






-
桂花Osmanthus fragrans为木犀科Oleaceae木犀属Osmanthus,是中国十大传统名花之一,也是园林造景常用植物。根据开花时间不同,桂花可以分为秋桂和四季桂;根据花色差异,秋桂又可以分为丹桂、金桂和银桂。已有研究分析了桂花不同花色品种呈色物质成分,证实类胡萝卜素的种类及其质量分数是决定桂花花色的最主要因素[1−2]。目前,桂花类胡萝卜素的定性定量及其代谢途径中相关催化酶基因已被陆续分离得到[3−5]。桂花不同花色品种花瓣所含的类胡萝卜素中,β-胡萝卜素相对含量最高[1]。桂花番茄红素β-环化酶OfLCYB具备使番茄红素两端环化转化为β-胡萝卜素的能力,且OfLCYB对番茄红素的底物亲和性强于其他番茄红素环化酶,是桂花类胡萝卜素代谢途径中的关键催化酶[6−7]。沈子又等[8]分离得到了OfLCYB基因启动子,发现其启动子序列均包含有TATA-box、CAAT-box响应元件及水杨酸、赤霉素、脱落酸等激素响应元件等,但目前有关桂花OfLCYB基因上游转录因子的筛选及鉴定鲜见报道。
已有研究认为:ERF[9]、MYB[10]、NAC[11]等转录因子参与调控植物类胡萝卜素代谢。AP2/ERF转录因子家族具有众多的家族成员。根据AP2/ERF结构域的数目和序列特征,AP2/ERF家族转录因子分为AP2、ERF、CBF/DREB、RAV和Soloist这5个亚组,其中ERF类转录因子仅含有1个AP2/ERF结构域。ERF转录因子通过结合下游靶基因的GCC (GCCGCC)或DRE (CCGAC)序列[12]调节基因的表达,参与调节植物生长发育、生物或非生物胁迫应答、调控果实成熟等。此外,在拟南芥Arabidopsis thaliana[9]、番茄Solanum lycopersicum[13]和苹果Malus domestica[14]中还发现B2亚组的ERF转录因子具有调控植物类胡萝卜素合成的功能。拟南芥B2亚组ERF转录因子包括At3g16770.1(AtERF72/AtRAP2.3)、At1g72360.2 (AtERF73)、At1g53910.1 (AtERF74/AtRAP2.2)等5个成员。AtRAP2.2蛋白可以结合到拟南芥AtPSY启动子和AtPDS启动子的ATCTA元件上,从而调控相关基因的表达[15]。在苹果MdPSY1和MdPSY2基因启动子中也存在多个ATCTA顺式作用元件,能被AtRAP2.3的同源基因蛋白AP2D15强烈激活表达[14]。在黄龙胆Gentiana lutea[16]中,GlLCYB、GlLCYE、GlZEP、GlPDS、GlZDS、GlBCH基因的启动子上均存在ATCTA作用元件,说明ATCTA元件广泛存在于类胡萝卜素合成基因启动子上,表明B2亚组的ERF转录因子可能对一系列类胡萝卜素代谢基因具有调控作用。
本研究以桂花丹桂品种‘堰虹桂’O. fragrans ‘Yanhong Gui’为材料,首先对OfLCYB基因启动子的ATCTA顺式作用元件进行分析,再对桂花B2亚组的ERF转录因子基因进行序列分析和表达分析,利用酵母单杂交技术筛选和鉴定与OfLCYB互作的关键B2亚组的OfERF转录因子,不仅可以扩展桂花花色研究领域,同时为揭示桂花类胡萝卜素代谢的调控网络提供理论依据,为桂花品种培育和种质创新提供新的思路。
-
选择浙江农林大学桂花资源圃生长状况良好的地栽桂花品种‘堰虹桂’为材料,分别采集‘堰虹桂’的新鲜嫩叶以及顶壳期(S1)、铃梗期(S2)、初花期(S3)、盛开期(S4)的花瓣样品[17],每个样品3次生物学重复,取样时间均为10:00。上述叶片与花瓣样品快速采集后放入液氮冷冻,随后保存于−80 ℃超低温冰箱,供后续使用。
-
根据诺禾致源的Ultraclean plant DNA purification Kit试剂盒操作说明提取‘堰虹桂’的嫩叶鲜样DNA。借助PlantCARE数据库(
http://bioinformatics.psb.ugent.be/webtools/plantcare/html/ )分析启动子顺式作用元件。根据OfLCYB的启动子序列信息[8]设计引物,以‘堰虹桂’嫩叶DNA为模板扩增得到其启动子。以‘堰虹桂’不同时期的花瓣cDNA为模板,以OfLCYB基因序列设计表达引物,以桂花OfACT基因[18]为内参基因,按照TB Green® Premix Ex TM TapⅡ说明进行实时荧光定量聚合酶链式反应(RT-qPCR)分析。引物序列见表1。利用参照基因的2−ΔΔCt法计算目的基因的相对表达量。表 1 PCR引物序列
Table 1. PCR primer sequences
引物名称 引物序列(5′→3′) LCYB-PRO-GW-F ggggacaagtttgtacaaaaaagcaggcttcCTGCTTCTTGTTGTTGTACG LCYB-PRO-GW-R ggggaccactttgtacaagaaagctgggtcCAATTTTGGCATGTTCTTAG OfLCYB-qF GAAAGGAGACGCCAAAGGGAG OfLCYB-qR GGAAGAAATAGCCGAGATGATAAGA 说明:小写字母表示部分attB序列。 -
使用天根公司RNA perp Pure Plant Kit试剂盒,根据产品说明提取‘堰虹桂’不同时期的花瓣RNA。随后用紫外分光光度计和质量分数为1%琼脂糖凝胶电泳检测总RNA浓度和质量。按照PrimeScriptTM RT Master Mix说明书将检验合格的盛花期RNA进行反转录。
应用Prot-Param在线软件 (
http://web.expasy.org/protparam/ ) 预测所编码蛋白的分子量、理论等电点、不稳定系数等;采用MEGAX软件中的邻位相邻法(NJ)进行同源聚类,建立系统发育树,并采用Bootstrap法(重复1 000次) 评估检测系统进化树。运用DNAMAN 7.0对4个OfERF基因推测所得的序列进行多序列比对分析。 -
以‘堰虹桂’不同时期的花瓣cDNA为模板,以筛选得到的桂花B2亚组OfERFs序列设计引物,以桂花OfACT为内参基因。分析方法参照1.2.1。引物序列见表2。
表 2 OfERFs基因RT-qPCR引物序列
Table 2. RT-qPCR primer sequences of OfERFs
引物名称 引物序列(5′→3′) 引物名称 引物序列(5′→3′) OfERF73a-qF CTGAAGAGAAACCGCCAACAA OfERF72a-qR GGGTAGTAAACTTCTTGTTGCTGCGTA OfERF73a-qR TTAACGCCATCAGAAGACACAAGT OfERF72b-qF CAAATATCCTATGTTCAGAGG OfERF73b-qF AATTGGGATGCCGCCTCA OfERF72b-qR ATAGCATACCATAACATACCA OfERF73b-qR TTAAATCCCACCAAACATAGCACT OfACT-qF CCCAAGGCAAACAGAGAAAAAAT OfERF72a-qF CCAACCCCACCGGCTC OfACT-qR ACCCCATCACCAGAATCAAGAA -
通过Gateway方法构建pAbAi-OfLCYB-pro载体,之后利用限制性内切酶BstB I线性化质粒pAbAi-OfLCYB-pro、阳性对照p53-AbAi以及阴性对照pAbAi载体。按照Yeastmaker™ Yeast Transformation System 2 User Manual产品说明制备酵母感受态,并将线性化的质粒转入感受态细胞中,涂布于尿嘧啶缺陷培养基(SD/-Ura)酵母板筛选培养基上,28 ℃倒置培养2~3 d。挑取单菌落扩大培养,提取酵母DNA。以粗提酵母DNA为模板,进行PCR检验。用质量分数为0.9% 的无菌氯化钠溶液稀释菌液,D(600)=0.002时,均匀涂布于金担子素A (AbA)不同浓度的SD/-Ura固体培养基上,倒置于28 ℃培养箱内培养2~3 d,以检测AbAr基因本底表达水平。将pGADT7-OfERF72a、pGADT7-OfERF72b、pGADT7-OfERF73a、pGADT7-OfERF73b和pGADT7-53、pGADT7分别转入诱饵菌株pAbAi-OfLCYB-pro和阳性对照p53-AbAi、阴性对照pAbAi的酵母感受态细胞,悬浮液均匀涂布于SD/-Leu缺陷培养基上,倒置于30 ℃培养箱内培养3~5 d,再将长出的单菌落分别在亮氨酸缺陷培养基(SD/-Leu)与含300 μg·L−1的亮氨酸缺陷培养基[SD/-Leu/AbA(300 μg·L−1)]点斑检测其互作情况。
-
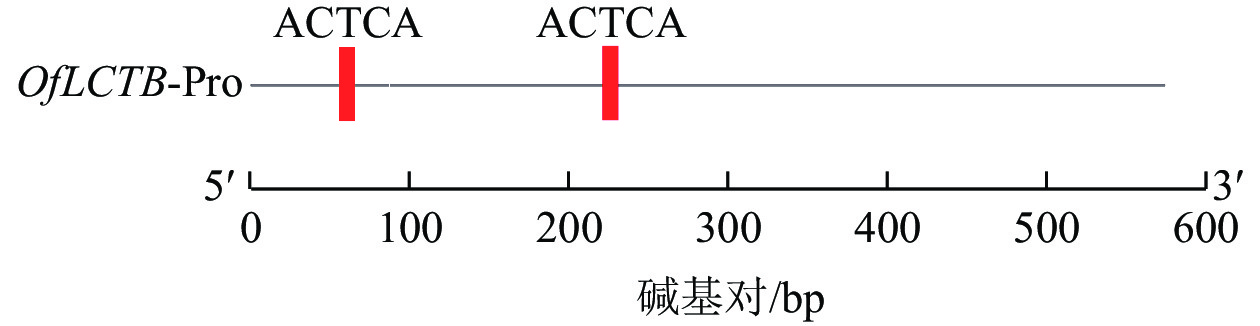
由图1可见:桂花OfLCYB基因启动子序列含有2个ATCTA顺式作用元件。
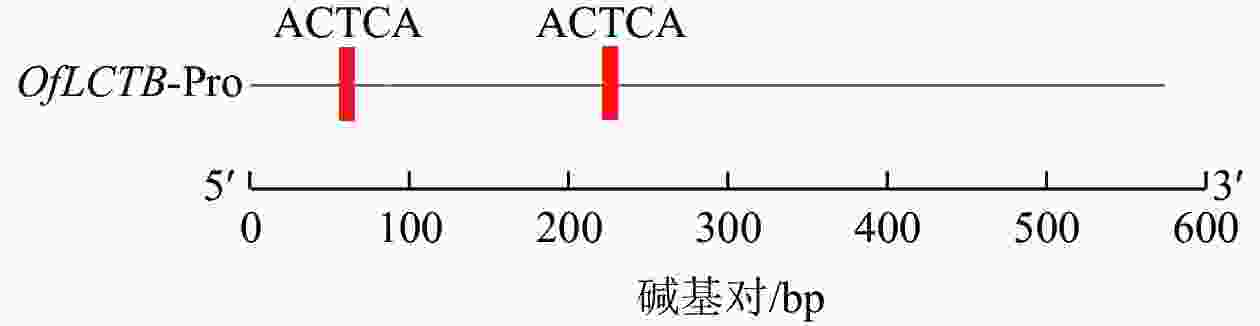
图 1 OfLCYB启动子的ATCTA顺式作用元件分析
Figure 1. Analysis of ATCTA cis-acting elements of the OfLCYB promoter
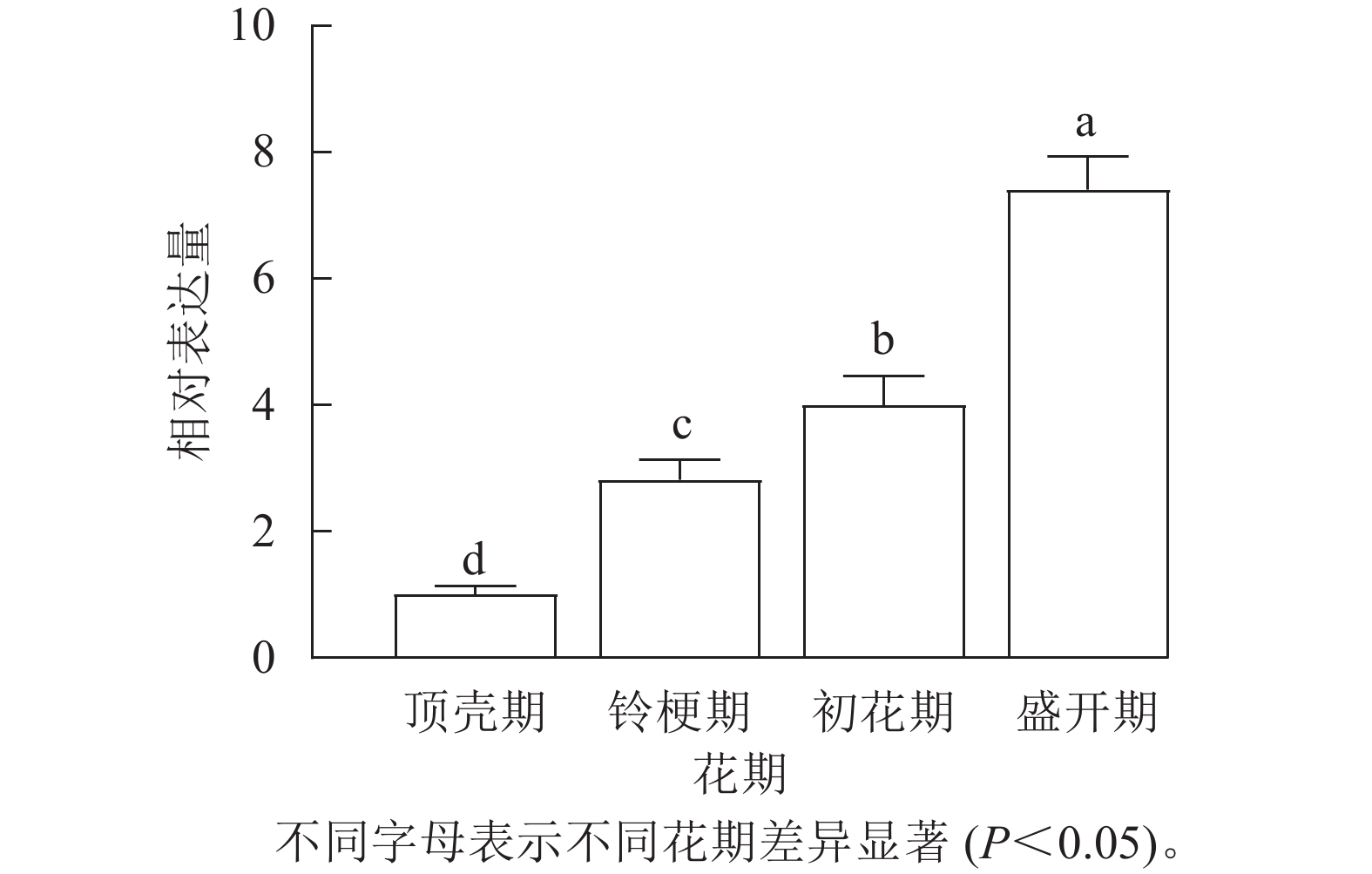
通过荧光定量检测‘堰虹桂’不同发育时期花瓣中OfLCYB的表达水平(图2),发现OfLCYB的表达量从顶壳期到盛开期逐渐升高,在盛开期表达量最高。
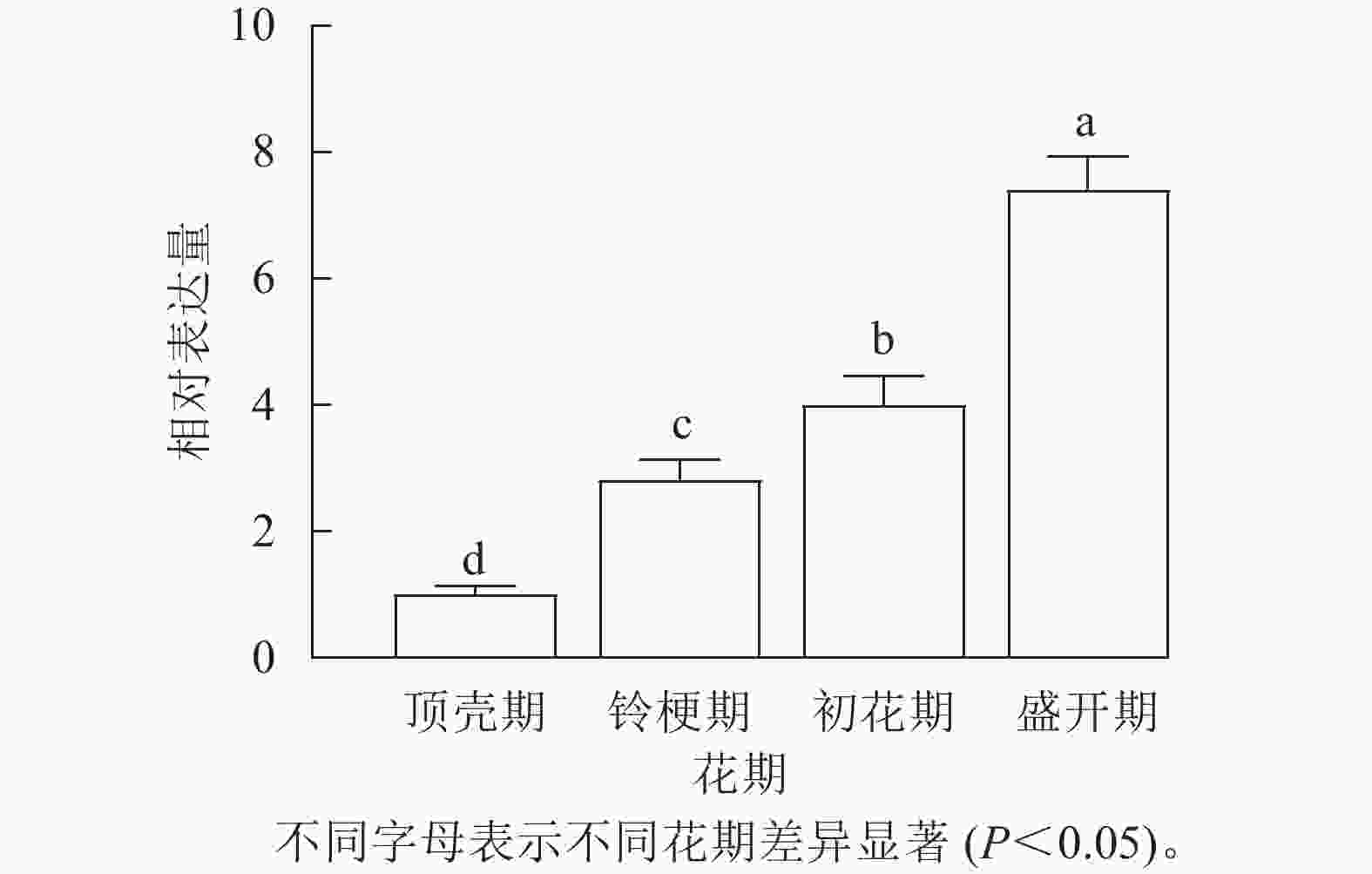
图 2 OfLCYB在‘堰虹桂’不同花期的表达
Figure 2. Expression of OfLCYB at different flowering stages in O. fragrans ‘Yanhong Gui’
-
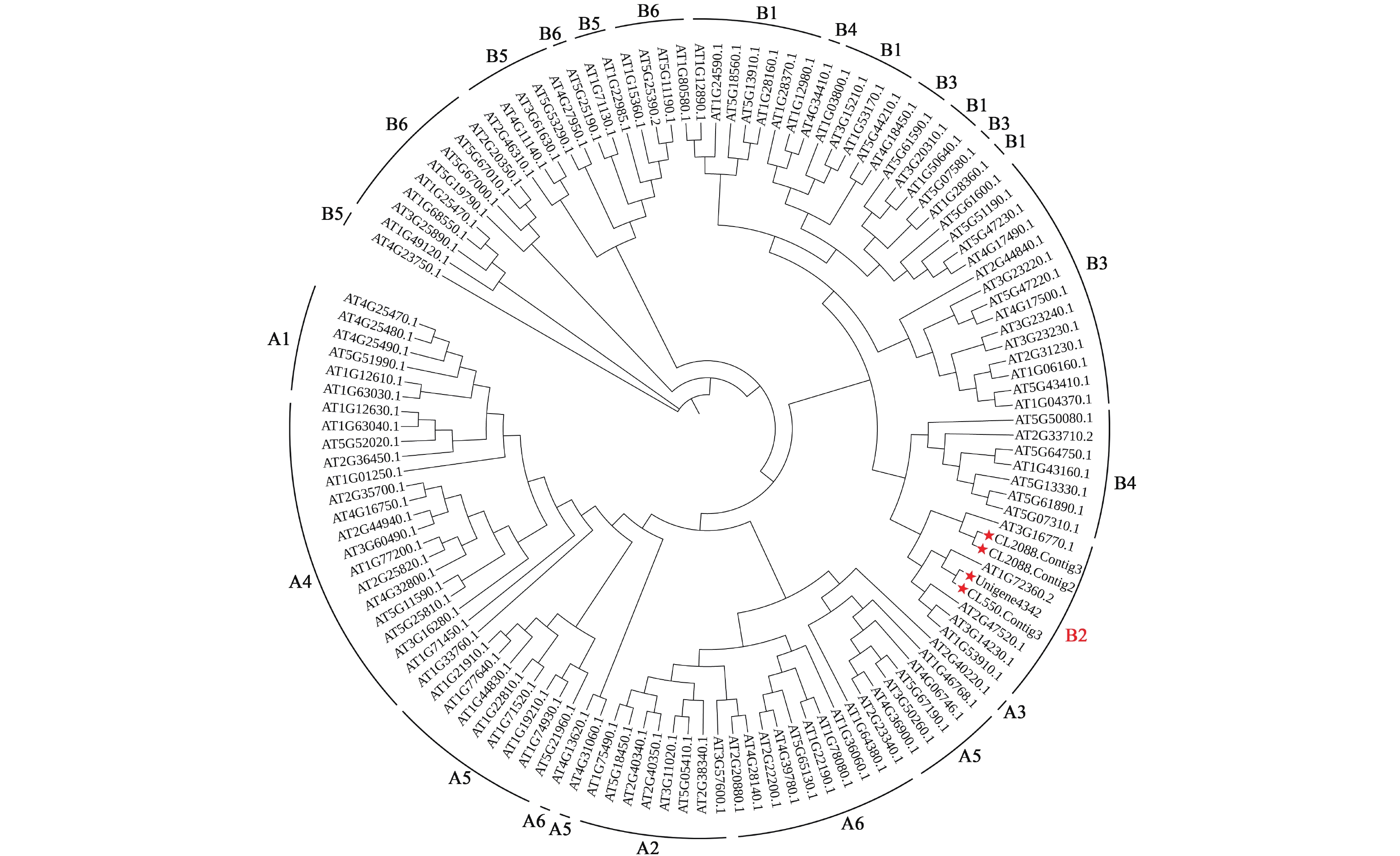
通过对桂花转录组数据库分析,筛选获得4个B2亚组ERF有关的Unigene序列。利用MEGAX软件对4个桂花OfERFs氨基酸全长和拟南芥ERF家族的122个成员的氨基酸序列构建系统进化树,结果显示:4个桂花OfERFs与5个拟南芥ERF序列聚集在B2亚组(图3)。其中CL2088.Contig2和CL2088.Contig3聚为一小支,与拟南芥At3g16770.1 (AtERF72)的关系最为接近,将CL2088.Contig2和CL2088.Contig3分别命名为OfERF72a和OfERF72b。此外,CL550.Contig3、Unigene4342与拟南芥At1g72360.2 (AtERF73)关系较近,将CL550.Contig3和Unigene4342分别命名为OfERF73a和OfERF73b。
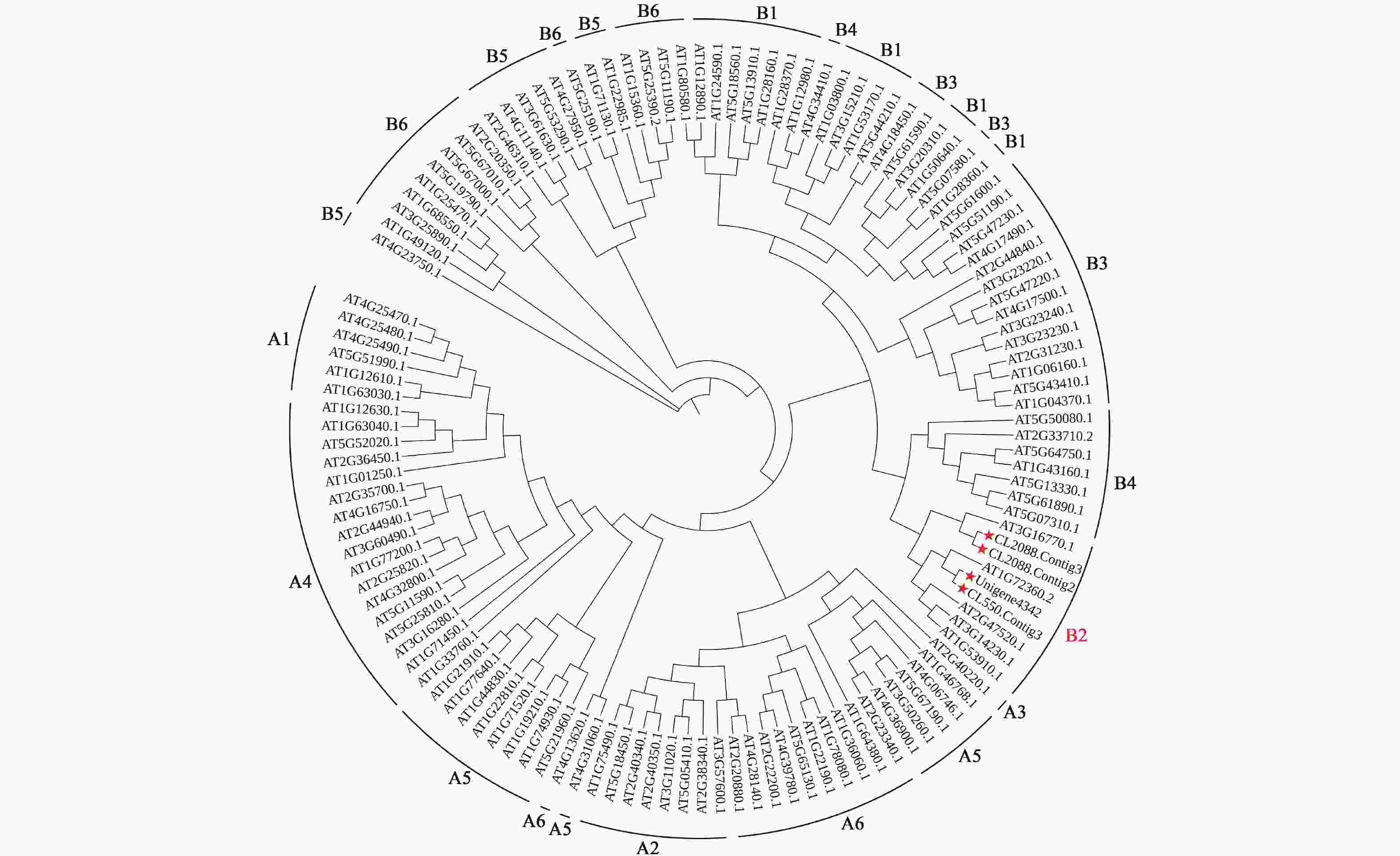
图 3 桂花B2亚组OfERFs系统发育分析
Figure 3. Phylogenetic analysis of OfERFs in subgroup B2 of O. fragrans
多序列比对分析发现(图4) :4个OfERF基因均包含1个AP2保守结构域。4个OfERFs蛋白序列的基本理化性质(表3)分析发现:OfERF72a基因的氨基酸数量为232个,分子量为26 144 Da;OfERF72b基因的氨基酸数量为228个,分子量为25 841 Da;OfERF73a基因的氨基酸数量为386个,分子量为43 632 Da;OfERF73b基因的氨基酸数量为375个,分子量为41 607 Da。4个OfERF的理论等电点为4.63~5.33,均属于偏酸性蛋白质;总平均亲水指数均为负值,都属于亲水性蛋白。OfERF72a、OfERF72b、OfERF73a不稳定系数分别为43.67、54.42、43.21,判断为不稳定的蛋白质;OfERF73b不稳定系数为38.40,判断为稳定的蛋白质。

图 4 B2亚组OfERFs氨基酸序列比对分析
Figure 4. Amino acid multiple sequence alignment analysis of OfERFs of subgroup B2
表 3 B2亚组OfERFs基本理化性质分析
Table 3. Analysis of basic physicochemical properties OfERFs of subgroup B2
基因名称 氨基酸数量/个 分子量/Da 理论等电点 不稳定系数 总平均亲水指数 OfERF72a 232 26 144 5.33 43.67 −0.744 OfERF72b 228 25 841 5.30 54.42 −0.796 OfERF73a 386 43 632 4.63 43.21 −0.739 OfERF73b 375 41 607 5.01 38.40 −0.710 -
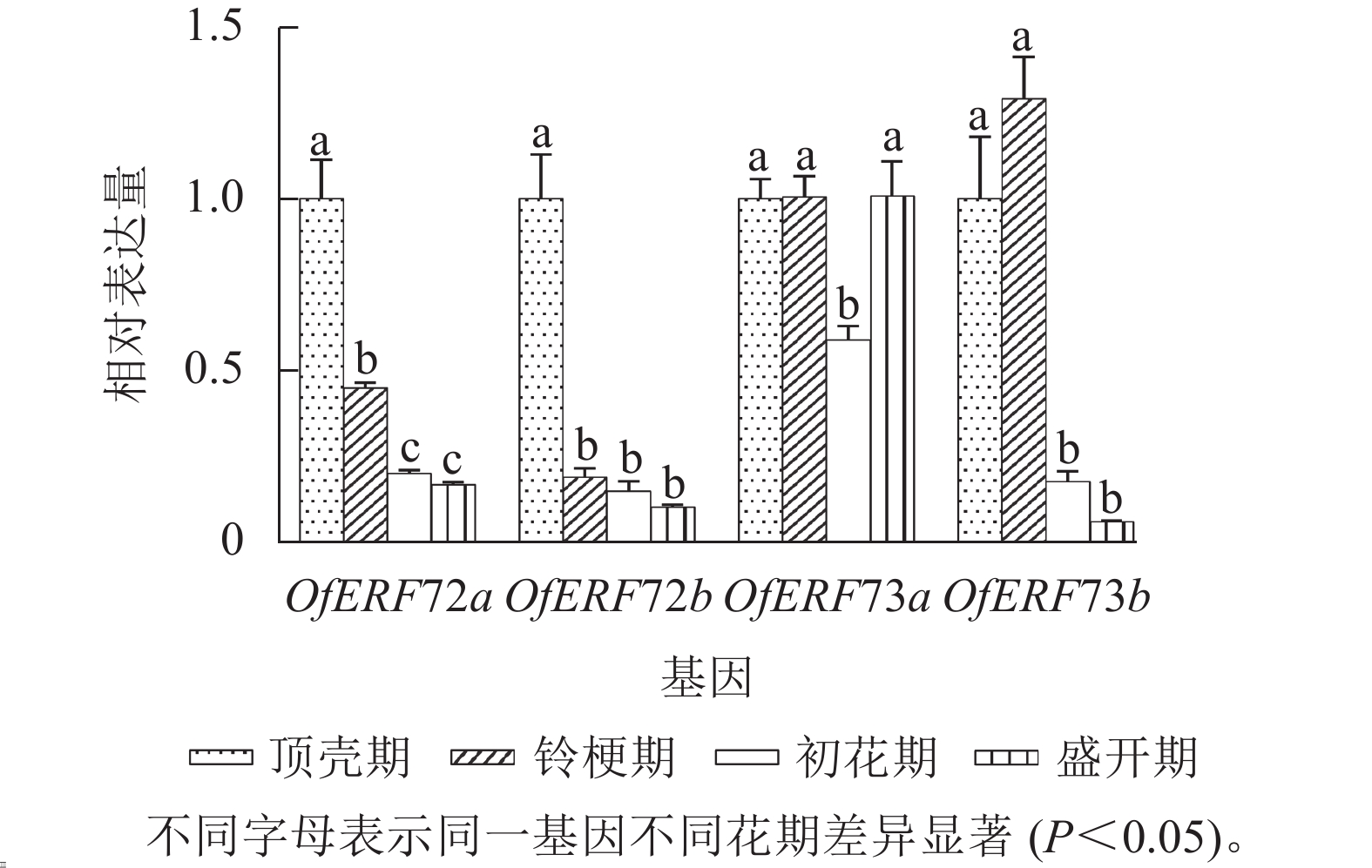
利用RT-qPCR技术分析‘堰虹桂’不同发育时期花瓣中OfERF72a、OfERF72b、OfERF73a与OfERF73b相对表达量(图5)发现:从顶壳期到盛开期,OfERF72a、OfERF72b的相对表达量基本呈现逐渐下降的趋势,OfERF73a的相对表达量在顶壳期、铃梗期与盛花期之间差异较小,在初花期相对表达量略有下降。OfERF73b的相对表达量在顶壳期、铃梗期较高,随后在初花期相对表达量显著下降(P<0.05)。
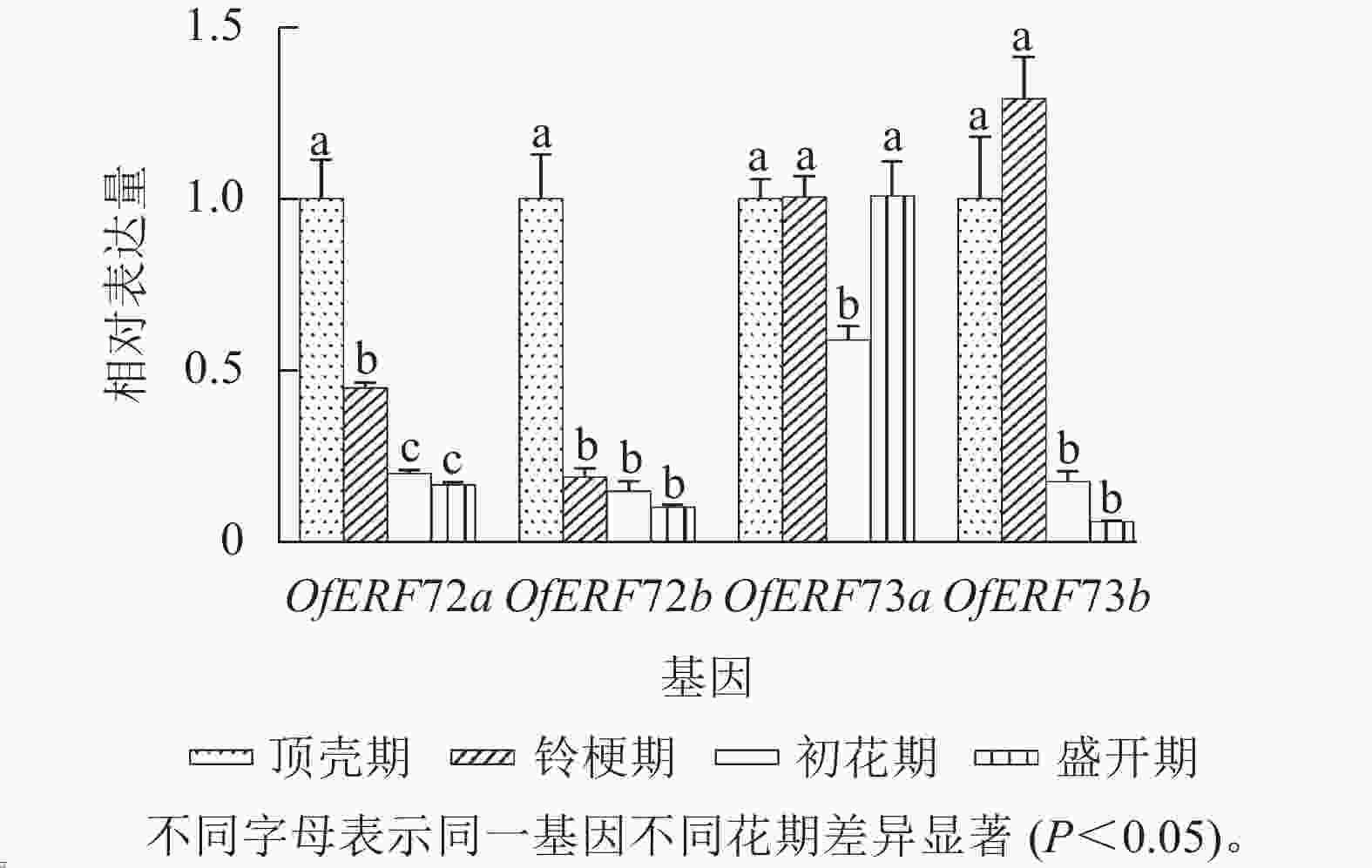
图 5 B2亚组OfERF在‘堰虹桂’不同花期的表达
Figure 5. Expression of OfERF genes of subgroup B2 at different flowering stages in O. fragrans ‘Yanhong Gui’
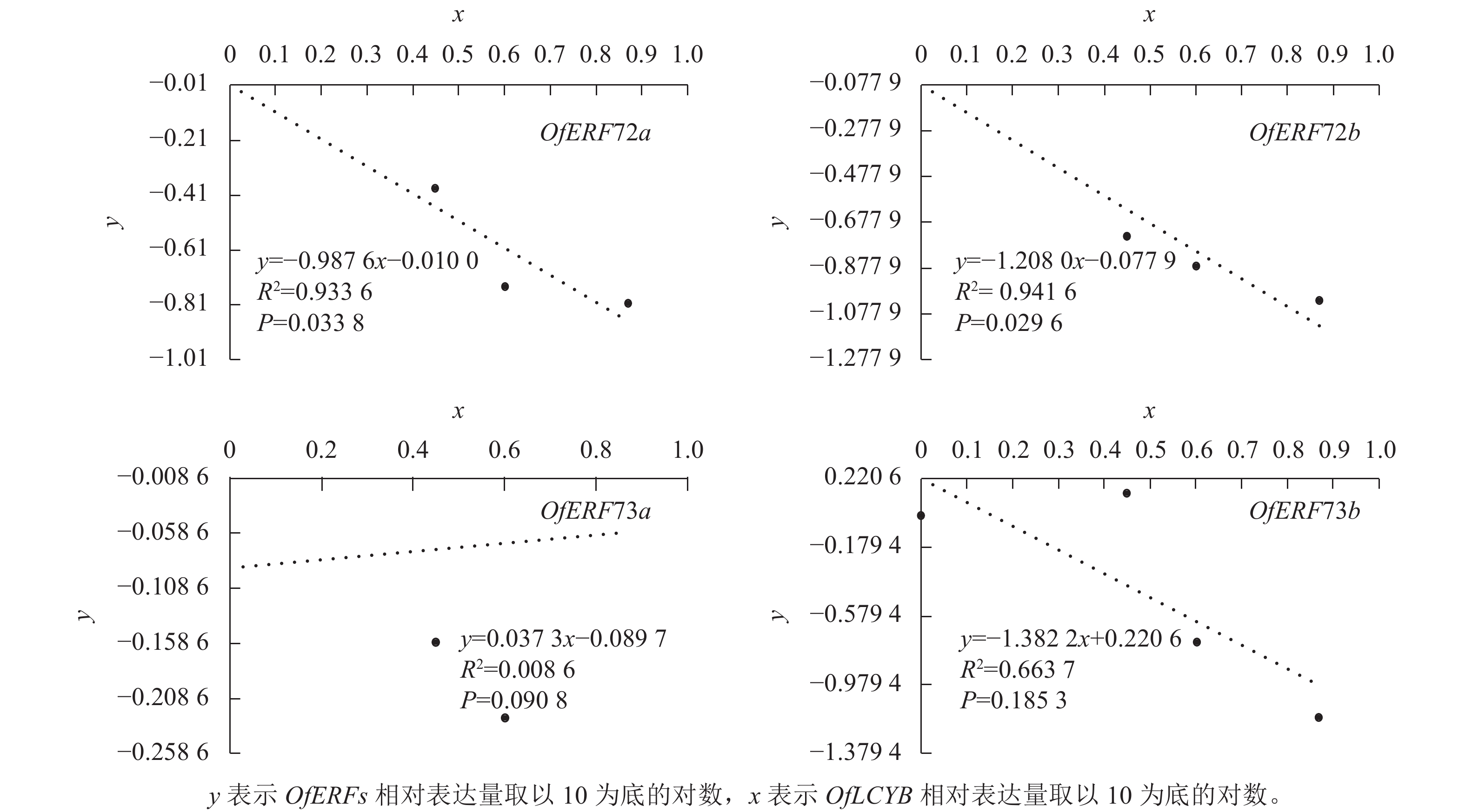
为了验证OfERFs与OfLCYB之间的关系,用y表示OfERFs的相对表达量取以10为底的对数,用x表示OfLCYB相对表达量取以10为底的对数进行相关性分析(图6)。其中,OfERF72a直线回归方程为y= − 0.987 6x − 0.010 0,决定系数(R2)为0.933 6,P=0.033 8;OfERF72b直线回归方程为y= − 1.208 0x − 0.077 9,R2=0.941 6,P=0.029 6。OfLCYB的表达水平与OfERF72a、OfERF72b呈显著负相关。
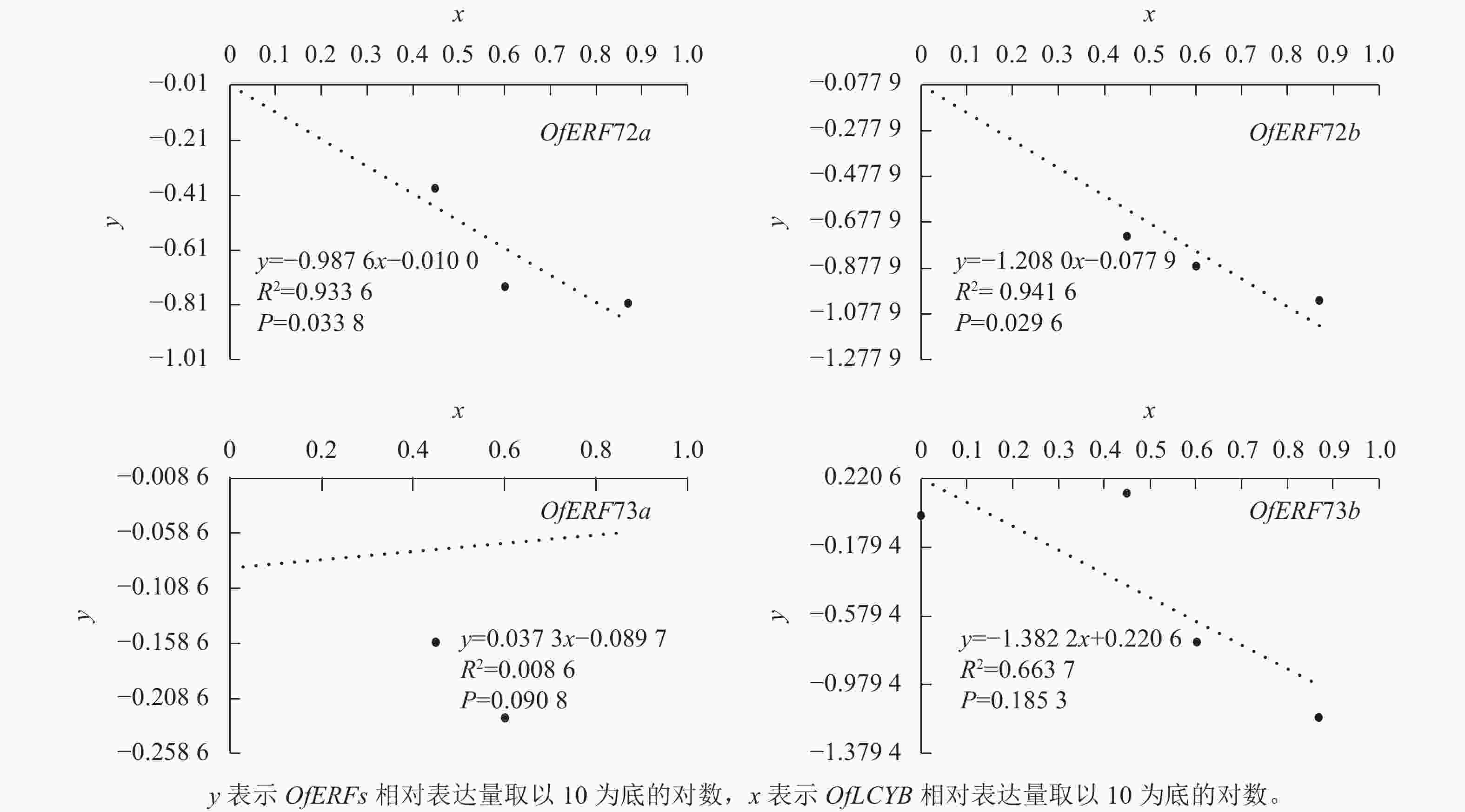
图 6 OfERFs与OfLCYB相对表达量的相关性分析
Figure 6. Correlation analysis of relative expression levels of OfERFs with OfLCYB
-
为了探究B2亚组OfERFs与OfLCYB启动子之间是否存在物理互作,同时将阴性对照pAbAi+pGADT7、阳性对照p53-AbAi+pGADT7-Rec-p53以及实验组pAbAi-OfLCYB-Pro+AD-OfERF分别接种于SD/-Leu与SD/-Leu/AbA (300 μg·L−1)的酵母培养基上,于30 ℃倒置培养3~5 d。结果发现(图7):在SD/-Leu培养基上,酵母均能正常生长,而在SD/-Leu/AbA (300 μg·L−1)培养基上,只有阳性对照与pAbAi-OfLCYB-Pro+AD-OfERF72b正常生长,其余酵母菌均不能生长,表明OfERF72b可以与OfLCYB启动子物理结合。

图 7 OfERF蛋白与OfLCYB启动子互作验证
Figure 7. Verification of physical interaction between OfERF proteins and OfLCYB promoter
-
本研究得到4个桂花‘堰虹桂’B2亚组的OfERFs基因,编码区长度为687~1 161 bp,编码228~386个氨基酸残基。拟南芥B2亚组ERF At1g53910.1、At1g72360.2、At2g47520.1、At3g14230.1以及At3g16770.1分别编码358、262、171、397和248个氨基酸残基[15]。牡丹Paeonia suffruticosa ERF家族中B2亚组基因PsERF1编码区长度为1 158 bp,编码385个氨基酸残基[19]。在番木瓜Carica papaya中,属于B2亚组的基因CpERF4、CpERF6、CpERF9则分别编码431、253、234个氨基酸残基[20]。而在番茄ERF中,其B2亚组的SlERF6、SlERF.E.1、SlERF90、SlERF91、SlERF.A.3分别编码255、260、386、1 454和372个氨基酸残基[21]。由此可以发现:同一物种B2亚组ERF基因编码不同长度的氨基酸序列,推测其不同成员的功能存在差异。
对4个桂花OfERFs基因的氨基酸序列进行系统进化分析,发现OfERFs与拟南芥B2亚组ERF聚集在一起,说明它们的同源性较高。其中2个基因与At3g16770.1 (AtERF72/AtRAP2.3)聚为一支,2个基因与At1g72360.2 (AtERF73/AtRAP2.2)聚为另一小支。据此将4个OfERFs基因分别命名OfERF72a与OfERF72b、OfERF73a与OfERF73b。桂花OfERF72与OfERF73均存在2个拷贝,说明桂花OfERF基因家族成员在进化和扩张过程中与基因重复事件有着紧密联系。在拟南芥中,AtERF72能够与缺铁反应基因IRT1、HA2和CLH1的启动子区域结合,负调控拟南芥的缺铁响应。与野生型植株相比,AtERF72突变体中铁和镁质量分数显著增加[22]。AtRAP2.3的同源基因SlERF6被证实是番茄中类胡萝卜素合成的负调控因子[13]。此外,在苹果中也有研究证明:AtRAP2.3的同源基因AP2D15可以负调控苹果PSY1和PSY2基因启动子序列中的ATCTA顺式作用元件[14]。拟南芥AtRAP2.2可以结合到拟南芥AtPSY启动子和AtPDS启动子的ATCTA元件上调控基因的表达,过表达AtRAP2.2后导致植物体内类胡萝卜素降低[15]。
桂花花瓣中主要类胡萝卜素为β-胡萝卜素,其生物合成由OfLCYB直接催化生成,是桂花花瓣中类胡萝卜素代谢的重要催化酶[23]。OfLCYB基因启动子中存在2个ATCTA顺式作用元件,推测其响应B2亚组ERF转录因子的调控。AtRAP2.2蛋白可以结合到拟南芥AtPSY启动子和AtPDS启动子的ATCTA元件上,从而调控相关基因的表达[15]。在苹果MdPSY1和MdPSY2基因启动子中也存在多个ATCTA顺式作用元件,能被AtRAP2.3的同源基因蛋白AP2D15强烈激活表达[14]。进一步研究发现:OfERF72a和OfERF72b的表达趋势与OfLCYB基因呈显著负相关。酵母单杂交结果表明:OfERF72b与OfLCYB启动子存在物理结合,表明B2亚组的OfERF72b可能通过结合OfLCYB基因启动子ATCTA顺式作用元件调控其表达。ATCTA元件也存在于桂花OfPSY[24]和OfCCD1[25]等其他类胡萝卜素代谢基因的启动子上,其是否响应B2亚组的ERF转录因子的调控需要进一步研究。
-
本研究基于桂花‘堰虹桂’转录组数据筛选了4个OfERF基因,OfERF72a与OfERF72b基因表达量均随着开花进程逐渐下降,与OfLCYB基因的表达量显著负相关。OfLCYB基因启动子含有2个ATCTA顺式作用元件,OfERF72b与OfLCYB启动子之间存在互作,表明OfERF72b可能参与调控OfLCYB的表达。
Screening and identification of ERF transcription factors of B2 subgroup involved in regulating lycopene β-cyclase gene LCYB in Osmanthus fragrans
-
摘要:
目的 筛选并鉴定参与调控桂花Osmanthus fragrans番茄红素β-环化酶OfLCYB基因的B2亚组ERF转录因子。 方法 以桂花品种‘堰虹桂’O. fragrans ‘Yanhong Gui’为材料,从桂花转录组数据库筛选B2亚组OfERF基因,通过生物信息学分析、实时荧光定量PCR (RT-qPCR)以及酵母单杂交技术,对OfERF基因序列和表达特性及其对OfLCYB基因启动子结合情况进行分析。 结果 OfLCYB基因启动子序列含有2个ATCTA顺式作用元件;基于桂花转录组数据库筛选出4个B2亚组ERF基因,均包含1个AP2保守结构域;RT-qPCR结果表明:OfERF72a与OfERF72b基因表达量均随着开花进程逐渐下降,与OfLCYB基因表达显著负相关,P分别为0.033 8、0.029 6;酵母单杂交结果证明:OfERF72b与OfLCYB启动子之间存在物理结合。 结论 OfERF72b可能通过调控OfLCYB的转录参与桂花类胡萝卜素的代谢。图7表3参25 Abstract:Objective This study aims to screen and identify ERF transcription factors of B2 subgroup involved in regulating lycopene β-cyclase OfLCYB gene of Osmanthus fragrans. Method ‘Yanhong Gui’, a cultivar of O. fragrans, was used as the material to screen OfERF genes of B2 subgroup from the O. fragrans transcriptome database. Bioinformatic analysis, real-time quantitative PCR (RT-qPCR) and yeast one-hybridization were used to analyze the sequence and expression characteristics of the OfERF gene and its binding to the OfLCYB gene promoter. Result The promoter of the OfLCYB gene contained two ATCTA cis-acting elements. Four OfERF genes of B2 subgroup were screened based on the O. fragrans transcriptome database, all of which contained an AP2 conserved structural domain. The RT-qPCR results showed that the expression levels of OfERF72a and OfERF72b genes gradually decreased with the flowering process, and were significantly negatively correlated with the expression of OfLCYB gene, with P values of 0.0338 and0.0296 , respectively. The results of yeast one-hybridization proved that there was a physical binding between OfERF72b and the OfLCYB promoter.Conclusion OfERF72b may participate in the metabolism of carotenoid in O. fragrans by regulating the transcription of OfLCYB. [Ch, 7 fig. 3 tab. 25 ref.] -
Key words:
- Osmanthus fragrans /
- carotenoid /
- ERF transcription factors /
- gene function
-
图 1 OfLCYB启动子的ATCTA顺式作用元件分析
Figure 1 Analysis of ATCTA cis-acting elements of the OfLCYB promoter
图 2 OfLCYB在‘堰虹桂’不同花期的表达
Figure 2 Expression of OfLCYB at different flowering stages in O. fragrans ‘Yanhong Gui’
图 3 桂花B2亚组OfERFs系统发育分析
Figure 3 Phylogenetic analysis of OfERFs in subgroup B2 of O. fragrans
图 4 B2亚组OfERFs氨基酸序列比对分析
Figure 4 Amino acid multiple sequence alignment analysis of OfERFs of subgroup B2
图 5 B2亚组OfERF在‘堰虹桂’不同花期的表达
Figure 5 Expression of OfERF genes of subgroup B2 at different flowering stages in O. fragrans ‘Yanhong Gui’
图 6 OfERFs与OfLCYB相对表达量的相关性分析
Figure 6 Correlation analysis of relative expression levels of OfERFs with OfLCYB
图 7 OfERF蛋白与OfLCYB启动子互作验证
Figure 7 Verification of physical interaction between OfERF proteins and OfLCYB promoter
表 1 PCR引物序列
Table 1. PCR primer sequences
引物名称 引物序列(5′→3′) LCYB-PRO-GW-F ggggacaagtttgtacaaaaaagcaggcttcCTGCTTCTTGTTGTTGTACG LCYB-PRO-GW-R ggggaccactttgtacaagaaagctgggtcCAATTTTGGCATGTTCTTAG OfLCYB-qF GAAAGGAGACGCCAAAGGGAG OfLCYB-qR GGAAGAAATAGCCGAGATGATAAGA 说明:小写字母表示部分attB序列。  下载: 导出CSV
下载: 导出CSV
表 2 OfERFs基因RT-qPCR引物序列
Table 2. RT-qPCR primer sequences of OfERFs
引物名称 引物序列(5′→3′) 引物名称 引物序列(5′→3′) OfERF73a-qF CTGAAGAGAAACCGCCAACAA OfERF72a-qR GGGTAGTAAACTTCTTGTTGCTGCGTA OfERF73a-qR TTAACGCCATCAGAAGACACAAGT OfERF72b-qF CAAATATCCTATGTTCAGAGG OfERF73b-qF AATTGGGATGCCGCCTCA OfERF72b-qR ATAGCATACCATAACATACCA OfERF73b-qR TTAAATCCCACCAAACATAGCACT OfACT-qF CCCAAGGCAAACAGAGAAAAAAT OfERF72a-qF CCAACCCCACCGGCTC OfACT-qR ACCCCATCACCAGAATCAAGAA
下载: 导出CSV
表 3 B2亚组OfERFs基本理化性质分析
Table 3. Analysis of basic physicochemical properties OfERFs of subgroup B2
基因名称 氨基酸数量/个 分子量/Da 理论等电点 不稳定系数 总平均亲水指数 OfERF72a 232 26 144 5.33 43.67 −0.744 OfERF72b 228 25 841 5.30 54.42 −0.796 OfERF73a 386 43 632 4.63 43.21 −0.739 OfERF73b 375 41 607 5.01 38.40 −0.710
下载: 导出CSV
-
[1] WANG Yiguang, ZHANG Chao, DONG Bin, et al. Carotenoid accumulation and its contribution to flower coloration of Osmanthus fragrans [J/OL]. Frontiers Plant Science, 2018, 9 : 1499[2024-03-25]. DOI: 10.3389/fpls.2018.01499. [2] MARHUENDA-MUÑOZ M, HURTADO-BARROSO S, TRESSERRA-RIMBAU A, et al. A review of factors that affect carotenoid concentrations in human plasma: differences between Mediterranean and Northern diets [J]. European Journal of Clinical Nutrition, 2019, 72(2): 18−25. [3] BALDERMANN S, KATO M, KUROSAWA M, et al. Functional characterization of a carotenoid cleavage dioxygenase 1 and its relation to the carotenoid accumulation and volatile emission during the floral development of Osmanthus fragrans Lour. [J]. Journal of Experimental Botany, 2010, 61(11): 2967−2977. [4] BALDERMANN S, KATO M, FLEISCHMANN P, et al. Biosynthesis of α- and β-ionone, prominent scent compounds, in flowers of Osmanthus fragrans [J]. Acta Biochimica Polonica, 2012, 59(1): 79−81. [5] HAN Yuanji, WANG Xiaohui, CHEN Weicai, et al. Differential expression of carotenoid-related genes determines diversified carotenoid coloration in flower petal of Osmanthus fragrans [J]. Tree Genetics & Genomes, 2014, 10: 329−338. [6] YUAN Hui, ZHANG Junxiang, NAGESWARAN D, et al. Carotenoid metabolism and regulation in horticultural crops [J/OL]. Horticulture Research, 2015, 2 (1): 15036[2024-03-25]. DOI: 10.1038/hortres.2015.36. [7] NISAR N, LI Li, LU Shan, et al. Carotenoid metabolism in plants [J]. Molecular Plant, 2015, 8(1): 68−82. [8] 沈子又, 张超, 董彬, 等. 桂花OfLCYB和OfLCYE启动子的克隆和活性分析[J]. 生物技术通报, 2018, 34 (1): 137−143. SHEN Ziyou, ZHANG Chao, DONG Bin, et al. Cloning and expression analysis of the promoters of OfLCYB and OfLCYE in Osmanthus fragrans [J]. Biotechnology Bulletin, 2018, 34 (1): 137−143. [9] NAKANO T, SUZUKI K, FUJIMURA T, et al. Genome-wide analysis of the ERF gene family in Arabidopsis and rice [J]. Plant Physiology, 2006, 140(2): 411−432. [10] SAGAWA J M, STANLEY L E, LAFOUNTAIN A M, et al. An R2R3-MYB transcription factor regulates carotenoid pigmentation in Mimulus lewisii flowers [J]. New Phytologist, 2015, 209(3): 1049−1057. [11] FU Changchun, HAN Yanchao, FAN Zhongqi, et al. The papaya transcription factor CpNAC1 modulates carotenoid biosynthesis through activating phytoene desaturase genes CpPDS2/4 during fruit ripening [J]. Journal of Agricultural & Food Chemistry, 2016, 64(27): 5454−5463. [12] HAO Dongyun, OHME-TAKAGI M, SARAI A. Unique mode of GCC box recognition by the DNA-binding domain of ethylene-responsive element-binding factor (ERF domain) in plant [J]. Journal of Biological Chemistry, 1998, 273(41): 26857−26861. [13] LEE J M, JOUNG J G, MCQUINN R, et al. Combined transcriptome, genetic diversity and metabolite profiling in tomato fruit reveals that the ethylene response factor SlERF6 plays an important role in ripening and carotenoid accumulation [J]. The Plant Journal, 2012, 70(2): 191−204. [14] AMPOMAH-DWAMENA C, DRIEDONKS N, LEWIS D, et al. The phytoene synthase gene family of apple (Malus×domestica) and its role in controlling fruit carotenoid content [J/OL]. BMC Plant Biology, 2015, 15 : 185[2024-03-25]. DOI: 10.1186/s12870-015-0573-7. [15] WELSCH R, MAASS D, VOEGEL T, et al. Transcription factor RAP2.2 and its interacting partner SINAT2: stable elements in the carotenogenesis of Arabidopsis leaves [J]. Plant Physiology, 2007, 145(3): 1073−1085. [16] 杨青杰. 黄龙胆类胡萝卜素生物合成基因启动子的功能分析[D]. 长春: 东北师范大学, 2013. YANG Qingjie. Functional Characterization of the Gentiana lutea Carotenoid Biosynthesis Gene Promoters [D]. Changchun: Northeast Normal University, 2013. [17] ZHANG Chao, WANG Yiguang, FU Jianxin, et al. Transcriptomic analysis and carotenogenic gene expression related to petal coloration in Osmanthus fragrans ‘Yanhong Gui’ [J]. Trees, 2016, 30: 1207−1223. [18] ZHANG Chao, FU Jianxin, WANG Yiguang, et al. Identification of suitable reference genes for gene expression normalization in the quantitative real-time PCR analysis of sweet osmanthus (Osmanthus fragrans Lour. ) [J/OL]. PLoS One, 2015, 10 (8): e0136355[2024-03-25]. DOI: 10.1371/journal.pone.0136355. [19] 吴凡, 张超, 郭加, 等. 牡丹切花ERF转录因子基因的分离与表达分析[J]. 园艺学报, 2016, 43 (1): 109−120. WU Fan, ZHANG Chao, GUO Jia, et al. Isolation and expression analysis of ERF transcription factor genes in tree peony cut flowers [J]. Acta Horticulturae Sinica, 2016, 43 (1): 109−120. [20] 陈永萍, 高峰, 申艳红, 等. 番木瓜ERF家族与果实成熟相关成员的分析[J]. 园艺学报, 2019, 46 (2): 252−264. CHEN Yongping, GAO Feng, SHEN Yanhong, et al. The analysis of ERFs related to fruit ripening in papaya [J]. Acta Horticulturae Sinica, 2019, 46 (2): 252−264. [21] 徐志璇. 番茄AP2/ERF超家族重鉴定及SlERF. D. 3基因的功能研究[D]. 泰安: 山东农业大学, 2020. XU Zhixuan. Reidentification of Tomato AP2/ERF Transcription Factor Superfamily and Functional Analysis of SlERF. D. 3 [D]. Taian: Shandong Agricultural University, 2020. [22] LIU Wei, LI Qiwei, WANG Yi, et al. Ethylene response factor AtERF72 negatively regulates Arabidopsis thaliana response to iron deficiency [J]. Biochemical and Biophysical Research Communications, 2017, 491(3): 862−868. [23] QING Hongshen, CHEN Jiahong, JIANG Lingli, et al. Functional characterization of two lycopene cyclases from sweet osmanthus (Osmanthus fragrans) [J/OL]. Scientia Horticulturae, 2022, 299 : 11062[2024-03-25]. DOI: 10.1016/j.scienta.2022.111062. [24] 周俊杰, 王艺光, 董彬, 等. 桂花OfPSY、OfPDS和OfHYB基因启动子克隆及表达特性分析 [J]. 浙江农林大学学报, 2023, 40(1): 64−71. ZHOU Junjie, WANG Yiguang, DONG Bin, et al. Cloning and expression characterization of OfPSY, OfPDS and OfHYB gene promoters in Osmanthus fragrans [J]. Journal of Zhejiang A&F University, 2023, 40(1): 64−71. [25] 刘玉成, 王艺光, 张超, 等. 桂花OfCCD1基因启动子克隆与表达特性 [J]. 浙江农林大学学报, 2018, 35(4): 596−603. LIU Yucheng, WANG Yiguang, ZHANG Chao, et al. Cloning and transient expression assay of OfCCD1 gene promoters from Osmanthus fragrans [J]. Journal of Zhejiang A&F University, 2018, 35(4): 596−603. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20240316
 点击查看大图
点击查看大图
计量
- 文章访问数: 1578
- HTML全文浏览量: 404
- PDF下载量: 32
- 被引次数: 0
