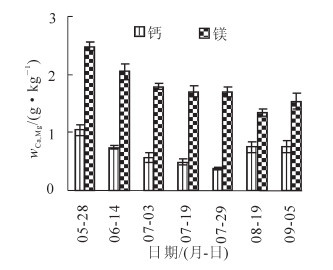
香榧种子中钙、镁质量分数在速生期和充实期变化趋势相似(图 5),5-7月均呈现下降趋势,钙元素在7月降为最低点,降幅为64.4%,8月后达到平稳状态;镁元素至8月降为最低点,降幅为37.9%,后又有所回升,在整个过程中镁质量分数一直高于钙元质量分数。

图 5 香榧种子中钙、锾元素的动态变化
Figure 5. Changes of Ga, Mg contents in Torreya grandis ‘Merrillii' fruits
-
香榧种子中锌质量分数显著高于铜、锰(图 6),锌质量分数在种子充实期呈现下降趋势,与叶片中锌元素的积累趋势,从一个方面说明种子中锌的主要积累时期在缓生期(第1年4月至第2年5月),锰质量分数在种子整个充实期呈现下降趋势,降幅为77.3%,铜质量分数则前后变化不明显。

图 6 香榧种子中锰、铜和锌元素的动态变化
Figure 6. Changes of Mn, Cu and Zn contents in Torreya grandis ‘Merrillii' fruits
-
在种子的速生期和充实期的5个月中,香榧叶片和种子中矿质元素之间的相关分析见表 1。相关分析结果显示:叶子和果实中的相同元素之间虽然存在一定的相关性,但并不显著,而叶子中磷元素与果实中钙、镁、锰存在着显著的相关性(P < 0.05)。叶子中铜元素与果实中锌、锰也存在着显著的相关性(P < 0.05)。
表 1 香榧叶片和种子中矿质元素之间的相关系数
Table 1. Correlation of mineral elements between leaves and fruits in Torreya grandis ‘Merrillii'
种子矿质元素 叶片矿质元素 氮 憐 钾 钙 镁 锌 铜 锰 N 0.521 0.729 0.456 -0.422 -0.127 0.018 -0.721 -0.272 P 0.546 0.802 0.533 -0.960** 0.378 -0.547 -0.875 -0.922* K -0.148 -0.292 -0.159 0.704 -0.313 0.644 0.383 0.787 Ca 0.713 0.906* 0.646 -0.678 0.012 -0.085 -0.892* -0.523 Mg 0.553 0.835 0.551 -0.924* 0.434 -0.493 -0.910* -0.866 Zn 0.656 0.924* 0.655 -0.812 0.446 -0.264 -0.965** -0.686 Cu 0.563 0.769 0.480 -0.528 -0.195 -0.068 -0.763 -0.391 Mn 0.662 0.924* 0.645 -0.933* 0.379 -0.403 -0.974** -0.840 说明:**表示相关系数在0:01水平极显著相关;*表示相关系数在0.05水平显著相关。 -
从本研究结果来看:氮、磷2种元素,无论叶子还是种子在5-6月均呈现下降趋势,且叶子中下降更为明显,这与张春苗[10]研究结果一致。叶片中氮、磷2种元素在果实膨大期下降后又呈现上升趋势。而钾元素,叶子中下降幅度较大,但在种子中呈现上升趋势,说明在种子速生期开始之前,3-4月应该相应追施氮、磷、钾肥料,特别是氮、磷2种肥料。
随着种子体积膨大的结束,果壳的逐渐形成,种子进入内部充实期,7月中旬后种子中钾元素增加速率变大,而叶子中钾元素继续呈现下降趋势,说明在7月应适当补充钾肥,以保证叶子和种子对钾肥的需求。
杨光等[15]报道:木瓜Chaenomeles sinensis叶片中钙、镁质量分数变化很相似,即都在波动中呈上升趋势。解红恩[16]研究表明:山核桃Carya cathayensis果叶中钙、镁质量分数变化规律相似。本研究表明:香榧叶片和种子中镁质量分数均呈现下降趋势,说明在种子速生期和充实期无需施入镁肥,但按照“归还学说”,应考虑在种子采摘后,增加镁肥,保证幼果在缓生期对镁肥的需求。与叶片中钙质量分数一直呈上升的趋势相反,种子中钙质量分数在8月中旬前一直呈下降趋势,后期略有上升,说明在种子进入速生期,应该适当增施钙肥。
对于锰元素,仅香榧林地土壤研究有所报道,香榧林地不缺锰[17],其他尚未见报道,而且叶片中锰质量分数较高,铜、锌质量分数较低。但在种子中锌质量分数较高,铜、锰质量分数较低,说明叶片中锌、锰质量分数与种子中锌、锰质量分数无直接相关性,在进行叶营养诊断时应加以考虑。
相关分析的结果表明:香榧叶片中磷质量分数与种子中钙、锌和锰质量分数均存在显著的正相关关系;叶片中钙质量分数与种子中磷质量分数存在极显著的负相关关系,与镁、锰存在显著的负相关关系;叶片中的铜质量分数与种子中锌、锰质量分数呈极显著的负相关关系,与钙和镁存在着显著的负相关关系;同样叶片中锰元素质量分数与种子中磷元素质量分数存在着显著的负相关关系。鉴于香榧叶片与种子中不同元素间的协同或者拮抗作用,特别是磷肥与微量元素锌、锰之间存在显著的正相关,在施肥时应该注意各种矿质元素用量的协调,保证香榧树体营养均衡,植株健康,提高产量的同时,避免香榧种子中重金属超标。由此推断:种子进入速生期前应适当增施氮肥、磷肥和钙肥,7月中旬适当补施钾肥,种子采收后补施镁肥,并重视磷肥和微肥的配合施用,对香榧产量和质量会产生积极影响。
Mineral elements in leaves and seeds of Torreya grandis ‘Merrillii’ during seed development
-
摘要: 为了探讨香榧Torreya grandis ‘Merrillii’生长期叶片营养状况与果实营养状况的关系, 对香榧在种子速生期和充实期叶片和种子中8种矿质元素进行动态分析。结果表明:①香榧叶片中氮、磷、钾在种子膨大初期下降幅度较大, 分别为26.5%, 23.1%和36.2%;种子中氮、磷的下降幅度分别为15.4%和8.3%, 钾元素呈现上升趋势。②叶片中钙、锰、铜在整个实验过程中均呈现上升趋势, 香榧叶片中镁元素一直呈现下降趋势; 在种子中这4种元素呈现先下降后上升的趋势。③叶片中磷质量分数与种子中钙、锌和锰质量分数均存在显著的正相关关系(P < 0.05), 而叶片中钙质量分数与种子中磷质量分数存在极显著的负相关关系(P < 0.01)。由此推断:种子进入速生期前应适当增施氮肥、磷肥和钙肥, 7月中旬适当补施钾肥, 种子采收后补施镁肥, 并重视磷肥和微肥的配合施用, 对香榧产量和质量会产生积极影响。Abstract: To explore the relationship between leaf and seed nutrients of Torreya grandis ‘Merrillii', eight mineral elements (N, P, K, Ca, Mn, Cu, Mg, and Zn) were quantified. Then, analysis of their dynamic changes during fast growth and seed filling was conducted using a correlation analysis. Results at the early stage when seeds enlarged showed 1) decreases in the leaves for N (26.5%), P (23.1%), and K (36.2%); decreases in the seeds for N (15.4%) and P (8.3%); and an increase in the seeds for K. 2) In leaves, Ca, Mn, and Cu increased, but Mg decreased; whereas in the seeds at first they decreased but later increased. 3) In fruits, Ca, Zn, and Mn were significantly and positively correlated (r=0.906, 0.924, 0.924, P < 0.05) to P in leaves; whereas, P in seeds was significantly and negatively correlated (r=-0.960, P < 0.01) to Ca in leaves. Therefore, to have a positive impact on yield and quality of T. grandis ‘Merrillii', before the fast growth of seeds, more N, P, and Ca fertilizer should be applied; in mid-July, K fertilizer should be added; post-harvest, Mg fertilizer should be applied; and a combined application of microelements and P fertilizer should be added during seed development.
-
Key words:
- cash forestry /
- Torreya grandis ‘Merrillii' /
- leaf /
- seed /
- mineral element /
- dynamic variation
-
图 1 香榧叶片中氮、磷和钾元素的动态变化
Figure 1 Changes of N, P and K contents in leaves of Torreya grandis ‘Merrillii'
图 2 香榧叶片中钙、锾元素的动态变化
Figure 2 Changes of Ca and Mg contents in leaves of Torreya grandis ‘Merrillii'
图 3 香榧叶片中锰、铜、锌元素的动态变化
Figure 3 Changes of Mn, Cu and Zn contents in leaves of Torreya grandis ‘Merrillii'
图 4 香榧种子中氮、磷、钾元素的动态变化
Figure 4 Changes of N, P and K contents in Torreya grandis Torreya grandis ‘Merrillii' fruits
图 5 香榧种子中钙、锾元素的动态变化
Figure 5 Changes of Ga, Mg contents in Torreya grandis ‘Merrillii' fruits
图 6 香榧种子中锰、铜和锌元素的动态变化
Figure 6 Changes of Mn, Cu and Zn contents in Torreya grandis ‘Merrillii' fruits
表 1 香榧叶片和种子中矿质元素之间的相关系数
Table 1. Correlation of mineral elements between leaves and fruits in Torreya grandis ‘Merrillii'
种子矿质元素 叶片矿质元素 氮 憐 钾 钙 镁 锌 铜 锰 N 0.521 0.729 0.456 -0.422 -0.127 0.018 -0.721 -0.272 P 0.546 0.802 0.533 -0.960** 0.378 -0.547 -0.875 -0.922* K -0.148 -0.292 -0.159 0.704 -0.313 0.644 0.383 0.787 Ca 0.713 0.906* 0.646 -0.678 0.012 -0.085 -0.892* -0.523 Mg 0.553 0.835 0.551 -0.924* 0.434 -0.493 -0.910* -0.866 Zn 0.656 0.924* 0.655 -0.812 0.446 -0.264 -0.965** -0.686 Cu 0.563 0.769 0.480 -0.528 -0.195 -0.068 -0.763 -0.391 Mn 0.662 0.924* 0.645 -0.933* 0.379 -0.403 -0.974** -0.840 说明:**表示相关系数在0:01水平极显著相关;*表示相关系数在0.05水平显著相关。  下载: 导出CSV
下载: 导出CSV
-
[1] 黎章矩, 程晓建, 戴文圣, 等.香榧品种起源考证[J].浙江林学院学报, 2005, 22(4):443-448. LI Zhangju, CHENG Xiaojian, DAI Wensheng, et al. Origin of Torreya grandis ‘Merrillii'[J]. J Zhejiang For Coll, 2005, 22(4):443-448. [2] 黎章矩, 戴文圣.中国香榧[M].北京:科学出版社, 2007. [3] 潘立忠, 张森, 王延书, 等.落叶果树果实矿质营养的研究现状[J].安徽农学通报, 2006, 12(10):77-80. Pan Lizhong, ZHANG Sen, WANG Yanshu, et al. Advances in research on mineral nutrition of fruit of common deciduous fruit tree[J]. Auhui Agric Sci Bull, 2006, 12(10):77-80. [4] 陈海霞, 刘明月.观赏植物的矿质营养研究进展[J].湖南农业科学, 2007(5):168-170. CHEN Haixia, LIU Mingyue. Research progress of mineral nutrition of ornamental plant[J]. Hunan Agric Sci, 2007(5):168-170. [5] 丁壮, 张彦东, 齐学军.红松人工林矿质营养元素含量与储量的研究[J].植物研究, 2008, 28(4):503-508. DING Zhuang, ZHANG Yandong, QI Xuejun. Content and storages of mineral nutritive elements in plantations of Pinus koraiensis[J]. Bull Bot Res, 2008, 28(4):503-508. [6] 阴黎明, 王力华, 刘波.文冠果叶片养分元素含量的动态变化及再吸收特性[J].植物研究, 2009, 29(6):685-691. YIN Liming, WANG Lihua, LIU Bo. Dynamic variation and resorption of nutrient elements in the leaves of Xanthoceras sorbifolia Bunge[J]. Bull Bot Res, 2009, 29(6):685-691. [7] 刘桂东, 姜存仓, 王运华, 等.柑橘对不同矿质营养元素效应的研究进展[J].土壤通报, 2010, 41(6):1518-1523. LIU Guidong, JIANG Cuncang, WANG Yunhua, et al. A review on effects of mineral nutrients on Citrus[J]. Chin J Soil Sci, 2010, 41(6):1518-1523. [8] 陈巍, 郭秀珠, 黄品湖, 等.四季柚生育期叶片和果实中矿质元素含量变化的研究[J].植物营养与肥料学报, 2013, 19(3):664-669. CHEN Wei, GUO Xiuzhu, HUANG Pinhu, et al. Variations of mineral elements contents in Sijiyou pummel leaves and fruits during the development period[J]. J Plant Nutr Fert, 2013, 19(3):664-669. [9] 徐爱春, 陈庆红, 顾霞, 等.猕猴桃叶片矿质营养元素含量年变化动态与果实品质的关系[J].湖北农业科学, 2011, 50(24):5126-5130. XU Aichun, CHEN Qinghong, GU Xia, et al. Relationship between fruit quality and changing trend of mineral nutrients in kiwifruit leaves[J]. Hubei Agric Sci, 2011, 50(24):5126-5130. [10] 张春苗.香榧林地土壤肥力及树木营养诊断与施肥研究[D].临安:浙江农林大学, 2011. ZHANG Chunmiao. Soil Fertility in Orchards and Plant Nutrition and Fertilization of Torreya grandis ‘Merrillii'[D]. Lin'an:Zhejiang A & F University, 2011. [11] 吴翔.高温胁迫下香榧叶片的生理生化反应[D].临安:浙江农林大学, 2010. WU Xiang. The Physiological and Biochemical Effect on Leaves under the High Temperature Stress[D]. Lin'an:Zhejiang A & F University, 2010. [12] 戴文圣, 黎章矩, 喻卫武, 等.香榧林地土壤养分状况的调查分析[J].浙江林学院学报, 2006, 23(2):140-144. DAI Wensheng, LI Zhangju, YU Weiwu, et al. Soil nutrients in Torreya grandis ‘Merrillii' plantation[J]. J Zhejiang For Coll, 2006, 23(2):140-144. [13] 鲁如坤, 刘晓松, 陈少华.土壤农业化学分析方法[M].北京:中国农业科技出版社, 2000.146-226. [14] 马建军, 张立彬.野生欧李生长期矿质营养元素含量的变化[J].园艺学报, 2004, 31(2):165-168. MA Jianjun, ZHANG Libin. Change of mineral nutrient elements content in growing period of Cerasus humili[J]. Acta Hort Sin, 2004, 31(2):165-168. [15] 杨光, 简坤, 史继孔.木瓜叶片中矿质元素含量分析[J].中国果树, 2005(1):17-19. YANG Guang, JIAN Kun, SHI Jikong. Study of the content of mineral nutritious elements in Chaenomeles speciosa(Sweet) Nakai leaves[J]. China Fruits, 2005(1):17-19. [16] 解红恩.山核桃植株营养动态变化规律研究[D].临安:浙江农林大学, 2008. XIE Hongen. The Dynamic Changes of Carya cathayensis Sarg. Nutrients[D]. Lin'an:Zhejiang A & F University, 2008. [17] 戴文圣, 黎章矩, 曹福亮, 等.香榧林地土壤及其种子矿物元素含量分析[J].浙江林业科技, 2006, 26(2):14-18. DAI Wensheng, LI Zhangju, CAO Fuliang, et al. Mineral elements in Torreya grandis ‘Merrillii' seeds and their forest land soils[J]. J Zhejiang For Sci Technol, 2006, 26(2):14-18. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2014.05.010
 点击查看大图
点击查看大图
计量
- 文章访问数: 4185
- HTML全文浏览量: 808
- PDF下载量: 571
- 被引次数: 0
