





-
线粒体广泛存在于动物和植物的每个个体之中,作为真核细胞中一种半自主的细胞器,能够满足生命过程如生长发育、新陈代谢及移动所需要的能源需求,对诸如细胞功能调节、老龄衰退或死亡等现象发挥着重要的作用[1−2]。相较于其他类型的基因而言,线粒体结构简单且遵循母系遗传,同时演化速度更快,变异频率也更高[3]。昆虫线粒体基因组是一串长约14~19 bp的闭合双链环状DNA分子,共有37个基因,由13个蛋白质编码基因(PCGs)、22个转运RNA(tRNAs)以及2个核糖体RNA(rRNAs)构建而成[4]。通过线粒体基因组研究昆虫的进化规律和系统发育关系已经成为成熟的手段[5]。
线粒体基因组技术主要应用于昆虫起源、分子进化、系统发育等研究领域,涉及到包括鞘翅目Coleoptera、半翅目Hemiptera、膜翅目Hymenoptera等在内的昆虫[6−10]。鳞翅目Lepidoptera作为昆虫纲中第二大目,害虫种类繁多,因此在昆虫研究中具有重要的代表性[11]。暗影饰皮夜蛾Garella ruficirra隶属于鳞翅目夜蛾科Noctuidae丽夜蛾亚科Chloephorinae[12],在河北、河南、山东、江苏、浙江和江西等省均有不同程度的危害发生[13]。暗影饰皮夜蛾是薄壳山核桃Carya illinoinensis、板栗Castanea mollissima等经济林的重要害虫[12, 14],薄壳山核桃害虫种类繁多[15],近年来暗影饰皮夜蛾在薄壳山核桃林内新发生且呈现逐年上升的趋势,现有的研究多数集中在发生情况的简报和防治建议方面[16−17]。对其遗传特征、与同类昆虫之间的亲缘关系研究还未见报道。因此,对暗影饰皮夜蛾进行全面的线粒体全基因组分析,除了能丰富对夜蛾科尤其是丽夜蛾亚科昆虫线粒体基因组的了解外,还能为暗影饰皮夜蛾的分子系谱、群落遗传特性以及分子生态环境等研究提供数据支持。
本研究通过对暗影饰皮夜蛾线粒体的碱基组成、起始密码子、终止密码子、二级结构等进行研究,将其与已经公布的鳞翅目昆虫的线粒体基因组相比较,通过最大似然法和贝叶斯法联合构建系统发育树,以期分析暗影饰皮夜蛾线粒体基因组特征与系统发育关系,为暗影饰皮夜蛾的演变历程和系统发展提供分子证据。
-
2022年10月,在江苏省东台市(32°47′42″N,120°31′08″E)薄壳山核桃林中采集暗影饰皮夜蛾幼虫危害的薄壳山核桃果实,带回实验室。解剖出来的幼虫放置在−80 ℃超低温冰箱保存,用于提取DNA。选择龄期一致的幼虫个体作为研究对象,利用DNA提取试剂盒(德国Qiagen公司)提取暗影饰皮夜蛾的总DNA,使用质量浓度为1%的琼脂糖凝胶检测该幼虫DNA的纯度和浓度。
-
检测总DNA质量后,采用全基因组鸟枪法(WGS),使用Illumina Miseq平台进行双端测序(PE)构建文库,在得到原始序列后,通过FastQC 平台对原始序列进行质量剪切和过滤:包括去除读长(reads)中的接头(adapter)序列,剪切去除5′端包含有非腺嘌呤(A)、胸腺嘧啶(T)、胞嘧啶(C)、鸟嘌呤(G)的碱基;修剪低质量的reads末端;去除无法确定的碱基信息(N)含量大于5的reads;去掉adapter以及经过质量修剪之后长度不足25 bp的小片段,最终得到高质量的干净数据(clean data)[18−19]。
-
采用A5-miseq v20150522以及SPAdes v 3.10.0软件对获得的高质量clean data进行从头组装,从而得到重叠群(contig)和支架(scaffold)序列[20−22]。使用Mummer v 3.1软件进一步剔除线粒体基因组序列中模糊核苷酸和质量平均值低于G30(质量值大于等于30的碱基所占比例)的reads,进行数据重组,得到contig和scaffold。将完整的线性contigs上传到MITOS网页服务器(
http://mitos2.bioinf.uni-leipzig.de/index.py )进行功能注释[22−23]。选择“Genetic Code”的可选设置为05-inverterbrate,其余设置按照默认参数进行选择。使用Organella Genome Draw网络服务器工具(http://ogdraw.mpimp-golm.mpg.de/ )对样本的环状线粒体基因组进行可视化处理[22, 24]。按照拼接序列的测序深度,使用美国国家生物技术信息中心(NCBI)中的nt文库进行高测序深度的BLAST分析,并将其与拼接后的序列进行比较,获得预测基因的注释信息[22]。将序列上传至NCBI数据库中,获得登录号为ON125428。 -
使用
网址进行暗影饰皮夜蛾线粒体基因组圈图的绘制,接着使用MEGA 6软件计算G. ruficirra包括AT含量、AT偏斜以及GC偏斜等在内的核苷酸组成情况,同时对蛋白质编码基因的氨基酸使用情况和相对同义密码子使用度(RSCU)进行分析[22]。应用tRNAscan-SE Search Server v1.21[25]预测暗影饰皮夜蛾tRNA基因的二级结构模型,分析暗影饰皮夜蛾的碱基组成差异。结合区域、螺旋、螺旋跨膜区域和其他不规则区域,对ATP8蛋白的二级结构进行预测,同时对ATP8蛋白的氨基酸组成和编码序列组成进行分析。 -
基于夜蛾科13个种昆虫线粒体全基因组的核苷酸序列,选择天蛾科Sphingidae 2个种作为外群,使用
http://www.phylo.org 网址联合建立最大似然法(ML)和贝叶斯法(BI)系统发育树。以NCBI下载的12种夜蛾科昆虫的序列作为参考,与3种天蛾科昆虫的序列联合进行系统发育分析。通过贝叶斯框架和BEAST v 1.6.1软件包对克隆进化的分化时间进行估算,同时选用J model test软件得到最佳替代模型GTR+I+G (表1)。
表 1 用于线粒体基因组分析的昆虫物种信息
Table 1. Information of the insect species for phylogenetic analysis in mitochondrial genome
类群 总科 属 物种 内群 夜蛾科Noctuidae Garella 暗影饰皮夜蛾G. ruficirra G. musculana G. rotundipennis G. nilotica G. curiosa 皮夜蛾属Nycteola N. indica 亚皮夜蛾N. asiatica 饰纹夜蛾属Antoculeora 饰银纹夜蛾A. ornatissima Ctenoplusia 白条夜蛾C. albostriata C. ogovana 银纹夜蛾C. agnata 饰夜蛾属Pseudoips P. prasinana 外群 天蛾科Sphingidae 蛀野螟属Conogethes 桃蛀螟C. punctiferalis 秆野螟属Ostrinia 玉米螟O. nubilalis -
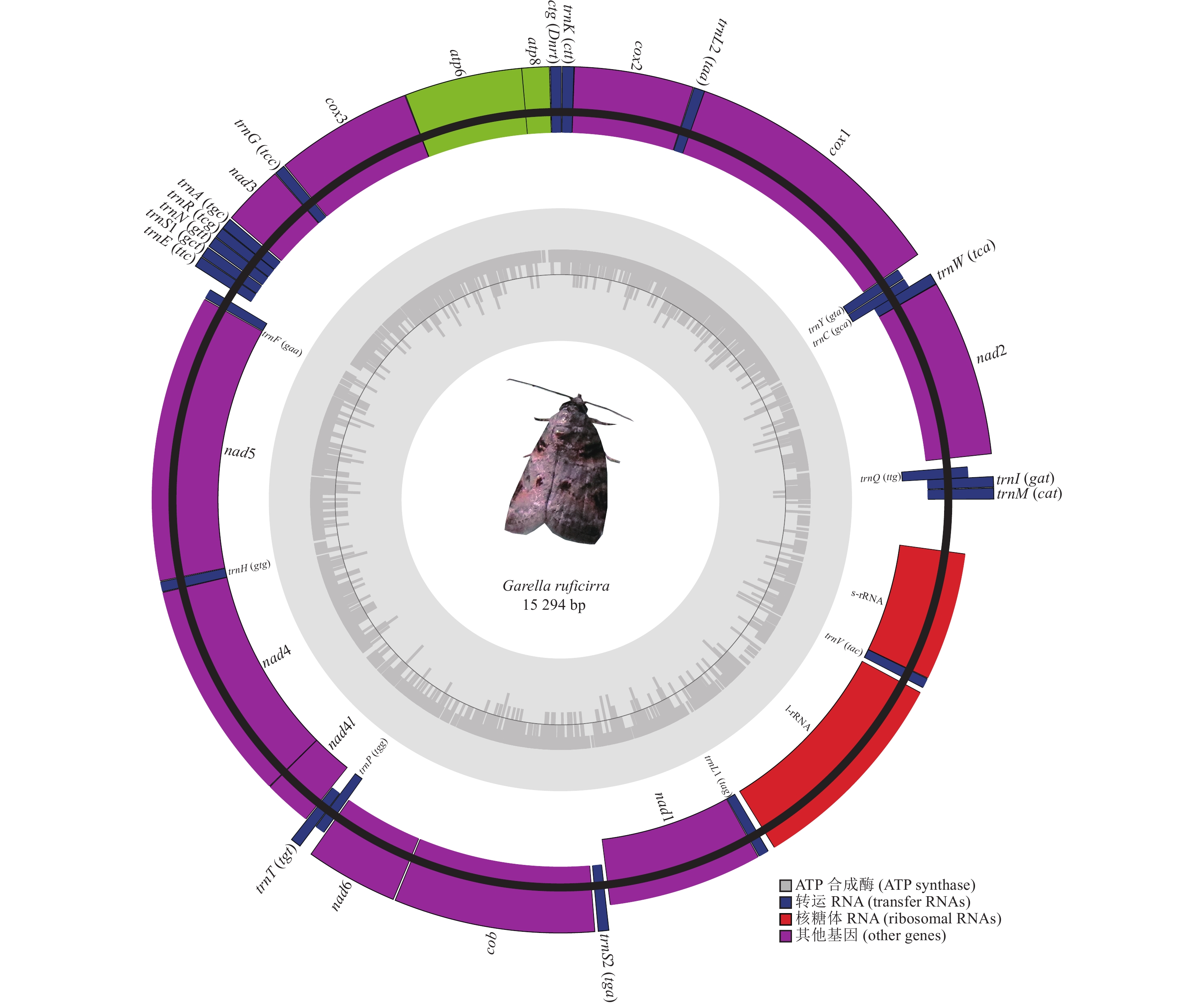
暗影饰皮夜蛾线粒体基因组全长为15 294 bp,呈闭合环状的双链结构,整个线粒体基因组编码了37个基因,其中包括13个蛋白质编码基因(PCGs)、22个tRNA基因、2个rRNA基因(rrnL和rrnS)以及1个A+T富含区域。其中,9个蛋白质编码基因以及14个tRNA基因编码在N链上,其余14个基因编码在J链上。其线粒体基因组在25个不同的位置存在长度约1~66 bp的基因间间隔区,最长间隔序列位于trnE和trnF基因之间,其中有6对基因为重叠状态,重叠长度在1~7 bp,5对基因相邻,包括trnV-rrnS (图1和表2)。
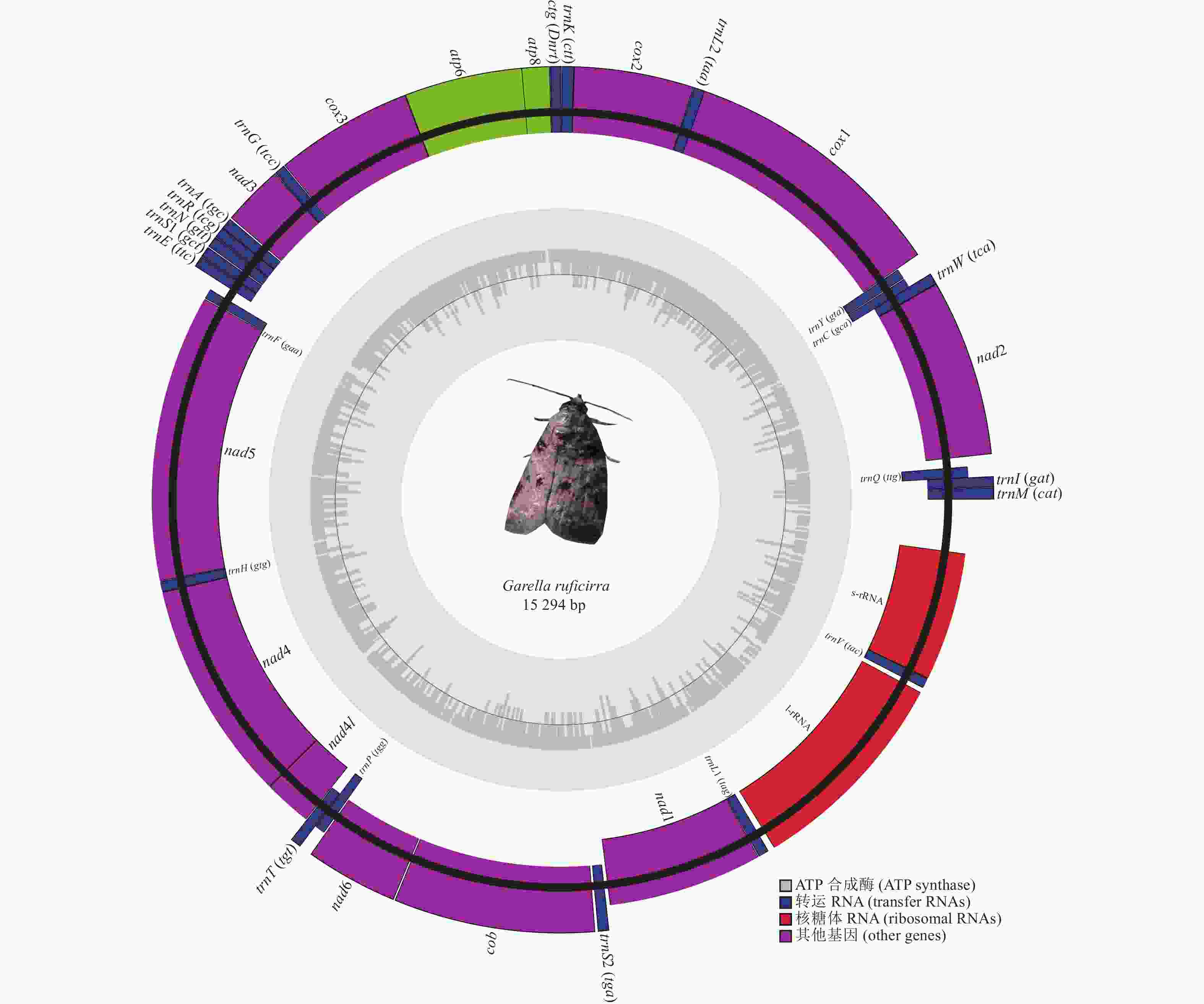
图 1 暗影饰皮夜蛾线粒体基因组结构
Figure 1. Structure of the mitochondrial genome of G. ruficirra
表 2 暗影饰皮夜蛾线粒体基因组组成
Table 2. Organization of the mitochondrial genome of G. ruficirra
基因 编码链 位置 长度/
bp起始密
码子终止密
码子反密
码子基因间隔
核苷酸基因 编码链 位置 长度/
bp起始密
码子终止密
码子反密
码子基因间隔
核苷酸trnM N 1~68 68 CAT trnN N 6 062~6 128 67 GTT 3 trnI N 69~133 65 GAT −3 trnS1 N 6 132~6 197 66 GCT trnQ J 131~199 69 TTG 57 trnE N 6 198~6 263 66 TTC 65 nad2 N 257~1 270 1 014 ATT TAA −2 trnF J 6 329~6 392 64 GAA 2 trnW N 1 269~1 336 68 TCA −8 nad5 J 6 395~8 137 1 743 ATT TAA trnC J 1 329~1 391 63 GCA 2 trnH J 8 138~8 203 66 GTG −1 trnY J 1 394~1 459 66 GTA 2 nad4 J 8 203~9 543 1 341 ATG TAA −1 cox1 N 1 462~3 000 1 539 ATG TAA −5 nad4l J 9 543~9 836 294 ATG TAA 2 trnL2 N 2 996~3 062 67 TAA trnT N 9 839~9 904 66 TGT cox2 N 3 063~3 747 685 ATG T(AA) −3 trnP J 9 905~9 969 65 TGG 6 trnK N 3 745~3 815 71 CTT nad6 N 9 976~10 509 534 ATT TAA 6 trnD N 3 816~3 882 67 GTC cob N 10 516~11 667 1152 ATG TAA 13 atp8 N 3 883~4 044 162 ATT TAA −7 trnS2 N 11 681~11 746 66 TGA 19 atp6 N 4 038~4 715 678 ATG TAA −1 nad1 J 11 766~12 704 939 ATG TAA 1 cox3 N 4 715~5 503 789 ATG TAA 2 trnL1 J 12 706~12 773 68 TAG 36 trnG N 5 506~5 570 65 TCC rrnL J 12 810~14 099 1290 27 nad3 N 5 571~5 924 354 ATC TAA 6 trnV J 14 127~14 191 65 TAG −1 trnA N 5 931~5 998 68 TGC −1 rrnS J 14 191~14 972 782 −13 trnR N 5 998~6 061 64 TCG -
暗影饰皮夜蛾线粒体基因组A、T、C、G含量分别为39.02%、41.51%、11.55%和7.92%,线粒体基因组的A+T含量为80.53%,表现为A、T碱基偏向性(表3),蛋白质编码基因的A+T含量在71.42% (cox1)~90.74% (atp8)范围内[22]。
表 3 线粒体基因组核苷酸组成
Table 3. Nucleotide composition of the complete mitochondrial genome
基因序列 碳基含量/% AT偏斜 GC偏斜 A C G T A+T G+C 全基因组 39.02 11.55 7.92 41.51 80.53 19.47 −0.0309 −0.1867 蛋白质编码基因 33.38 9.95 11.15 45.52 78.90 21.10 −0.1538 0.0566 密码子第1位 36.13 9.49 16.78 37.60 73.73 26.27 −0.0199 0.2777 密码子第2位 22.08 16.17 13.42 48.33 70.41 29.59 −0.3728 −0.0930 密码子第3位 41.94 4.20 3.23 50.63 92.57 7.43 −0.0938 −0.1295 tRNA基因 42.33 7.53 11.23 38.90 81.23 18.77 0.0422 0.1971 rRNA基因 44.16 4.97 10.14 40.73 84.89 15.11 0.0404 0.3419 控制区 47.14 3.70 1.01 48.15 95.29 4.71 −0.0106 −0.5714 整个核苷酸链显示轻微的AT偏斜和中度的GC偏斜(表3和图2)。在鳞翅目其他昆虫的线粒体基因组中同样存在AT偏斜和GC偏斜的情况[25−28]。暗影饰皮夜蛾的蛋白质编码基因和密码子第1位的AT偏斜为负,GC偏斜为正,密码子第2位和第3位产生的AT偏斜与GC偏斜均为负,tRNA和rRNA产生的AT偏斜和GC偏斜全部为正,蛋白质编码基因中T和G的占比高于A和C。
图 2 编码蛋白质各氨基酸的百分含量
Figure 2. Percentage of each amino acid of proteins coded
-
在暗影饰皮夜蛾线粒体基因组编码的13个蛋白质基因中,有9个基因编码在N链上,另外4个基因编码在J链上(图2),蛋白质编码基因总长为11 224 bp,占全基因组的73.39% (表2)。
暗影饰皮夜蛾13个蛋白质编码基因都是以ATN作为起始密码子,多数都是以ATG作为起始密码子,nad3以ATC作为起始密码子,而nad2、atp8、nad5、nad6以ATT作为起始密码子(表2)。大多数蛋白质编码基因都是以TAA作为终止密码子,只有cox2以不完整的T作为终止密码子,这与多数昆虫线粒体基因组以TAA或TAG作为终止密码子的情况相同[29]。
对暗影饰皮夜蛾蛋白质编码基因的氨基酸使用情况和相对同义密码子使用度(RSCU)的分析结果(表4)表明:除终止密码子外,暗影饰皮夜蛾线粒体基因组共有3 745个密码子,与多数昆虫线粒体编码出来的蛋白质密码子在3 585~3 746的范围一致。线粒体基因组中使用频率最高的4个密码子分别是UUU (苯丙氨酸Phe)、UUA (亮氨酸Leu)、UAU (酪氨酸Tyr)、AUA (异亮氨酸Ile),这4个密码子在线粒体基因组中的使用次数分别为488、474、315、304次,因此,蛋白质编码基因中编码最频繁的氨基酸依次为Leu (17.0%)、Ile (16.3%)、Phe (11.0%)、Tyr (8.0%)。使用最少的氨基酸是色氨酸Trp (0.52%),以C和G为结尾的密码子出现频率较低,以U和A为结尾的密码子通常RSCU大于1,说明U和A为基因组密码子中的偏爱碱基,C和G为基因组密码子中的非偏爱碱基。
表 4 氨基酸使用频率和相对同义密码子使用度
Table 4. Usage of amino acids and the relative synonymous codon usage
氨基酸 密码子 使用次数/次 RSCU 氨基酸 密码子 使用次数/次 RSCU 丙氨酸(Ala) GCU
GCC
GCA
GCG48
5
24
122.16
0.22
1.08
0.54脯氨酸(Pro) CCG
CCA
CCU
CCC9
24
38
160.41
1.10
1.75
0.74半胱氨酸(Cys) UGU
UGC39
131.50
0.50谷氨酰胺(Gln) CAG
CAA24
700.51
1.49天冬氨酸(Asp) GAU
GAC96
151.73
0.27精氨酸(Arg) CGU
CGC
CGA
CGG10
0
12
50.47
0.00
0.56
0.23谷氨酸(Glu) GAG
GAA38
760.67
1.33苯丙氨酸(Phe) UUU
UUC488
971.67
0.33酪氨酸(Tyr) UAU
UAC315
381.78
0.22甘氨酸(Gly) GGG
GGA
GGU
GGC36
56
48
21.01
1.58
1.35
0.06丝氨酸(Ser) UCU
UCC
UCA
UCG77
27
66
171.91
0.67
1.64
0.42甲硫氨酸(Met) AUG 68 1.00 色氨酸(Trp) UGG 23 1.00 天冬氨酸(Asn) AAU
AAC295
331.80
0.20苏氨酸(Thr) ACG
ACA
ACU
ACC7
30
56
250.24
1.02
1.90
0.85赖氨酸(Lys) AAG
AAA61
2500.39
1.61亮氨酸(Leu) UUG
UUA
CUG
CUA
CUU
CUC77
474
28
53
97
220.62
3.79
0.22
0.42
0.77
0.18缬氨酸(Val) GUG
GUA
GUU
GUC18
70
75
120.41
1.60
1.71
0.27组氨酸(His) CAU
CAC72
111.73
0.27异亮氨酸(Ile) AUA
AUU
AUC304
373
421.27
1.56
0.18终止(End) UGA
UAG
UAA77
82
2850.52
0.55
1.93 -
暗影饰皮夜蛾线粒体基因组的22个tRNA基因总长度为1 458 bp (表2),在暗影饰皮夜蛾线粒体全基因组中占比为9.53%,在N链上编码14个tRNA基因,J链上编码8个tRNA基因。暗影饰皮夜蛾线粒体基因组的22个tRNA基因个体长度分布在63 bp (trnC)~71 bp (trnK)范围。rrnL分布在trnL和trnV之间、J链上的rrnS则分布在trnV和线粒体控制区(CR)之间。rrnL长度为1 290 bp,rrnS长度为782 bp,rrnL的A+T含量为84.81%,rrnS的A+T含量为85.04%。从表3可以看出:AT偏斜和GC偏斜都表现为正数,说明暗影饰皮夜蛾在A和G碱基具有偏向性。
通过
http://mitos.bioinf.uni-leipzig.de/ 网址分析暗影饰皮夜蛾tRNA基因的二级结构模型,其中21个tRNA均具有典型的三叶草形态,只有trnS1的DHU臂被环结构取代,无法形成三叶草形态(图3)。暗影饰皮夜蛾的tRNA二级结构中,有22对碱基出现了错配现象,在普遍的U-G错配中,氨基酸接受臂上存在4对,TΨC臂上存在2对,反密码子臂存在4对,DHU臂上存在10对,其余2对错配分别为DHU臂上存在1对A-A错配,TΨC臂上存在1对A-A错配。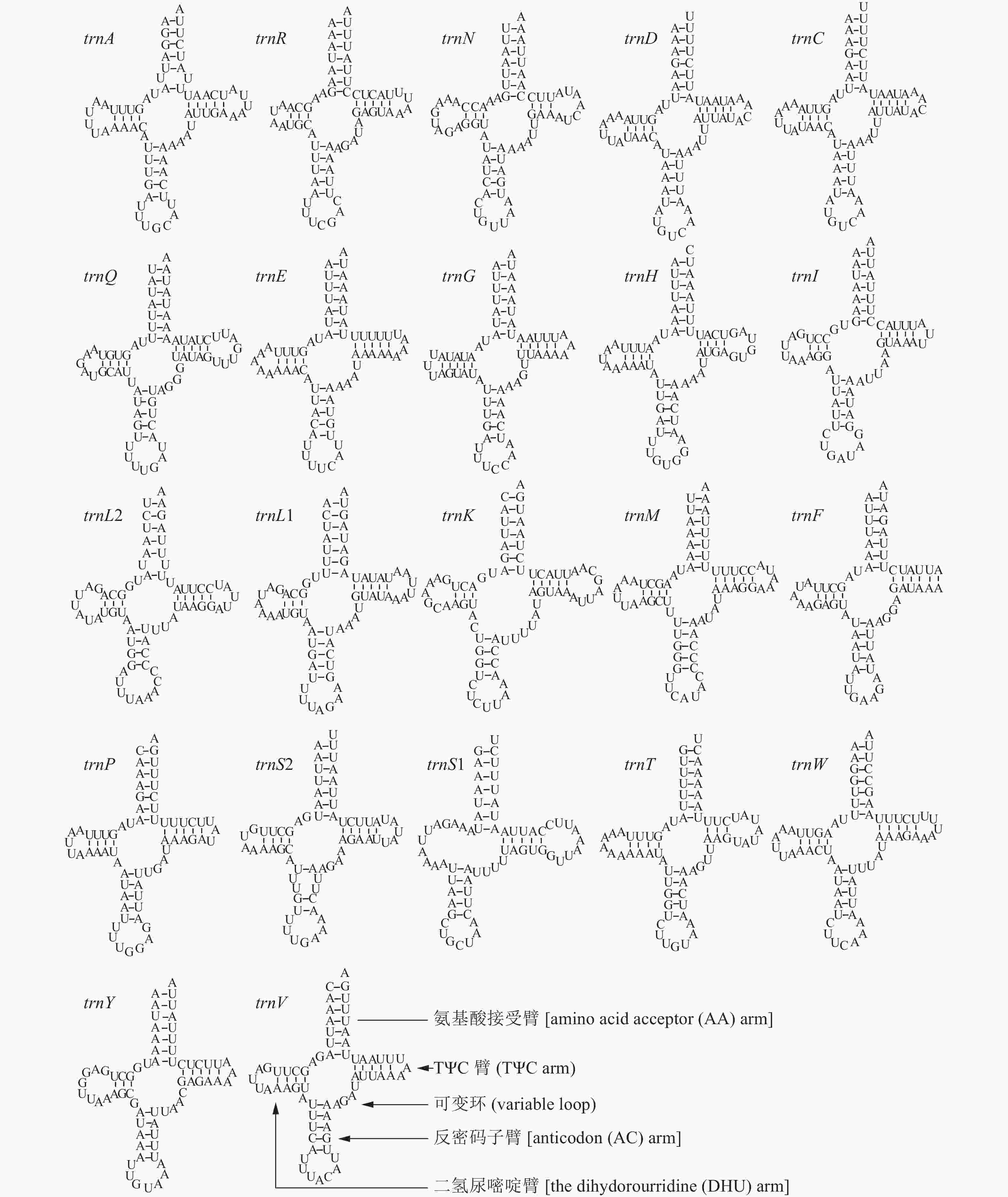
图 3 tRNA基因二级结构比较
Figure 3. Comparison on the secondary structure of tRNA genes
-
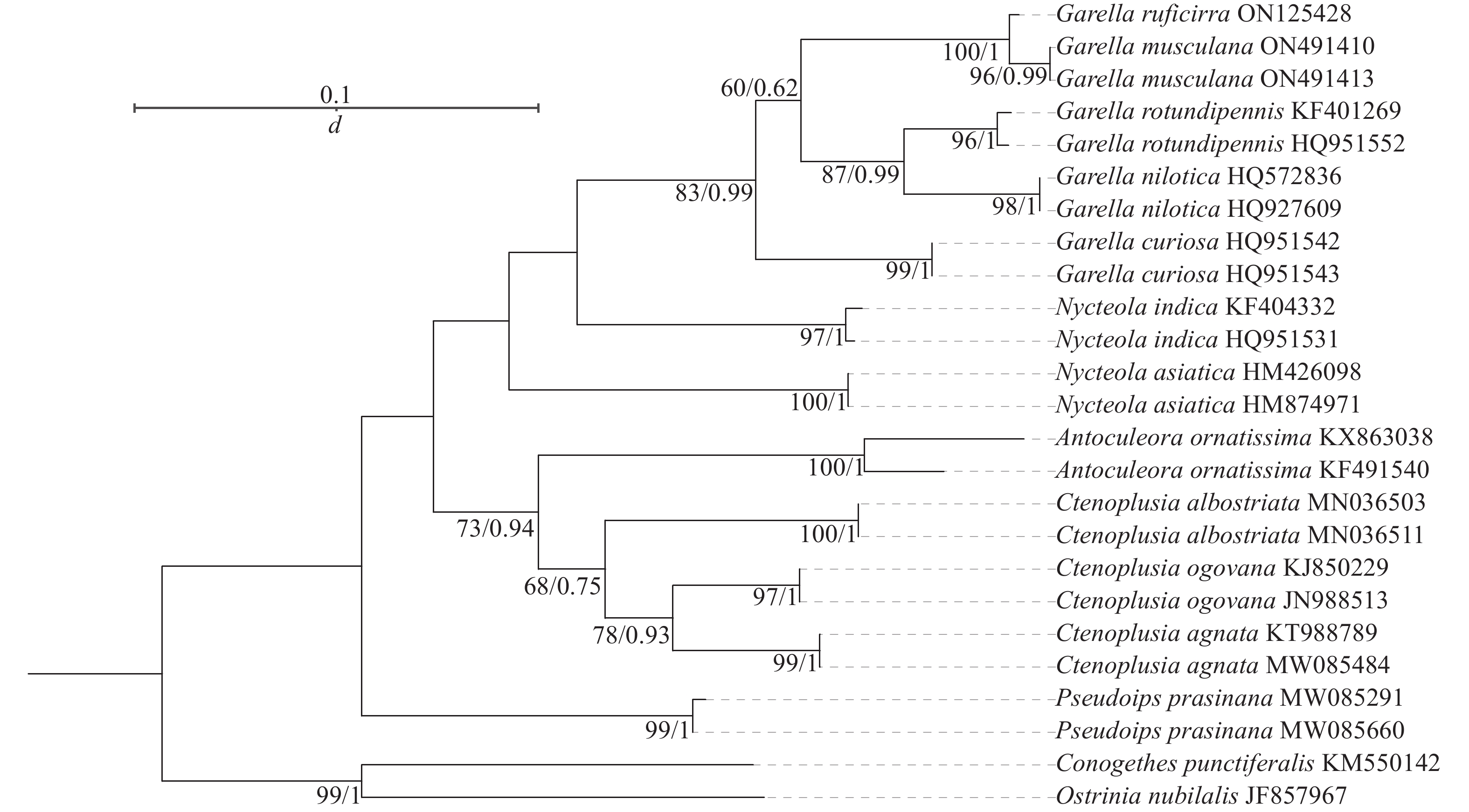
系统发育树结果(图4)表明:5个夜蛾属Garella与皮夜蛾属Nycteola亲缘关系最近,饰纹夜蛾属Antoculeora与Ctenoplusia属亲缘关系最近,与Garella和Nycteola稍远,这4种夜蛾与饰夜蛾属Pseudoips亲缘关系最远。Garella属的5种夜蛾之间,暗影饰皮夜蛾与G. musculana亲缘关系最近,G. rotundipennis与G. nilotica亲缘关系最近,与暗影饰皮夜蛾与G. musculana稍远,这4种夜蛾与G. curiosa亲缘关系最远。
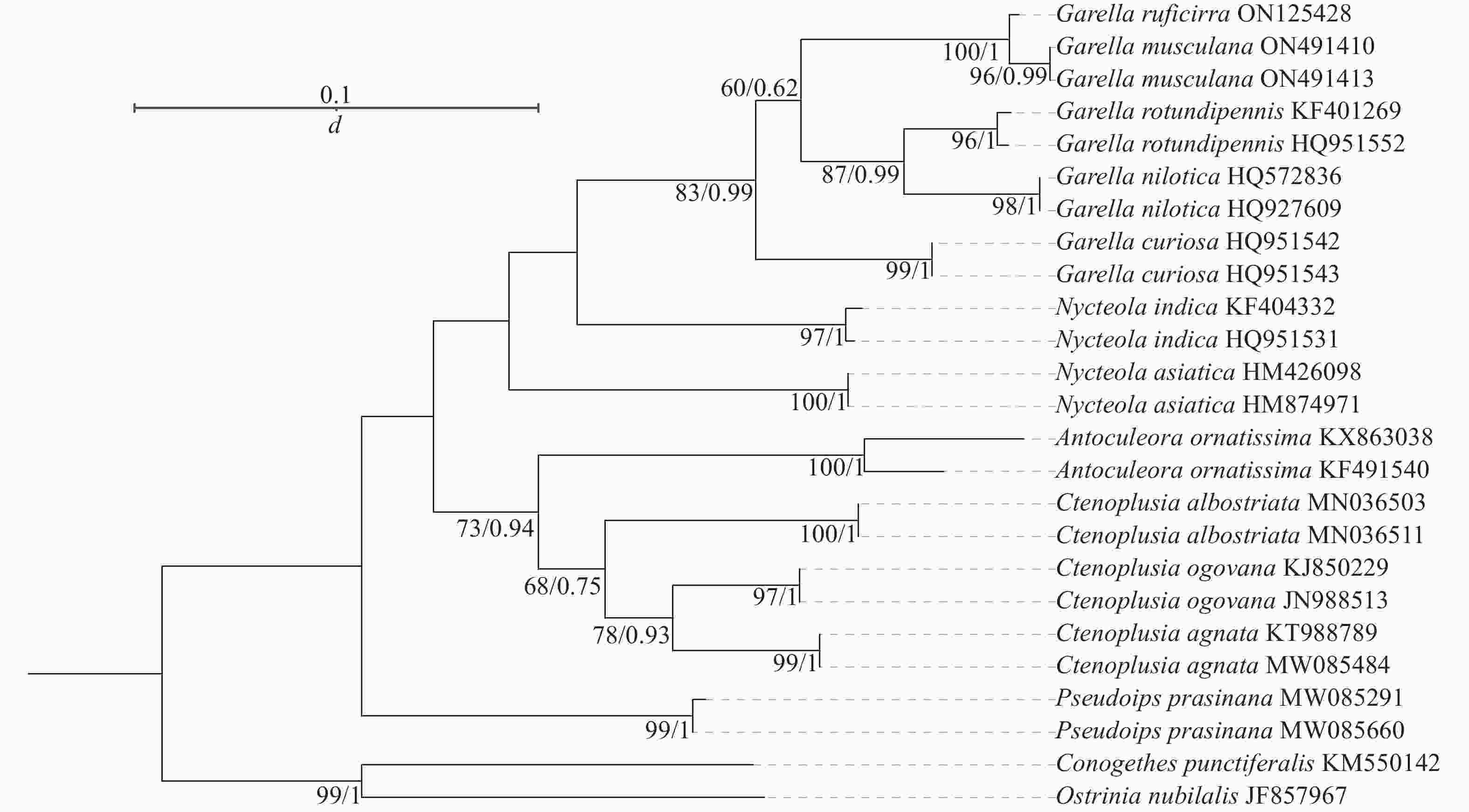
图 4 夜蛾科昆虫线粒体基因组蛋白质编码基因序列的系统发育树
Figure 4. Phylogenetic tree of based on protein-coding genes sequences of Noctuidae species
-
20世纪80年代暗影饰皮夜蛾在河南、山东等省已有发现,分布范围逐步扩大,在多个地区已成为板栗的主要害虫[30−32],近年来在薄壳山核桃主栽区逐年持续暴发。本研究表明:暗影饰皮夜蛾线粒体基因组长度为15 294 bp,在鳞翅目昆虫线粒体基因组报道的长度范围内[33]。其线粒体基因组所有区域表现出AT偏向性,与网纹蟒Reticulated python、绒螨目Trombidiformes螨类、对虾科Penaeidae物种相同[34−36]。而trnM-trnI-trnQ的基因排列方式在鳞翅目多个物种中均有存在[37−40],这种重排方式十分典型[41−43]。暗影饰皮夜蛾线粒体基因组所有蛋白质编码基因均以ATN为起始密码子,这与大多数鳞翅目昆虫一致[29]。暗影饰皮夜蛾cox1的起始密码子均为ATG,ATG是最常用的起始密码子,这与多数夜蛾科昆虫一致,与果蝇Drosophila melanogaster的起始密码子(ATT)不同[44−45]。除cox2以不完整的T为终止密码子外,其余13个蛋白质编码基因的终止密码子都是TAA,蛋白质编码基因中使用不完全终止密码子在无脊椎动物中十分常见[46−47]。其线粒体基因组有21个tRNA为典型的三叶草构造,只有tRNASer(AGN)缺少DHU臂,这在多种昆虫中均有出现,是夜蛾科的普遍特征[25−26]。
夜蛾科是鳞翅目中最大的一个科,而目前对夜蛾科线粒体基因组的研究种类较少,研究内容不全面,尤其是Garella属,目前除暗影饰皮夜蛾以外,其他物种均未进行线粒体基因组测定,因此需要补充大量夜蛾科各属昆虫线粒体全基因组序列的信息,为进一步探讨夜蛾科各属之间的系统发育关系提供基础数据。
-
暗影饰皮夜蛾的线粒体全基因组的碱基含量为T>A>C>G,表现为AT偏向性,基因组排列顺序为trnM-trnI-trnQ,存在基因重排现象。暗影饰皮夜蛾线粒体基因组的二级结构为典型的三叶草构造。系统发育分析表明:暗影饰皮夜蛾和G. musculana的亲缘关系最近。
Sequencing and analysis of the complete mitochondrial genome of Garella ruficirra
-
摘要:
目的 对薄壳山核桃Carya illinoensis害虫暗影饰皮夜蛾Garella ruficirra线粒体基因组进行测序和分析,并在基因组水平上探讨其在夜蛾科Noctuidae中的分类地位,为探索夜蛾科昆虫的系统发育关系以及演化进程提供参考。 方法 利用二代测序技术从头组装获取暗影饰皮夜蛾的线粒体基因组,并对线粒体基因组结构特点和碱基组成进行分析;同时,采用最大似然法和贝叶斯法联合构建了夜蛾科5个属、12个种的线粒体基因组系统发育树,分析暗影饰皮夜蛾在夜蛾科中的系统发育地位。 结果 暗影饰皮夜蛾线粒体基因组全长共为15 294 bp,其中包括13个蛋白质编码基因、22个转运RNA基因、2个核糖体RNA基因以及鳞翅目Lepidoptera昆虫典型的腺嘌呤(A)+胸腺嘧啶(T),即A+T富含区,该区域的A+T含量为80.53%,具有明显的AT偏向性。暗影饰皮夜蛾的基因排列顺序为trnM-trnI-trnQ,与包括夜蛾科昆虫在内的大多数鳞翅目昆虫基因排列次序相符。13个蛋白质编码基因的起始密码子全部为ATN。22个tRNA基因中除trnS1的DHU臂缺失,其余均为典型的三叶草结构。对线粒体基因组研究发现:夜蛾科5个属之间,Garella与皮夜蛾属Nycteola亲缘关系最近,与饰夜蛾属Pseudoips亲缘关系最远。 结论 暗影饰皮夜蛾的线粒体基因组中出现了基因重排的现象,系统发育关系支持暗影饰皮夜蛾和Garella musculana聚为1个分支。图4表4参47 Abstract:Objective This study, with a comprehensive sequencing and analysis of the mitochondrial genome of Garella ruficirra as well as an investigation into the phylogenetic position of Noctuidae through mitochondrial genome sequencing, is aimed to offer a valuable reference for delving into the phylogenetic relationships and evolutionary processes within the Noctuidae as well as exploring the phylogenetic relationship and evolutionary process of Noctuidae. Method First, the Illumina sequencing technique was employed to sequence the complete mitochondrial genome of G. ruficirra, allowing for the analysis of its overall characteristics and base composition. Subsequently, a phylogenetic tree encompassing mitochondrial genomes from 5 genera of Noctuidae, comprising 12 species, was constructed using both maximum likelihood and Bayesian methods so as to facilitate the examination of the phylogenetic relationships within the Noctuidae. Result The circular mitochondrial genome of G. ruficirra spanned 15 294 base pairs and encompasses 13 protein synthesis-related genes, 22 transfer RNA genes, and two ribosomal RNA genes and its gene arrangement conformed to the standard configuration observed in Lepidoptera mitogenomes, featuring a region rich in adenine and thymine, an A+T content of 80.53% and a significant AT bias. The gene order of trnM-trnI-trnQ in G. ruficirra aligned with other Noctuidae species and the majority of Lepidoptera with ATN sequences serving as initiation codons for all 13 protein-coding genes. While trnS1 lacked a DHU arm, the remaining tRNA genes exhibited a typical cloverleaf-shaped structure. According to the study of mitochondrial genome, Garella was most closely related to Nycteola and least closely related to Pseudoips among the 5 genera. Conclusion Gene rearrangement was evident in the mitochondrial genome of G. ruficirra. Phylogenetic analyses, indicating that the clustering of G. ruficirra and G. musculana samples was within a single clade. This study has furnished valuable insights into the evolutionary patterns of Garella and offered a foundational framework for investigating the phylogeny and evolution of Noctuidae within the Lepidoptera order. [Ch, 4 fig. 4 tab. 47 ref.] -
Key words:
- Garella ruficirra /
- mitochondrial genome /
- phylogenetic relationship /
- gene rearrangement
-
图 4 夜蛾科昆虫线粒体基因组蛋白质编码基因序列的系统发育树
拉丁名所代表的物种名见表1。
Figure 4 Phylogenetic tree of based on protein-coding genes sequences of Noctuidae species
表 1 用于线粒体基因组分析的昆虫物种信息
Table 1. Information of the insect species for phylogenetic analysis in mitochondrial genome
类群 总科 属 物种 内群 夜蛾科Noctuidae Garella 暗影饰皮夜蛾G. ruficirra G. musculana G. rotundipennis G. nilotica G. curiosa 皮夜蛾属Nycteola N. indica 亚皮夜蛾N. asiatica 饰纹夜蛾属Antoculeora 饰银纹夜蛾A. ornatissima Ctenoplusia 白条夜蛾C. albostriata C. ogovana 银纹夜蛾C. agnata 饰夜蛾属Pseudoips P. prasinana 外群 天蛾科Sphingidae 蛀野螟属Conogethes 桃蛀螟C. punctiferalis 秆野螟属Ostrinia 玉米螟O. nubilalis  下载: 导出CSV
下载: 导出CSV
表 2 暗影饰皮夜蛾线粒体基因组组成
Table 2. Organization of the mitochondrial genome of G. ruficirra
基因 编码链 位置 长度/
bp起始密
码子终止密
码子反密
码子基因间隔
核苷酸基因 编码链 位置 长度/
bp起始密
码子终止密
码子反密
码子基因间隔
核苷酸trnM N 1~68 68 CAT trnN N 6 062~6 128 67 GTT 3 trnI N 69~133 65 GAT −3 trnS1 N 6 132~6 197 66 GCT trnQ J 131~199 69 TTG 57 trnE N 6 198~6 263 66 TTC 65 nad2 N 257~1 270 1 014 ATT TAA −2 trnF J 6 329~6 392 64 GAA 2 trnW N 1 269~1 336 68 TCA −8 nad5 J 6 395~8 137 1 743 ATT TAA trnC J 1 329~1 391 63 GCA 2 trnH J 8 138~8 203 66 GTG −1 trnY J 1 394~1 459 66 GTA 2 nad4 J 8 203~9 543 1 341 ATG TAA −1 cox1 N 1 462~3 000 1 539 ATG TAA −5 nad4l J 9 543~9 836 294 ATG TAA 2 trnL2 N 2 996~3 062 67 TAA trnT N 9 839~9 904 66 TGT cox2 N 3 063~3 747 685 ATG T(AA) −3 trnP J 9 905~9 969 65 TGG 6 trnK N 3 745~3 815 71 CTT nad6 N 9 976~10 509 534 ATT TAA 6 trnD N 3 816~3 882 67 GTC cob N 10 516~11 667 1152 ATG TAA 13 atp8 N 3 883~4 044 162 ATT TAA −7 trnS2 N 11 681~11 746 66 TGA 19 atp6 N 4 038~4 715 678 ATG TAA −1 nad1 J 11 766~12 704 939 ATG TAA 1 cox3 N 4 715~5 503 789 ATG TAA 2 trnL1 J 12 706~12 773 68 TAG 36 trnG N 5 506~5 570 65 TCC rrnL J 12 810~14 099 1290 27 nad3 N 5 571~5 924 354 ATC TAA 6 trnV J 14 127~14 191 65 TAG −1 trnA N 5 931~5 998 68 TGC −1 rrnS J 14 191~14 972 782 −13 trnR N 5 998~6 061 64 TCG
下载: 导出CSV
表 3 线粒体基因组核苷酸组成
Table 3. Nucleotide composition of the complete mitochondrial genome
基因序列 碳基含量/% AT偏斜 GC偏斜 A C G T A+T G+C 全基因组 39.02 11.55 7.92 41.51 80.53 19.47 −0.0309 −0.1867 蛋白质编码基因 33.38 9.95 11.15 45.52 78.90 21.10 −0.1538 0.0566 密码子第1位 36.13 9.49 16.78 37.60 73.73 26.27 −0.0199 0.2777 密码子第2位 22.08 16.17 13.42 48.33 70.41 29.59 −0.3728 −0.0930 密码子第3位 41.94 4.20 3.23 50.63 92.57 7.43 −0.0938 −0.1295 tRNA基因 42.33 7.53 11.23 38.90 81.23 18.77 0.0422 0.1971 rRNA基因 44.16 4.97 10.14 40.73 84.89 15.11 0.0404 0.3419 控制区 47.14 3.70 1.01 48.15 95.29 4.71 −0.0106 −0.5714
下载: 导出CSV
表 4 氨基酸使用频率和相对同义密码子使用度
Table 4. Usage of amino acids and the relative synonymous codon usage
氨基酸 密码子 使用次数/次 RSCU 氨基酸 密码子 使用次数/次 RSCU 丙氨酸(Ala) GCU
GCC
GCA
GCG48
5
24
122.16
0.22
1.08
0.54脯氨酸(Pro) CCG
CCA
CCU
CCC9
24
38
160.41
1.10
1.75
0.74半胱氨酸(Cys) UGU
UGC39
131.50
0.50谷氨酰胺(Gln) CAG
CAA24
700.51
1.49天冬氨酸(Asp) GAU
GAC96
151.73
0.27精氨酸(Arg) CGU
CGC
CGA
CGG10
0
12
50.47
0.00
0.56
0.23谷氨酸(Glu) GAG
GAA38
760.67
1.33苯丙氨酸(Phe) UUU
UUC488
971.67
0.33酪氨酸(Tyr) UAU
UAC315
381.78
0.22甘氨酸(Gly) GGG
GGA
GGU
GGC36
56
48
21.01
1.58
1.35
0.06丝氨酸(Ser) UCU
UCC
UCA
UCG77
27
66
171.91
0.67
1.64
0.42甲硫氨酸(Met) AUG 68 1.00 色氨酸(Trp) UGG 23 1.00 天冬氨酸(Asn) AAU
AAC295
331.80
0.20苏氨酸(Thr) ACG
ACA
ACU
ACC7
30
56
250.24
1.02
1.90
0.85赖氨酸(Lys) AAG
AAA61
2500.39
1.61亮氨酸(Leu) UUG
UUA
CUG
CUA
CUU
CUC77
474
28
53
97
220.62
3.79
0.22
0.42
0.77
0.18缬氨酸(Val) GUG
GUA
GUU
GUC18
70
75
120.41
1.60
1.71
0.27组氨酸(His) CAU
CAC72
111.73
0.27异亮氨酸(Ile) AUA
AUU
AUC304
373
421.27
1.56
0.18终止(End) UGA
UAG
UAA77
82
2850.52
0.55
1.93
下载: 导出CSV
-
[1] 孙照鑫, 冯红. 线粒体相关内质网膜对线粒体功能的影响[J]. 自然杂志, 2023, 45(2): 127 − 138. SUN Zhaoxin, FENG Hong. Effect of mitochondria-associated endoplasmic reticulum membranes on mitochondrial function [J]. Chinese Journal of Nature, 2023, 45(2): 127 − 138. [2] CAMERON S L. Insect mitochondrial genomics: implications for evolution and phylogeny [J]. Annual Review of Entomology, 2014, 59(1): 95 − 117. [3] 王丹阳, 王予彤, 于良斌, 等. 绿眼赛茧蜂线粒体基因组全序列测定和分析[J]. 昆虫学报, 2020, 63(8): 1028 − 1038. WANG Danyang, WANG Yutong, YU Liangbin, et al. Sequencing and analysis of the complete mitochondrial genome of Zele chlorophthalmus (Hymenoptera: Braconidae) [J]. Acta Entomologica Sinica, 2020, 63(8): 1028 − 1038. [4] 章士美. 中国经济昆虫志: 半翅目(一)[M]. 北京: 科学出版社, 1985. ZHANG Shimei. Economic Insects of China: Hemiptera (1) [M]. Beijing: Science Press, 1985. [5] 朱国渊, 张永科, 孔祥东, 等. 二斑素瓢虫线粒体基因组全序列测定和分析[J]. 南方农业学报, 2023, 54(5): 1293 − 1302. ZHU Guoyuan, ZHANG Yongke, KONG Xiangdong, et al. Sequencing and of the complete mitochondrial genome of Illeis bistigmosa [J]. Journal of Southern Agriculture, 2023, 54(5): 1293 − 1302. [6] 杨金宏, 谢满超, 文欣茹, 等. 茶网蝽线粒体基因组全序列测定及系统发育分析[J]. 茶叶科学, 2022, 42(6): 839 − 850. YANG Jinhong, XIE Manchao, WEN Xinru, et al. The complete mitochondrial genome sequence and phylogenetic analysis of the Stephanitis chinensis [J]. Journal of Tea Science, 2022, 42(6): 839 − 850. [7] 赵乐, 贺屹成, 李钢, 等. 蚜蝇科昆虫线粒体基因组特征及系统发育研究[J]. 生命科学, 2022, 34(11): 1421 − 1430. ZHAO Le, HE Yicheng, LI Gang, et al. Characterization and phylogenetic analysis of the complete mitochondrial genome Srphidae (Insecta: Diptera) [J]. Chinese Bulletin of Life Sciences, 2022, 34(11): 1421 − 1430. [8] 赵卓, 刘晨阳, 瓮青芬, 等. 茶六斑褐锦斑蛾Sorita pulchella线粒体基因组特征与系统发育分析[J]. 河南农业大学学报, 2023, 57(2): 277 − 287. ZHAO Zhuo, LIU Chenyang, WENG Qingfen, et al. Characterization of the mitochondrial genome and phylogenetic analysis of Sorita pulchella [J]. Journal of Henan Agricultural University, 2023, 57(2): 277 − 287. [9] 李兴艳, 梁文凯, 泽桑梓, 等. 松瘤小蠹线粒体基因组测序及分析[J]. 西南林业大学学报(自然科学), 2024, 44(1): 194 − 199. LI Xingyan, LIANG Wenkai, ZE Sangzi, et al. Sequencing and analysis of the mitochondrial genome of Orthotomicis erosus [J]. Journal of Southwest Forestry University (Natural Sciences), 2024, 44(1): 194 − 199. [10] 姚余江, 陈斌, 李廷景. 土蜂科线粒体基因组序列测定和分析[J]. 昆虫学报, 2023, 66(1): 99 − 107. YAO Yujiang, CHEN Bin, LI Tingjing. Sequencing and analysis of the mitochondrial genome of Scoliidae [J]. Acta Entomologica Sinica, 2023, 66(1): 99 − 107. [11] LAU A W Y, TAN L T H, AB M N S, et al. The chemistry of gut microbiome in health and diseases[J/OL]. Progress in Microbes & Molecular Biology, 2021, 4(1): a0000175[2024-01-20]. doi: 10.36877/pmmb.a0000175. [12] SINGH V, SOOD A K, SHASHANK P R. New record of pecan husk feeders, Garella ruficirra (Hampson) and Conogethes sp. from India [J]. Himachal Journal of Agricultural Research, 2016, 42(1): 75 − 76. [13] 杨霁虹, 付香斌, 童永强, 等. 豫南板栗林栗皮夜蛾的生物学特性和防治研究[J]. 信阳师范学院学报(自然科学版), 2006, 19(4): 419 − 421. YANG Jihong, FU Xiangbin, TONG Yongqiang, et al. Study on biological characteristics and control of Chinese chestnut Characoma ruficirra in southern Henan Province [J]. Journal of Xinyang Normal University (Natural Science Edition), 2006, 19(4): 419 − 421. [14] SINGH A P, RAMOLA G C. Garella ruficirra (Hampson, 1905)[Noctuoidea: Nolidae: Chloephorinae: Sarrothripini] infesting young shoots of Quercus lanata Smith, 1819 (Fabaceae) in Kumaon region of Uttarakhand [J]. Indian Forester, 2019, 146(2): 183 − 184. [15] 顾建强, 陈东辉, 徐奎源, 等. 薄壳山核桃林地昆虫物种多样性、功能多样性及其相互关系分析[J]. 浙江农林大学学报, 2015, 32(1): 116 − 122. GU Jianqiang, CHEN Donghui, XU Kuiyuan, et al. Insect species diversity, functional diversity, and their mutual relationship in a pecan stand [J]. Journal of Zhejiang A&F University, 2015, 32(1): 116 − 122. [16] 吴浙东, 王政懂, 邓玉英, 等. 浙中板栗皮夜蛾的发生及防治[J]. 中国南方果树, 2001, 30(5): 60 − 61. WU Zhedong, WANG Zhengdong, DENG Yuying, et al. Occurrence and control of Characama ruficrra of chestnut in central Zhejiang [J]. South China Fruits, 2001, 30(5): 60 − 61. [17] 郑瑞杰, 王德永. 辽宁省日本栗主要虫害及防治技术[J]. 农业科技与装备, 2010(8): 73 − 75. ZHENG Ruijie, WANG Deyong. Main pest and control techniques for Japanese chestnut in Liaoning Province [J]. Agricultural Science &Technology and Equipment, 2010(8): 73 − 75. [18] PEARSON W R. Rapid and sensitive sequence comparison with FASTP and FASTA [J]. Methods in Enzymology, 1990, 183: 63 − 98. [19] CHEN Shifu, ZHOU Yanqing, CHEN Yaru, et al. Fastp: an ultra-fast all-in-one FASTQ preprocessor [J]. Bioinformatics, 2018, 34(17): 844 − 847. [20] PRJIBELSKI A, ANTIPOV D, MELESHKO D, et al. Using SPAdes de novo assembler[J/OL]. Current Protocols in Bioinformatics, 2020, 70(1): e102[2024-01-20]. doi: 10.1002/cpbi.102. [21] COIL D, JOSPIN G, DARLING A E. A5-miseq: an updated pipeline to assemble microbial genomes from Illumina MiSeq data [J]. Bioinformatics, 2014, 31(4): 587 − 589. [22] 李妍, 李志红, 张威, 等. 闽鸠蝙蛾(鳞翅目: 蝙蝠蛾科)线粒体基因组全序列测定和分析[J]. 浙江大学学报(农业与生命科学版), 2023, 49(2): 179 − 190. LI Yan, LI Zhihong, ZHANG Wei, et al. Sequencing and analysis of the complete mitochondrial genome of Endoclita minanus (Lepidoptera: Hepialidae) [J]. Journal of Zhejiang University (Agriculture &Life Sciences), 2023, 49(2): 179 − 190. [23] ZIMIN A V, SALZBERG S L. The genome polishing tool POLCA makes fast and accurate corrections in genome assemblies[J/OL]. PLoS Computational Biology, 2020, 16(6): e1007981[2024-01-20]. doi:10.1101/2019.12.17.864991. [24] DRUMMOND A J, SUCHARD M A, XIE D, et al. Bayesian phylogenetics with BEAUti and the BEAST 1.7 [J]. Molecular Biology and Evolution, 2012, 29(8): 1969 − 1973. [25] CHAN P P, LIN B Y, MAK A J, et al. tRNAscan-SE 2.0: improved detection and functional classification of transfer RNA genes [J]. Nucleic Acids Research, 2021, 49(16): 9077 − 9096. [26] 江宏燕, 陈世春, 廖姝然, 等. 扁刺蛾线粒体基因组全序列特征及系统发育分析[J]. 茶叶科学, 2023, 43(4): 460 − 472. JIANG Hongyan, CHEN Shichun, LIAO Shuran, et al. The complete mitochondrial genome sequence and phylogenetic analysis of Thosea sinensis [J]. Journal of Tea Science, 2023, 43(4): 460 − 472. [27] 王瑶, 孔祥波, 张苏芳, 等. 云南松毛虫线粒体基因组全序列测定和分析[J]. 林业科学研究, 2019, 32(5): 11 − 20. WANG Yao, KONG Xiangbo, ZHANG Sufang, et al. Sequencing and analysis of complete mitochondrial genome of Dendrolimus houi Lajonquiere (Lepidopeta: Lasiocampidae) [J]. Forest Research, 2019, 32(5): 11 − 20. [28] 彭艳, 陈斌, 李廷景. 黄侧异腹胡蜂线粒体基因组全序列测定和分析[J]. 昆虫学报, 2017, 60(4): 464 − 474. PENG Yan, CHEN Bin, LI Tingjing. Sequencing and analysis of the complete mitochondrial genome of Parapolybia crocea (Hymenoptera: Vespidae) [J]. Acta Entomologica Sinica, 2017, 60(4): 464 − 474. [29] WOLSTENHOLME D R. Animal mitochondrial DNA: structure and evolution [J]. International Review of Cytology, 1992, 141: 173 − 216. [30] 刘金凤, 赵宾, 张保贵, 等. 伏牛山区板栗主要虫害的发生特点与防治方法[J]. 现代农业科技, 2012(20): 139 − 140. LIU Jinfeng, ZHAO Bin, ZHANG Baogui, et al. Occurrence characteristics and control methods of main insect pests in chestnut in Funiu mountain [J]. Modern Agricultural Science and Technology, 2012(20): 139 − 140. [31] 王明月. 丽水板栗主要害虫危害调查与噻虫啉防治技术研究[D]. 杭州: 浙江农林大学, 2012. WANG Mingyue. Investigation of the Pest Insects Damaged Chestnut in Lishui and Study on Control Techniques with Thiacloprid[D]. Hangzhou: Zhejiang A&F University, 2012. [32] 肖云丽, 汪玉平, 程水源, 等. 我国板栗害虫研究概述[J]. 环境昆虫学报, 2014, 36(3): 441 − 450. XIAO Yunli, WANG Yuping, CHENG Shuiyuan, et al. Review on pests of chestnut in China [J]. Journal of Environmental Entomolog, 2014, 36(3): 441 − 450. [33] 陈汕, 李粉红, 兰旭娥, 等. 三种斑野螟全线粒体基因组及螟蛾总科系统发育分析[J]. 应用昆虫学报, 2017, 54(1): 22 − 34. CHEN Shan, LI Fenhong, LAN Xu’ e, et al. Complete mitochondrial genomes of three Spilomelinae species and a preliminary phylogenetic analysis of the Pyraloidea (Insecta: Lepidoptera) [J]. Chinese Journal of Applied Entomology, 2017, 54(1): 22 − 34. [34] 白天, 何叶艳, 高旭龙, 等. 网纹蟒非入侵式采样和线粒体基因组分析[J]. 浙江农林大学学报, 2019, 36(3): 444 − 450. BAI Tian, HE Yeyan, GAO Xulong, et al. Non-invasive sampling and mitochondrial genome analysis in Python reticulatus [J]. Journal of Zhejiang A&F University, 2019, 36(3): 444 − 450. [35] 冯蕊, 方颖, 方瑜, 等. 绒螨目线粒体基因组研究进展[J]. 热带病与寄生虫学, 2022, 20(1): 43. FENG Rui, FANG Ying, FANG Yu, et al. Research progress in the complete mitochondrial genomes of the Trombidformes [J]. Journal of Tropical Diseases and Parasitology, 2022, 20(1): 43. [36] 朱雷宇, 朱志煌, 方民杰, 等. 对虾科物种线粒体基因组特征和系统发育分析[J]. 上海海洋大学学报, 2023, 32(2): 292 − 302. ZHU Leiyu, ZHU Zhihuang, FANG Minjie, et al. Characteristics and phylogenetic analysis of mitochondrial genome in the Penaeidae [J]. Journal of Shanghai Ocean University, 2023, 32(2): 292 − 302. [37] LI Xinyu, LIU Yuncan, ZHANG Rrusong, et al. The mitochondrial genome of Qinghuang_1, the first modern improved strain of Chinese oak silkworm, Antheraea pernyi (Lepidoptera: Saturniidae) [J]. Journal of Insects as Food and Feed, 2021, 7(2): 233 − 243. [38] HUANG Yufeng, BOZDOGAN H, CHEN T H, et al. The complete mitochondrial genome of Attacus atlas formosanus Villiard, 1969 (Lepidoptera: Saturniidae) [J]. Mitochondrial DNA Part B, 2022, 7(1): 219 − 221. [39] CHEN Dongbin, ZHANG Rusong, JIN Xiangdong, et al. First complete mitochondrial genome of Rhodinia species (Lepidoptera: Saturniidae): genome description and phylogenetic implication [J]. Bulletin of Entomological Research, 2022, 112(2): 243 − 252. [40] DAI Lishang, KAUSAR S, ABBAS M N, et al. Complete sequence and characterization of the Ectropis oblique mitochondrial genome and its phylogenetic implications [J]. International Journal of Biological Macromolecules, 2018, 107: 1142 − 1150. [41] BOORE J L, LAVROV D V, BROWN W M. Gene translocation links insects and crustaceans [J]. Nature, 1998, 392(6677): 667 − 668. [42] 王维, 孟智启, 石放雄, 等. 鳞翅目昆虫比较线粒体基因组学研究进展[J]. 科学通报, 2013, 58(30): 3017 − 3029. WANG Wei, MENG Zhiqi, SHI Fangxiong, et al. Advances in comparative studies of Lepidoptera (Arthropoda: Insecta) [J]. Chinese Science Bulletin, 2013, 58(30): 3017 − 3029. [43] 陈志腾, 杜予州. 昆虫线粒体基因组重排的研究进展[J]. 环境昆虫学报, 2016, 38(4): 843 − 851. CHEN Zhiteng, DU Yuzhou. Rearrangement of mitochondrial genome in insects [J]. Journal of Environmental Entomology, 2016, 38(4): 843 − 851. [44] 张方, 米志勇. 动物线粒体 DNA 的分子生物学研究进展[J]. 中国生物工程杂志, 1998, 18(3): 25 − 31, 6. ZHANG Fang, MI Zhiyong. Advances in molecular biology of animal mitochondrial DNA [J]. Progress in Biotechnology, 1998, 18(3): 25 − 31, 6. [45] 马婷婷, 林菲, 赵楠, 等. 入侵害虫甘薯凹胫跳甲的鉴定及线粒体基因组分析[J]. 昆虫学报, 2022, 65(10): 1354 − 1366. MA Tingting, LIN Fei, ZHAO Nan, et al. Identification and mitochondrial genome analysis of the sweetpotato flea beetle, Chaetocnema confifinis (Coleptera: Chrysomelidae), an invasive pest in the Chinese mainland [J]. Acta Entomologica Sinica, 2022, 65(10): 1354 − 1366. [46] ZHANG Wenqiang, LIN Long, DING Yuhui, et al. Comparative mitogenomics of jumping spiders with first complete mitochondrial genomes of Euophryini (Araneae: Salticidae)[J/OL]. Insects, 2023, 14(6): 517[2024-01-20]. doi: 10.3390/insects14060517. [47] WU Hua, CAO Li, HE Meiyu, et al. Interspecific hybridization and complete mitochondrial genome analysis of two ghost moth species[J/OL]. Insects, 2021, 12(11): 1046[2024-01-20]. doi: 10.3390/insects12111046. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20240138
 点击查看大图
点击查看大图
计量
- 文章访问数: 1868
- HTML全文浏览量: 848
- PDF下载量: 31
- 被引次数: 0
