




-
中国每天产生约5万t厨余垃圾,占城市生活垃圾的40%~60%[1]。现有的厨余垃圾处理模式中,饲料化的环境效应最好且成本效益高[2]。利用资源昆虫进行垃圾处理是目前厨余垃圾资源化利用的研究热点,拟黑多刺蚁Polyrhachis vicina幼蚁、黑水虻Hermetia illucens、黄粉虫Tenebrio molitor等昆虫均可以作为资源昆虫处理垃圾[3−5]。黑水虻可有效分解厨余垃圾并将其转化为油脂和蛋白质等高价值生物质[6],黄粉虫幼虫对厨余垃圾的利用率可达38.88%[7]。厨余垃圾含有一定量的金属元素,一般较难对其进行有效处理[8−10]。有研究表明:白蚁与微生物的共同作用,可使木质纤维素材料得到高效的利用[11],据统计,白蚁1 a可以消耗30~70亿t木质纤维素[12]。白蚁还可以通过取食腐烂的有机物和木材,富集重金属[13]。在培菌白蚁肠道和菌圃中存在不同质量分数的金属离子,这些金属离子可能对肠道共生微生物和木质纤维素降解酶的活力有一定影响[14−16]。白蚁肠道系统中许多种酶需要与某些金属离子相结合,来实现其催化活性,这些酶的催化活性在许多生物过程中起到至关重要的作用,例如产氢过程、呼吸过程和碳水化合物水解过程等[17−18]。
黑翅土白蚁Odontotermes formosanus对林木、绿化、堤坝等具有严重危害性,但同时也是一种重要的资源昆虫,在自然环境下,它可以高效降解木质纤维素[19]。黑翅土白蚁是否具有处理厨余垃圾的潜力目前尚不明确。本研究拟探究食物中含有的Al3+、Ca2+、Fe3+和Mg2+对黑翅土白蚁消化代谢的影响,为生产上仿生利用黑翅土白蚁来处理农林废弃物和厨余垃圾提供了新的思路。
-
供试黑翅土白蚁蚁巢采自福建省三明市大田县,带回室内在26 ℃避光条件下饥饿3 d后供处理。供试饵料为桂花Osmanthus fragrans枯枝木屑,采自浙江大学紫金港校区校友林,在80 ℃烘箱内烘48 h,研磨过20目筛;在100 ℃烘箱内烘24 h后备用。无水三氯化铝、无水氯化钙、无水三氯化铁和无水硫酸镁均为分析纯,购自沪试国药集团化学试剂有限公司。漆酶(laccase)和纤维素酶(cellulase)试剂盒购自苏州梦犀生物医药科技有限公司。
-
将无水三氯化铝用去离子水稀释至100.00、10.00、1.00、0.10、0.01 g·L−1 5个质量浓度,以去离子水为对照(ck)。每个处理取10.00 mL,分别与10 g木屑混合均匀,静置24 h后置于80 ℃烘箱中烘干48 h,得到含有Al3+质量分数分别为100.00、10.00、1.00、0.10、0.01 g·kg−1的5组Al3+饵料处理组和ck饵料。将饵料分别装入1.50 mL离心管中并压实,用电子天平称量获得饵料质量。取3个直径一致的蚁巢,在每个蚁巢中同时放入6组饵料。15 d后取出离心管,80 ℃烘干至恒量,去除泥土后称量剩余饵料质量,计算取食率,取平均值。与对照组无显著差异的Al3+最大质量分数为黑翅土白蚁对该金属离子的最大可取食质量分数。其他3种金属离子的最大可取食质量分数测定同上。分别以含最大可取食质量分数的Al3+、Ca2+、Fe3+、Mg2+饵料饲喂黑翅土白蚁,共4个处理组,以去离子水处理的饵料为对照(ck),设3次重复,持续饲喂30 d。
-
参考LI等[16]的方法。在蚁巢中随机挑选10只体长为0.45±0.05 mm 的低龄工蚁置于PTFE烧杯中,加入5.00 mL浓硝酸后,用封口膜密封,依次于80 ℃加热盘上加热1.0 h,160 ℃加热1.5 h,蒸发约0.50 mL。用孔径为0.22 μm的水系滤膜过滤,取0.75 mL滤液用去离子水定容至25.00 mL,得到体积分数约3%的浓硝酸稀释液。每个处理重复3次,使用电感耦合等离子发射光谱仪测定金属元素。
-
参考TOYAMA-KATO等[20]的方法。称取0.1 g上层菌圃于PTFE烧杯中,加入1.00 mL浓硝酸后,用封口膜进行密封。置于60 ℃加热盘上加热4.0 h后向烧杯内加入1.00 mL浓硝酸,再依次在加热盘75 ℃加热0.5 h,130 ℃加热0.5 h,最后200 ℃加热0.5 h。冷却后向烧杯内加入10.00 mL浓硝酸复溶,用孔径为0.22 μm的水系滤膜进行过滤,后续处理同1.2.2。
-
采用分光光度法对黑翅土白蚁工蚁及新建菌圃中的漆酶和纤维素酶活力进行测定,参照试剂盒说明书进行操作。
-
数据均为平均值±标准差,使用SPSS 26进行单因素方差分析,采用LSD法进行多重比较。
-
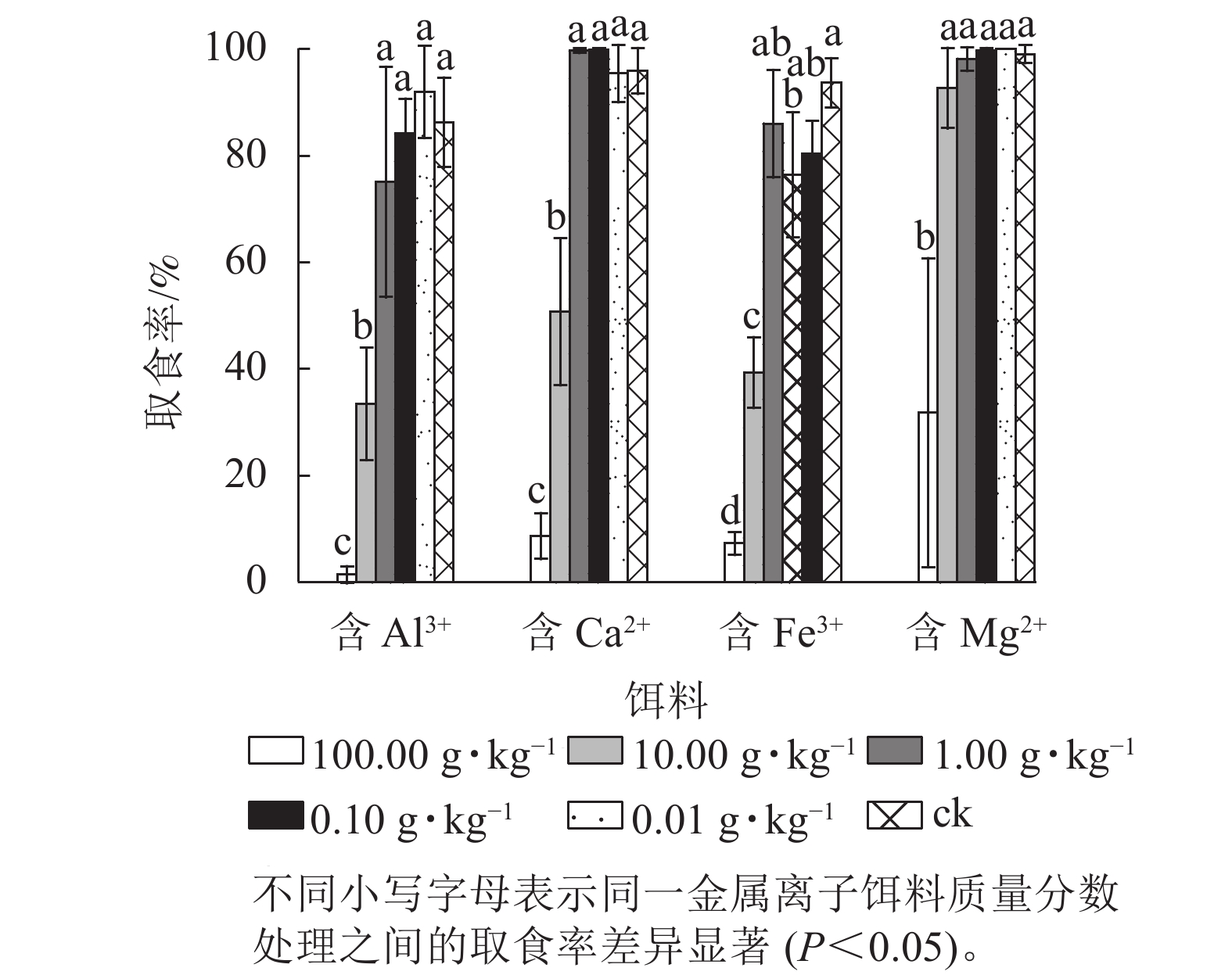
黑翅土白蚁对含Al3+、Ca2+和Fe3+饵料的最大可取食质量分数均为1.00 g·kg−1,对含Mg2+饵料的最大可取食质量分数为10.00 g·kg−1(图1)。对Al3+、Ca2+ 、Fe3+和Mg2+ 4种金属离子而言,黑翅土白蚁总体上偏好取食质量分数较低的饵料。同时,在供试的4种金属离子中,黑翅土白蚁对Mg2+的接受程度最高。

图 1 黑翅土白蚁对不同质量分数金属离子处理饵料的取食率
Figure 1. Bait consumption rate of O. formosanus treated with different mass fraction of metal ions
-
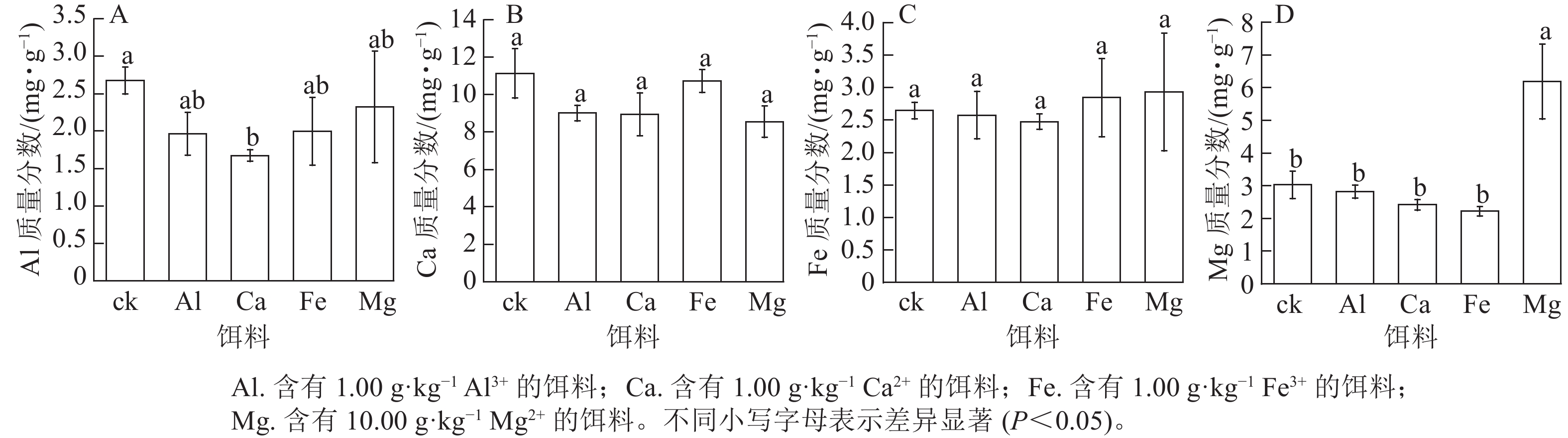
取食含有1.00 g·kg−1 Al3+、Ca2+或Fe3+的饵料后,黑翅土白蚁虫体对应的金属元素Al、Ca和Fe质量分数与ck之间无显著差异(图2A~C)。取食含有1.00 g·kg−1Ca2+的饵料后,黑翅土白蚁虫体内Al质量分数最低,与ck差异显著(P<0.05)。由图2D可见:取食含有10.00 g·kg−1Mg2+饵料后,黑翅土白蚁虫体内Mg 质量分数最高,为6.18 mg·g−1,与ck和取食其他3种金属元素的均差异显著(P<0.05)。说明只有取食含有10.00 g·kg−1 Mg2+的饵料,Mg会在黑翅土白蚁体内富集,而其他3种金属元素不会在黑翅土白蚁体内富集。
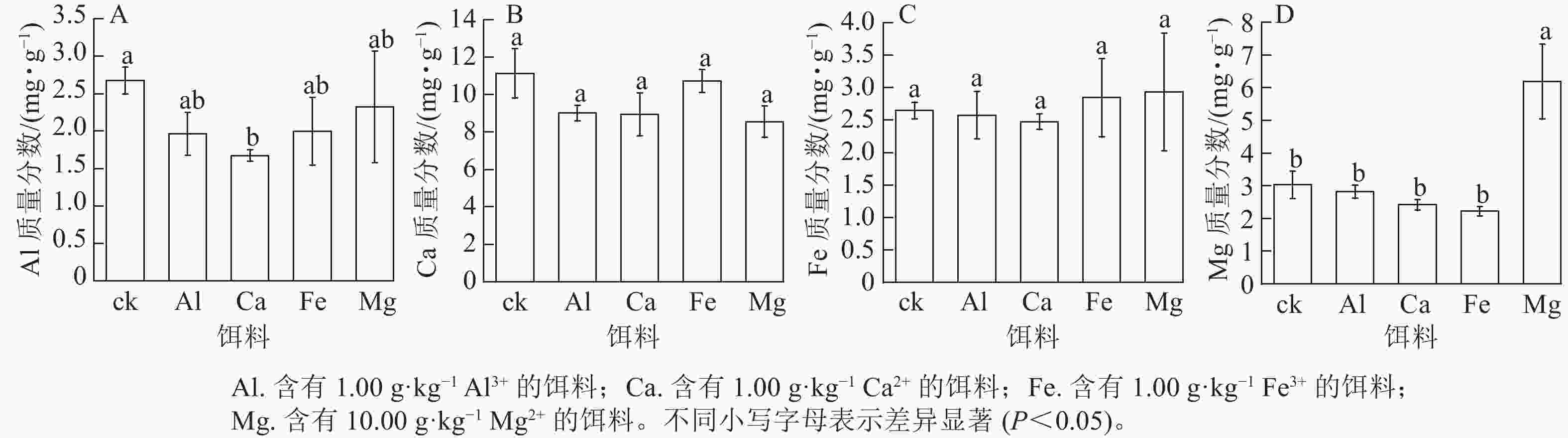
图 2 饵料饲喂后黑翅土白蚁虫体内金属元素Al、Ca、Fe和Mg质量分数
Figure 2. Mass fraction of metal elements Al, Ca, Fe and Mg in the body of O. formosanus after feeding with baits
-
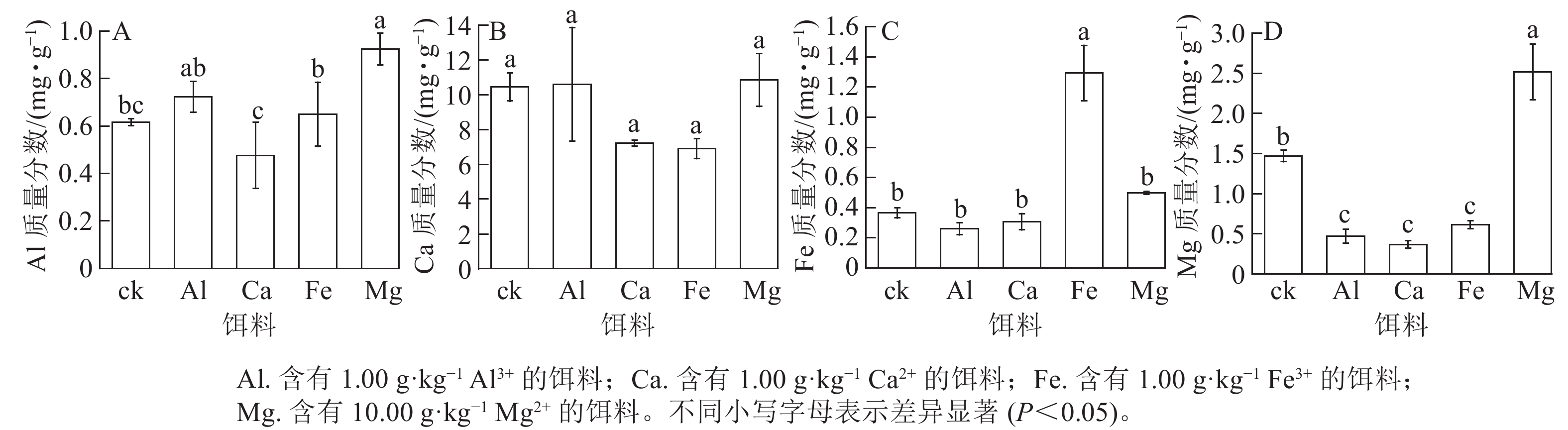
取食含有1.00 g·kg−1 Al3+或Ca2+的饵料后,其菌圃内对应的金属元素Al和Ca质量分数与ck之间无显著差异(图3A~B)。取食含有1.00 g·kg−1 Fe3+或含有10.00 g·kg−1Mg2+的饵料后,菌圃内对应的Fe质量分数为1.29 mg·g−1,Mg为2.52 mg·g−1,均显著高于ck (P<0.05)(图3C~D)。其中取食含有10.00 g·kg−1Mg2+饵料后菌圃内的Al质量分数较ck显著提高(P<0.05),取食含有1.00 g·kg−1 Al3+、Ca2+、Fe3+的饵料后,菌圃中Mg质量分数较ck显著降低(P<0.05)。说明取食含金属离子的饵料对黑翅土白蚁新建菌圃内相应金属元素质量分数有一定影响。就本研究而言,黑翅土白蚁取食含有1.00 g·kg−1 Fe3+或含有10.00 g·kg−1·Mg2+饵料后,对应的金属元素Fe和Mg会在菌圃中富集。
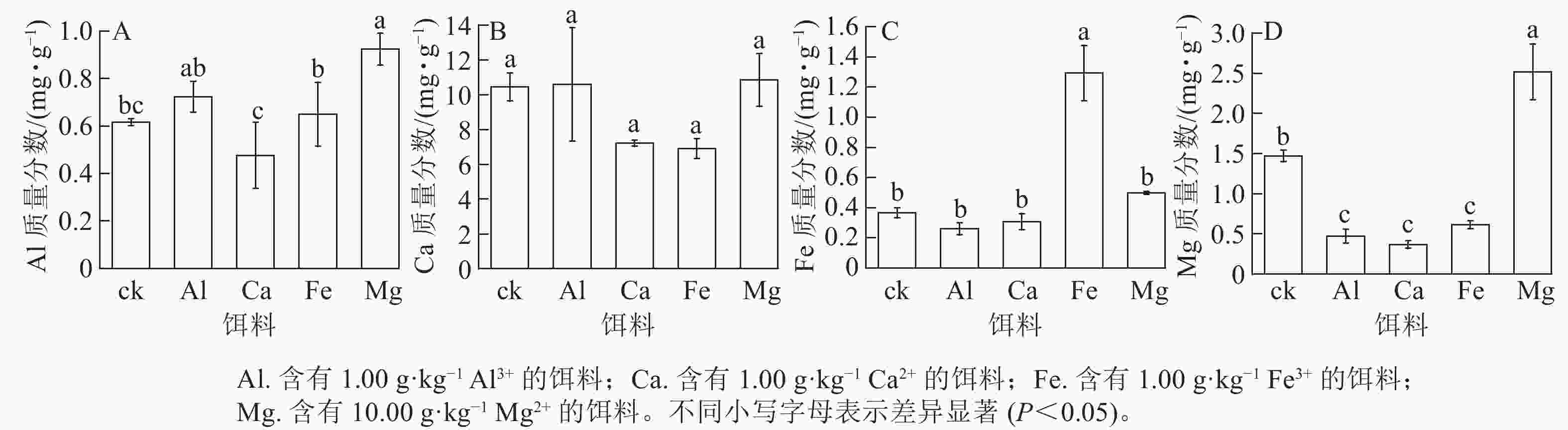
图 3 饵料饲喂后新建菌圃内金属元素Al、Ca、Fe和Mg质量分数
Figure 3. Mass fraction of metal elements Al, Ca, Fe and Mg in new-built fungus combs after feeding with baits
-
表1数据表明:1.00 g·kg−1 Al3+、Ca2+和Fe3+显著促进了黑翅土白蚁虫体内的漆酶活性(P<0.05),说明上述质量分数的3种金属离子可提高白蚁的木质素降解能力;1.00 g·kg−1 Fe3+显著抑制了新建菌圃内的漆酶活性(P<0.05),其酶活性仅为62.91 nmol·g−1·min−1,但被摄入食物中的10.00 g·kg−1 Mg2+显著促进(P<0.05),酶活性达到了419.65 nmol·g−1·min−1,说明Fe在菌圃富集会降低菌圃微生物的木质素降解能力,而Mg正好相反。1.00 g·kg−1 Ca2+和Fe3+显著抑制了黑翅土白蚁虫体内的纤维素酶活性(P<0.05),但10.00 g·kg−1 Mg2+则对该酶起到显著促进作用(P<0.05),说明不同金属离子在白蚁体内可能存在竞争关系,且Mg的富集会提高黑翅土白蚁虫体内的纤维素降解能力。1.00 g·kg−1 Fe3+显著促进了新建菌圃中的纤维素酶活性(P<0.05),其酶活性为1 098.77 μg·g−1·min−1,但10.00 g·kg−1 Mg2+显著抑制了新建菌圃中的纤维素酶活性(P<0.05),其酶活性为745.40 μg·g−1·min−1。说明Fe在菌圃富集会提高菌圃微生物的纤维素降解能力,而Mg在菌圃富集则会降低菌圃微生物的纤维素降解能力。
表 1 黑翅土白蚁虫体内及新建菌圃内漆酶和纤维素酶活性
Table 1. Activities of laccase and cellulase in the body of O. formosanus and in new-built fungus combs
处理 漆酶活性/(nmol·g−1·min−1) 纤维素酶活性/(μg·g−1·min−1) 虫体 菌圃 虫体 菌圃 ck 2.84±0.66 c 227.79±18.67 b 1129.21±111.55 b 923.25±31.04 b Al 15.65±2.48 a 151.11±17.25 bc 1253.91±54.33 b 932.67±36.95 b Ca 9.81±0.44 b 146.36±40.72 bc 682.67±70.10 c 907.02±34.90 b Fe 7.38±1.30 b 62.91±7.74 c 615.41±83.35 c 1098.77±108.31 a Mg 1.90±0.64 c 419.65±18.62 a 1600.71±13.19 a 745.40±32.09 c 说明:Al指含有1.00 g·kg−1 Al3+饵料;Ca指含有1.00 g·kg−1 Ca2+饵料;Fe指含有1.00 g·kg−1 Fe3+饵料;Mg指含有10.00 g·kg−1 Mg2+饵料。不同小写字母表示不同处理间差异显著(P<0.05)。 -
白蚁富含蛋白质、维生素和许多必需的矿物质元素[21] ,可以食用;培菌白蚁的共生真菌蚁巢伞,味道鲜美,营养价值高,研究还发现蚁巢伞属真菌具有镇痛抗炎、防癌抗癌、降血脂及抗氧化等药理作用和生物功能活性[22]。在本研究中,取食含有1.00 g·kg−1 Al3+、Ca2+、Fe3+或10.00 g·kg−1 Mg2+的饵料后,黑翅土白蚁仍可进行正常的生理活动并构建菌圃,由此可以看出黑翅土白蚁具有处理含有一定质量分数金属离子的厨余垃圾和农林废弃物的潜力。
培菌白蚁可以在肠道微生物的帮助下有效分解木质纤维素,菌圃内的细菌[23]可以通过产生包括纤维素酶在内的大量降解木质纤维素的酶,来实现与蚁巢伞协同降解木质纤维素的功能[24]。培菌白蚁的共生蚁巢伞可以产生纤维素酶、果胶质水解酶类、木聚糖酶等,同时也能产生与木质素分解有关的漆酶,培菌白蚁可以利用共生蚁巢伞高效降解木质纤维素[25−26]。LI等[27]发现Al3+对解淀粉芽孢杆菌漆酶活性有促进作用,与本研究中1.00 g·kg−1 Al3+能够促进黑翅土白蚁虫体内的漆酶活性结果一致,在饵料中添加1.00 g·kg−1 Al3+有利于提高黑翅土白蚁及其菌圃微生物联合降解木质素的能力,而不会显著影响纤维素降解能力,对黑翅土白蚁的资源性利用具有现实意义。肖东来等[28]研究发现Mg2+对香菇 Lentinuda edodes 羧甲基纤维素酶活性具有激活作用,而WANG等[29]的研究结果表明较高质量分数的Mg2+能够抑制黄粉虫纤维素酶活力。在本研究中,10.00 g·kg−1 Mg2+抑制了菌圃内的纤维素酶活性,却促进了黑翅土白蚁虫体内的纤维素酶活性,一方面是因为同种金属离子对不同来源的同一类酶的影响程度可能不同[30];另一方面,LI等[19]的研究已经证实白蚁和菌圃微生物对木质素的降解具有互补性,推测黑翅土白蚁能够通过自身和菌圃微生物对木质纤维素降解的互补性来抵抗一定质量分数金属离子的负面影响,有关金属离子对木质纤维素降解酶的联合影响有待进一步研究。
-
本研究确定了黑翅土白蚁对饵料中Al3+、Ca2+和Fe3+的最大可取食质量分数均为1.00 g·kg−1,对饵料中Mg2+的最大可取食质量分数为10.00 g·kg−1。金属元素Mg能够在黑翅土白蚁虫体内和菌圃内富集,不同金属离子对黑翅土白蚁虫体和菌圃内漆酶和纤维素酶活性的影响不同,其中1.00 g·kg−1 Al3+可以提高黑翅土白蚁及其菌圃微生物联合降解木质素的能力。黑翅土白蚁具有对含有金属离子的厨余垃圾和农林废弃物进行资源化处理的应用潜力,今后需要进一步研究混合金属离子对黑翅土白蚁消化代谢的影响,以推动其资源化处理厨余垃圾和农林废弃物的实际应用。
Effects of metal ions on digestion and metabolism of Odontotermes formosanus
-
摘要:
目的 探究金属离子对黑翅土白蚁Odontotermes formosanus消化代谢过程的影响,挖掘黑翅土白蚁对含有金属离子的厨余垃圾和农林废弃物进行资源化处理的潜在价值。 方法 根据浓度梯度法,将含有不同质量分数Al3+、Ca2+、Fe3+和Mg2+的饵料供给黑翅土白蚁,确定黑翅土白蚁对饵料中金属离子的最大可取食质量分数。采用电感耦合等离子体发射光谱仪(ICP-OES)测定黑翅土白蚁虫体内及其新建菌圃内的对应金属元素质量分数,明确饵料中Al3+、Ca2+、Fe3+和Mg2+对虫体内及新建菌圃内对应金属元素质量分数的影响。采用试剂盒法测定黑翅土白蚁虫体内及其新建菌圃内漆酶和纤维素酶的活性,确定饵料中Al3+、Ca2+、Fe3+和Mg2+对上述酶活性的影响。 结果 黑翅土白蚁对饵料中Al3+、Ca2+和Fe3+的最大可取食质量分数均为1.00 g·kg−1,对饵料中Mg2+的最大可取食质量分数为10.00 g·kg−1。黑翅土白蚁取食含有10.00 g·kg−1 Mg2+的饵料后,其虫体内和菌圃内都发生了Mg的富集;取食含有1.00 g·kg−1 Fe3+的饵料后,Fe仅在菌圃内富集。取食含有1.00 g·kg−1 Al3+的饵料,会显著提高黑翅土白蚁虫体内的漆酶活性(P<0.05);取食含有1.00 g·kg−1 Ca2+的饵料,会显著提高黑翅土白蚁虫体内的漆酶活性(P<0.05),但会显著降低纤维素酶活性(P<0.05);取食含有1.00 g·kg−1 Fe3+的饵料,会显著提高黑翅土白蚁虫体内的漆酶活性和菌圃中的纤维素酶活性(P<0.05),但会显著降低菌圃中的漆酶活性和黑翅土白蚁虫体内的纤维素酶活性(P<0.05);取食含有10.00 g·kg−1 Mg2+的饵料,会显著提高黑翅土白蚁虫体内的纤维素酶活性和菌圃中的漆酶活性(P<0.05),但会显著降低菌圃中的纤维素酶活性(P<0.05)。 结论 黑翅土白蚁可取食分别含有1.00 g·kg−1 Al3+、Ca2+和Fe3+的饵料和含有10.00 g·kg−1 Mg2+的饵料,1.00 g·kg−1 Al3+可以提高黑翅土白蚁和菌圃微生物对木质素的联合降解能力。黑翅土白蚁具有资源化处理厨余垃圾和农林废弃物的应用潜力。图3表1参30 Abstract:Objective This study aims to explore the role of metal ions in influencing the digestive generation process of Odontotermes formosanus, which is beneficial to explore the potential value of resource treatment of agricultural and forestry waste and kitchen waste containing metal ions by O. formosanus. Method According to the concentration gradient method, the bait containing different mass fraction of Al3+, Ca2+, Fe3+ and Mg2+ was fed to O. formosanus to determine the maximum edible mass fraction of metal ions in bait for O. formosanus. ICP-OES was used to measure the corresponding metal elements in O. formosanus body and new-built fungus combs, and to obtain the effects of feeding baits containing Al3+, Ca2+, Fe3+ and Mg2+ on the corresponding metal elements mass fraction in O. formosanus body and new-built fungus combs. The activities of laccase and cellulase in O. formosanus body and new-built fungus combs were determined by kit method, and the effects of feeding baits containing Al3+, Ca2+, Fe3+ and Mg2+ on the activities of these enzymes were determined. Result The maximum edible mass fraction of Al3+, Ca2+ and Fe3+ in baits of O. formosanus was 1.00 g·kg−1, and Mg2+ in baits was 10.00 g·kg−1. Mg enrichment occurred both in O. formosanus body and new-built fungus combs after feeding baits containing 10.00 g·kg−1 Mg2+; Fe was only enriched in new-built fungus combs after feeding baits containing 1.00 g·kg−1 Fe3+. The laccase activity in O. formosanus body improved significantly after feeding baits containing 1.00 g·kg−1 Al3+. The laccase activity in O. formosanus body improved significantly, but the cellulase activity reduced significantly after feeding baits containing 1.00 g·kg−1 Ca2+. The laccase activity in O. formosanus body and cellulase activity in new-built fungus combs improved significantly, but the laccase activity in new-built fungus combs and cellulase activity in O. formosanus body reduced significantly after feeding baits containing 1.00 g·kg−1 Fe3+. The cellulase activity in O. formosanus body and the laccase activity in new-built fungus combs improved significantly, but the cellulase activity in new-built fungus combs reduced significantly after feeding foods containing 10.00 g·kg−1 Mg2+. Conclusion O. formosanus can feed on baits containing 1.00 g·kg−1 Al3+, Ca2+ or Fe3+ or 10.00 g·kg−1 Mg2+, and 1.00 g·kg−1 Al3+ can improve the combined degradation of lignin by O. formosanus and fungus-comb microbiome. O. formosanus have application potential for resource treatment of agricultural and forestry waste and kitchen waste. [Ch, 3 fig. 1 tab. 30 ref.] -
Key words:
- Odontotermes formosanus /
- fungus combs /
- metal ion /
- enzyme activity
-
图 1 黑翅土白蚁对不同质量分数金属离子处理饵料的取食率
Figure 1 Bait consumption rate of O. formosanus treated with different mass fraction of metal ions
图 2 饵料饲喂后黑翅土白蚁虫体内金属元素Al、Ca、Fe和Mg质量分数
Figure 2 Mass fraction of metal elements Al, Ca, Fe and Mg in the body of O. formosanus after feeding with baits
图 3 饵料饲喂后新建菌圃内金属元素Al、Ca、Fe和Mg质量分数
Figure 3 Mass fraction of metal elements Al, Ca, Fe and Mg in new-built fungus combs after feeding with baits
表 1 黑翅土白蚁虫体内及新建菌圃内漆酶和纤维素酶活性
Table 1. Activities of laccase and cellulase in the body of O. formosanus and in new-built fungus combs
处理 漆酶活性/(nmol·g−1·min−1) 纤维素酶活性/(μg·g−1·min−1) 虫体 菌圃 虫体 菌圃 ck 2.84±0.66 c 227.79±18.67 b 1129.21±111.55 b 923.25±31.04 b Al 15.65±2.48 a 151.11±17.25 bc 1253.91±54.33 b 932.67±36.95 b Ca 9.81±0.44 b 146.36±40.72 bc 682.67±70.10 c 907.02±34.90 b Fe 7.38±1.30 b 62.91±7.74 c 615.41±83.35 c 1098.77±108.31 a Mg 1.90±0.64 c 419.65±18.62 a 1600.71±13.19 a 745.40±32.09 c 说明:Al指含有1.00 g·kg−1 Al3+饵料;Ca指含有1.00 g·kg−1 Ca2+饵料;Fe指含有1.00 g·kg−1 Fe3+饵料;Mg指含有10.00 g·kg−1 Mg2+饵料。不同小写字母表示不同处理间差异显著(P<0.05)。  下载: 导出CSV
下载: 导出CSV
-
[1] 刘雪松, 沈骏, 刘雪莲. 厨余垃圾资源化利用技术研究进展[J]. 现代化工, 2023, 43(4): 23 − 26, 31. LIU Xuesong, SHEN Jun, LIU Xuelian. Research progress of kitchen waste resource utilization technology [J]. Modern Chemical Industry, 2023, 43(4): 23 − 26, 31. [2] 李欢, 周颖君, 刘建国, 等. 我国厨余垃圾处理模式的综合比较和优化策略[J]. 环境工程学报, 2021, 15(7): 2398 − 2408. LI Huan, ZHOU Yingjun, LIU Jianguo, et al. Comprehensive comparison and optimal strategies of food waste treatment modes [J]. Chinese Journal of Environmental Engineering, 2021, 15(7): 2398 − 2408. [3] FU Shanfei, WANG Donghui, XIE Zhong, et al. Producing insect protein from food waste digestate via black soldier fly larvae cultivation: a promising choice for digestate disposal [J/OL]. Science of the Total Environment, 2022, 830: 154654[2023-03-20]. doi:10.1016/j.scitotenv.2022.154654. [4] JAGTAP S, GARCIA-GARCIA G, DUONG L, et al. Codesign of food system and circular economy approaches for the development of livestock feeds from insect larvae [J/OL]. Foods, 2021, 10(8): 1701[2023-03-20]. doi: 10.3390/foods10081701. [5] 张连俊. 黄粉虫和黑水虻联合转化厨余垃圾及虫沙应用初探[D]. 乌鲁木齐: 新疆农业大学, 2021. ZHANG Lianjun. A Preliminary Study on the Application of Combined Conversion of Food Waste and Worm Sand by Tenebrio Molitor and Hermetia illucens [D]. Urumqi: Xinjiang Agricultural University, 2021. [6] YIN Jingkai, GONG Xiaoyan, SUN Lina, et al. Study on the transformation of nutrients in kitchen waste by black soldier fly [J]. Journal of Agricultural Science and Technology, 2021, 23(6): 154 − 159. [7] CHEN Meiling, LING Yuanzhi, HUANG Ruqiang, et al. Optimization of feeding conditions of Tenebrio molitor Linnaeus larvae in food wastes treatment using response surface methodology [J]. Chinese Journal of Environmental Engineering, 2015, 9(5): 2455 − 2461. [8] ILMASARI D, KAMYAB H, YUZIR A, et al. A review of the biological treatment of leachate: available technologies and future requirements for the circular economy implementation [J/OL]. Biochemical Engineering Journal, 2022, 187: 108605[2023-03-20]. doi:10.1016/j.bej.2022.108605. [9] DANG Meirong, CHAI Junrui, XU Zengguang, et al. Soil water characteristic curve test and saturated-unsaturated seepage analysis in Jiangcungou municipal solid waste landfill, China [J/OL]. Engineering Geology, 2020, 264: 105374[2023-03-20]. doi: 10.1016/j.enggeo.2019.105374. [10] GU Binxian, JIANG Suqin, WANG Haikun, et al. Characterization, quantification and management of China’s municipal solid waste in spatiotemporal distributions: a review [J]. Waste Management, 2017, 61: 67 − 77. [11] 蒋宇彤, 张硕, 林子佳, 等. 白蚁消化系统转化和降解木质纤维素酶研究进展[J]. 微生物学报, 2020, 60(12): 2635 − 2649. JIANG Yutong, ZHANG Shuo, LIN Zijia, et al. Advances in lignocellulose-degrading enzymes from termites and symbiotic microbes [J]. Acta Microbiologica Sinica, 2020, 60(12): 2635 − 2649. [12] TOKUDA G, TSUBOI Y, KIHARA K, et al. Metabolomic profiling of C-13-labelled cellulose digestion in a lower termite: insights into gut symbiont function [J/OL]. Proceedings of the Royal Society B-Biological Sciences, 2014, 281(1789): 2014090[2023-03-20]. doi: 10.1098/rspb.2014.0990. [13] DENLOYE A A, ABDULSALAM L, BAKRE S, et al. Heavy metals in some termites species and their nests in Ojo, Lagos, Nigeria [J]. Animal Research International, 2015, 12(2): 2178 − 2183. [14] BALLOR N R, LEADBETTER J R. Analysis of extensive [FeFe] hydrogenase gene diversity within the gut microbiota of insects representing five families of dictyoptera [J]. Microbial Ecology, 2012, 63(3): 586 − 595. [15] ARANTES V, JELLISON J, GOODELL B. Peculiarities of brown-rot fungi and biochemical fenton reaction with regard to their potential as a model for bioprocessing biomass [J]. Applied Microbiology and Biotechnology, 2012, 94(2): 323 − 338. [16] LI Hongjie, SUN Jianzhong, ZHAO Jianming, et al. Physicochemical conditions and metal ion profiles in the gut of the fungus-growing termite Odontotermes formosanus [J]. Journal of Insect Physiology, 2012, 58(10): 1368 − 1375. [17] LIU Ning, YAN Xing, ZHANG Meiling, et al. Microbiome of fungus-growing termites: a new reservoir for lignocellulase genes [J]. Applied and Environmental Microbiology, 2011, 77(1): 48 − 56. [18] VU A T, NGUYEN N C, LEADBETTER J R. Iron reduction in the metal-rich guts of wood-feeding termites [J]. Geobiology, 2004, 2(4): 239 − 247. [19] LI Hongjie, YELLE D J, LI Chang, et al. Lignocellulose pretreatment in a fungus-cultivating termite [J]. Proceedings of the National Academy of Sciences of the United States of America, 2017, 114: 4709 − 4714. [20] TOYAMA-KATO Y, YOSHIDA K, FUJIMORI E, et al. Analysis of metal elements of hydrangea sepals at various growing stages by ICP-AES [J]. Biochemical Engineering Journal, 2003, 14(3): 237 − 241. [21] NETSHIFHEFHE S R, KUNJEKU E C, DUNCAN F D. Human uses and indigenous knowledge of edible termites in Vhembe District, Limpopo Province, South Africa [J/OL]. South African Journal of Science, 2018, 114(1/2): 2017-0145[2023-03-20]. doi: 10.17159/sajs.2018/20170145. [22] NHI N T N, KHANG D T, DUNG T N. Termitomyces mushroom extracts and its biological activities [J/OL]. Food Science and Technology, 2022, 42: e125921[2023-03-20]. doi: 10.1590/fst.125921. [23] 朱娅宁, 羊桂英, 周琪欢, 等. 黑翅土白蚁菌圃微生物对蚁巢伞生长的影响[J]. 浙江农林大学学报, 2022, 39(3): 598 − 606. ZHU Yaning, YANG Guiying, ZHOU Qihuan, et al. Impact of microorganisms of Odontotermes formosanus fungus-combs on the growth of Termitomyces heimii [J]. Journal of Zhejiang A&F University, 2022, 39(3): 598 − 606. [24] POULSEN M, HU Haofu, LI Cai, et al. Complementary symbiont contributions to plant decomposition in a fungus-farming termite [J]. Proceedings of the National Academy of Sciences of the United States of America, 2014, 111(40): 14500 − 14505. [25] NI Jinfeng, TOKUDA G. Lignocellulose-degrading enzymes from termites and their symbiotic microbiota [J]. Biotechnology Advances, 2013, 31(6): 838 − 850. [26] 沈毅, 漆梦雯, 羊桂英, 等. 黑翅土白蚁共生真菌对水稻秸秆生物降解研究[J]. 浙江农林大学学报, 2023, 40(6): 1224 − 1231. SHEN Yi, QI Mengwen, YANG Guiying, et al. Biodegradation of rice straw by symbiotic fungi of Odontotermes formosanus [J]. Journal of Zhejiang A&F University, 2023, 40(6): 1224 − 1231. [27] 李国富, 栗君, 卢磊, 等. 解淀粉芽孢杆菌LC03的分离及其芽孢漆酶性质研究[J]. 北京林业大学学报, 2013, 35(3): 116 − 121. LI Guofu, LI Jun, LU Lei, et al. Isolation of laccase-producing strain Bacillus amyloliquefaciens LC03 and characterization of its spore laccase [J]. Journal of Beijing Forestry University, 2013, 35(3): 116 − 121. [28] 肖冬来, 张迪, 林衍铨, 等. 金属离子对香菇纤维素酶、漆酶和木质素过氧化物酶活性的影响[J]. 中国食用菌, 2018, 37(3): 56 − 58. XIAO Donglai, ZHANG Di, LIN Yanquan, et al. Effects of metal ions on the carboxymethyl cellulase, laccase and lignin peroxidase activities of Lentinula edodes [J]. Edible Fungi of China, 2018, 37(3): 56 − 58. [29] 王国红, 耿俊丽, 黄祥财. 9种金属离子对黄粉虫纤维素酶活性及其生长发育的影响[J]. 福建师范大学学报(自然科学版), 2009, 25(3): 101 − 104, 118. WANG Guohong, GENG Junli, HUANG Xiangcai. Effects of 9 kinds of metal Ions on cellulase activity of Tenebrio molitor L. and the growth of larvae [J]. Journal of Fujian Normal University (Natural Science Edition), 2009, 25(3): 101 − 104, 118. [30] 安刚, 陶毅明, 龙敏南, 等. 金属离子对白蚁纤维素酶活力的影响[J]. 厦门大学学报(自然科学版), 2008, 47(增刊2): 107 − 109. AN Gang, TAO Yiming, LONG Minnan, et al. Effects of metal ion on the activity of cellulase in termites [J]. Journal of Xiamen University (Natural Science), 2008, 47(suppl 2): 107 − 109. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20230219
 点击查看大图
点击查看大图
计量
- 文章访问数: 1785
- HTML全文浏览量: 440
- PDF下载量: 37
- 被引次数: 0
