







-
miRNA是一类小的单链非编码核糖核酸序列,长度为22~24个核苷酸,通过与靶基因3′UTR区域结合进而抑制靶基因的表达[1-3]。miRNA具有高度的保守性、时序性和组织特异性,能够调控生物体特定的生理功能,在生物体生长、发育和疾病发生等过程中发挥着重要的作用[4]。网格蛋白(clathrin)介导的胞吞是信号分子进入细胞的主要途径,也是病毒进入细胞的重要途径。病毒侵入细胞是病毒增殖最为关键的一步,而网格蛋白介导的胞吞是病毒侵入细胞最主要也是最为典型的一种细胞胞吞途径。网格蛋白又称笼形蛋白[5],由PEARSE在1975年首次分离得到并命名[6]。网格蛋白的1个重链(clathrin heavy chain, CLTC)和1个轻链组成一个二聚体,3个二聚体组成三联体骨架结构,多个三联体骨架结构组成五边形或六边形网格结构的包被亚基,最后由这些包被亚基构成多面体的网格蛋白包被的囊泡结构-小窝,病毒即是通过与小窝蛋白上的病毒受体结合进入细胞的[7]。如登革病毒(dengue virus, DNV)[8-9],猴出血热病毒(simian hemorrhagic fever virus, SHFV)[10],丙型肝炎病毒(hepatitis C virus, HCV)[11-13],乙型脑炎病毒(japanese encephalitis virus, JEV)[14],蓝耳病毒(porcine reproductive and respiratory syndrome, PRRSV)[15],肠道病毒71型(enterovirus 71, EV-71)[16]等。该途径的抑制能够有效阻止病毒的感染,如用siRNA敲低CLTC基因,能够有效阻断乙脑病毒对猪Sus scrofa肾上皮细胞PK15的感染[14]。本研究旨在筛选出一批能够靶向猪CLTC基因miRNA,从而为筛选广谱抗病毒因子提供基础。鉴于双荧光素酶报告分析法的常规性和荧光定量的灵敏性,本研究结合2种方法,以增加基因筛选的可靠性。利用双荧光素酶报告系统获得靶向猪CLTC基因的miRNA:miR-205,miR-1,miR-129-5p和miR-206,并以点突变实验鉴定出miR-1通过种子区结合靶向作用于猪CLTC基因3′UTR。
-
幼小仓鼠Mesocricetus auratus肾细胞系(BHK-21)和猪肾上皮细胞系(PK15)购自中国典型培养物保藏中心;双荧光素酶检测试剂盒和psiCHECK2载体均购自Promega;miRNA模拟物(miRNA mimics)购自上海吉玛制药技术有限公司;Lipofectamine 2000购自Invitrogen;T4 DNA连接酶,DNA片段,限制性内切酶XholⅠ和NotⅠ购自Fermentas公司;SYBR Green试剂购自TOYOBO公司;HiFiScript cDNA第一链合成试剂盒购自北京康为世纪生物科技有限公司;引物由上海博尚生物技术有限公司合成。
-
根据miRBase(http://www.mirbase.org)数据库公布的猪miRNA(Release 21, 2014),利用Targetscan(http://www.targetscan.org),BioGps(http://www.biogps.org)软件进行miRNA的靶向分析;使用miRNAMap()软件鉴定miRNA的表达特异性,筛选出靶向猪CLTC基因3′UTR序列的miRNA,并合成相应的猪miRNA模拟物(上海吉玛制药技术有限公司)。
-
利用美国生物技术信息中心(NCBI)数据库中猪CLTC基因序列,设计引物并扩增猪CLTC基因3′UTR序列;采用XholⅠ和NotⅠ酶切位点,与psiCHECK2载体连接获得双荧光素酶报告载体,命名为Wild Type(WT)。设计miRNA种子区和猪CLTC基因3′UTR结合区的突变引物(表 1),使用引物重叠聚合酶链式反应(PCR)的方法,以猪CLTC基因3′UTR片段为模板,扩增得到CLTC基因3′UTR的上下游片段;再以上下游同源臂的混合物作为模板,PCR扩增得到含有突变结合位点的CLTC基因3′UTR目的片段;与psiCHECK2载体连接,经测序验证后获得CLTC靶位点突变报告载体,命名为Mutant Type(MT)。
表 1 构建含猪CLTC-3′UTR突变序列的双荧光素酶载体的突变引物
Table 1. Primers for constructing luciferase reporter gene vectors containing porcine CLTC mutation 3′UTR area
引物名称 引物序列(5'-3') 对应的miRNA MT1-R CGAAAAGTAGTCCAAGTAGAAATAAAGGTTACAAGAACA miR-205 MT1-F TTTCTACTTGGACTACTTTTCGTTTCTAACTGTAAAACTTGGA MT2-F TCTGTAACCGTATCATTTTAGAATTTATTTTCAAAGGG miR-1/206 MT2-R TAAAATGATACGGTTACAGAAATAAGCTTTTAACATAGGT MT3-F TAAACGTTTAATATTGGTATGTGACCATGCAAGACTGT miR-129-5p MT3-R CATACCAATATTAAACGTTTAGCTTTTCTTTGAATAAAAG Xhol I-F CCCTCGAGGACGGGAAGCTGATCCTGTAGT Not I-R ATTTGCGGCCGCTTCCACAAACAAAACTGAAGAACAG -
用含10%胎牛血清(体积分数) 的MEM(minimum essential media)培养基于体积分数为5%二氧化碳,37 ℃的环境条件下培养BHK-21细胞,并进行细胞传代。转染前一天将细胞接种至24孔板,接种密度为2.5×105个·孔-1,培养过夜后按照Lipofectamine 2000的说明书进行细胞瞬时转染。设置8个实验组:① 无miRNA无转染试剂的空白对照;② 阴性对照(NC),WT;③ miR-205,WT;④ miR-1,WT;⑤ miR-129-5p,WT;⑥ miR-206,WT;⑦ miR-19a,WT;⑧ miR-19b,WT。转染24 h后,收集细胞,按照双荧光检测试剂盒说明书进行荧光检测,计算萤火虫荧光素酶和海肾荧光素酶活性的比值,确定不同样品之间报告基因的激活程度。同时测定每组3个平行孔之间的相对发光比率(RLU),计算标准误差,统计不同转染组之间的差异。
-
以猪cDNA为模板设计扩增CLTC基因的定量引物,12孔板培养PK15细胞,接种密度为5×105个·孔-1,培养过夜后对细胞进行转染。设置8个实验组:① NC,WT;② miR-205,WT;③ miR-1,WT;④ miR-129-5p,WT;⑤ miR-206,WT;⑥ miR-19a,WT;⑦ miR-19b,WT;⑧ 无miRNA无转染试剂的空白对照。转染48 h后收集细胞,Trizol试剂提取细胞总RNA,并反转录成cDNA,定量PCR检测CLTC基因的mRNA的表达变化,统计每组CLTC基因的表达差异。
-
用双荧光素酶检测试剂盒检测miRNA对点突变重组载体荧光素酶活性值的影响,实验组设置如下:① 无miRNA无转染试剂的空白对照;② NC,WT;③ miR-205,WT;④ miR-205,MT;⑤ miR-1,WT;⑥ miR-1,MT;⑦ miR-206,WT;⑧ miR-206,MT;⑨ miR-129-5p,WT;⑩ miR-129-5p,MT。按照1.2.3进行双荧光素酶报告基因检测实验。
-
所有数据均以“平均数±标准差”表示。统计学分析方法采用SAS 8.0的t检验和GLM方差分析,P < 0.05表示差异显著,P < 0.01表示差异极显著。为消除误差,设置重复3次·实验组-1,取平均值。
-
利用生物信息学分析软件,初步预测出与猪CLTC基因3’UTR有互补结合位点的6条miRNA:miR-205,miR-1,miR-129-5p,miR-206,miR-19a,miR-19b(表 2),进行后续实验验证。
表 2 与猪CLTC基因3′UTR可能结合的miRNA
Table 2. Potential miRNA targeting 3′UTR of porcine CLTC gene
miRNA模拟物 CLTC-3'UTR碱基位置 表达特异性 miRNA成熟序列 (5'—3‘) miR-205 111-119 胸腺 UCCUUCAUUCCACCGGAGUCUG miR-1 173-181 肌肉 UGGAAUGUAAAGAAGUAUGUA miR-206 173-181 肌肉 UGGAAUGUAAGGAAGUGUGUGA miR-129-5p 437-443 肌肉 CUUUUUGCGGUCUGGGCUUGC miR-19a 303-310 脾脏 UGUGCAAAUCUAUGCAAAACUGA miR-19b 303-310 卵巢 UGUGCAAAUCCAUGCAAAACUGA -
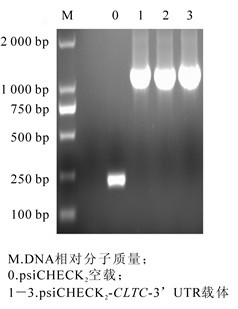
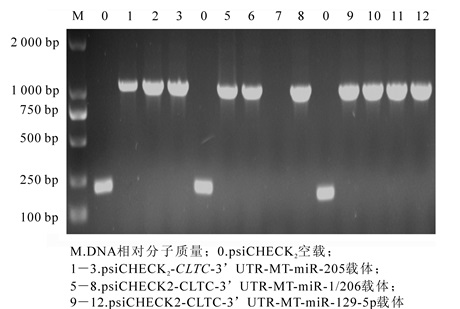
将猪CLTC基因3′UTR片断和CLTC-3′UTR点突变片断分别克隆至双荧光酶报告基因载体psiCHECK2中,获得重组质粒psiCHECK2-CLTC-3′UTR(图 1),并以鉴定引物进行菌液PCR鉴定。检测长度为1 200 bp的猪CLTC基因3′UTR的双荧光素酶载体(图 2)和猪CLTC基因3′UTR的双荧光素酶靶标突变载体psiCHECK2-3′UTR-MT(图 3),测序验证后提取质粒备用。

图 1 CLTC-3′UTR双荧光素酶报告基因载体的构建图谱
Figure 1. Constructing of luciferase reporter gene vectors or porcine CLTC-3′UTR vectors

图 2 菌液PCR验证猪CLTC-3'UTR双荧光素酶报告基因载体
Figure 2. Microbial validation of luciferase report gene vector

图 3 菌液PCR鉴定猪CLTC-3'UTR双荧光素酶靶位点突变载体
Figure 3. Microbial validation of porcine CLTC-3'UTR mutation vectors
-
双荧光酶报告基因载体(WT)分别与6种miRNA模拟物共转染BHK-21细胞,以NC为阴性对照,细胞培养24 h以后,双荧光素酶检测试剂盒检测荧光素酶活性(图 4)。结果显示:miR-205,miR-1,miR-206和miR-129-5p能显著降低荧光素酶活性值。miRNA模拟物转染PK15细胞以后,荧光定量PCR检测CLTC基因的表达量(图 5),结果显示:miR-1和miR-129-5p能够显著降低CLTC基因mRNA的表达水平,而miR-205和miR-206与对照组相比,差异不显著。

图 4 miRNA模拟物与psiCHECK2-CLTC-3′UTR共转染BHK-21细胞的相对荧光素酶活性
Figure 4. Relative luciferase activity of psiCHECK2-CLTC-3′UTR reporter cotransfected with miRNA mimics in BHK-21 cells

图 5 定量PCR检测miRNA模拟物转染PK15细胞后CLTC基因的表达量
Figure 5. CLTC gene expression was detected by q-PCR method after transfecting with miRNA mimics in PK15 cells
-
为了进一步验证各miRNA与猪CLTC-3′UTR结合的靶位点,我们选取筛选出的miR-205,miR-1,miR-206,miR-129-5p做下一步的点突变实验。将miRNA对猪CLTC基因3′UTR靶位点进行突变,其中miR-206和miR-1的靶位点相同,所以突变载体相同,分别构建miR-205,miR-129-5p,miR-1/ssc-miR-206对应CLTC靶点的突变载体。miRNA与突变载体(MT)或双荧光报告基因载体(WT)共转染BHK-21细胞后检测荧光素酶活性。结果显示:突变质粒psiCHECK2-CLTC-3′UTR和miR-1共转染细胞后,与对照组相比,其荧光素酶活性得以恢复(图 6),说明miR-1通过种子区域作用于猪CLTC基因的3′UTR抑制其表达。

图 6 种子区突变前后4种miRNA模拟物对荧光素酶表达的影响
Figure 6. Relative luciferase activity of different reporters in the presence or absence of miRNA seed sequence in BHK-21 cell
-
miRNA广泛存在于生物体内,对生物体的转录后基因表达调控起着关键的作用[17]。在植物细胞中,成熟的miRNA先与一种称为RNA诱导沉默复合体(RNA-induced silencing complex,RISC)的复合物结合,再特异性地与目标mRNA结合,引起mRNA的降解;动物细胞中,大部分的miRNA与其靶mRNA不完全互补,miRNA则通过与对应的mRNA的3′端非翻译区(3′UTR)结合阻止转录后的翻译,起到调节基因表达的作用[18]。愈来愈多的研究证实,miRNA一般通过2种方式调控病毒在宿主细胞内的增殖,一种是miRNA直接靶向病毒的mRNA序列,抑制病毒相关基因的表达,这类miRNA已有相关的研究报道,如miR-323,miR-491,miR-654靶向甲型H1N1流感病毒(H1N1 subtype influenza A virus)[20],miR-24和miR-93靶向水泡口炎病毒(vesicular stomatitis virus, VSV)[21]。另一种是miRNA通过靶向宿主基因调节细胞内的信号通路进而阻止病毒的增殖,这类miRNA的研究集中在脂质代谢方面,如miR-27a通过靶向控制脂质合成和运输的基因维甲酸X受体α(RXRα)和ATP结合盒转运子A1(ABCA1)调控脂质代谢,从而抑制丙型肝炎病毒的复制[22]。截至目前,在miRBase数据库软件中已鉴定出的人类miRNA有1 881种,小鼠的有1 193种[19],猪的有382种,但对猪miRNA的生物学功能尚不清楚。本研究利用生物信息学软件预测出能够与猪CLTC基因3′UTR靶向结合的6条miRNA:miR-205,miR-1,miR-206,miR-129-5p,miR-19a,miR-19b,利用双荧光素酶报告基因法和荧光定量PCR法进一步筛选出能够靶向配对的miRNA。结果显示:miR-129-5p,miR-1在双荧光报告基因实验和荧光定量PCR中都能够显著降低CLTC基因的表达量,而miR-205,miR-206在双荧光报告基因实验中能够显著降低CLTC基因的表达量,但在荧光定量PCR中差异不显著。为进一步验证靶向猪CLTC基因的miRNA,我们设计了点突变载体,将miRNA和其对应点突变载体共转染细胞检测荧光素酶表达活性。结果显示:miR-1通过种子区的结合而抑制靶基因猪CLTC基因的表达,miR-129-5p能够抑制CLTC基因的表达,但突变载体的荧光活性并没有得到恢复,说明miR-129-5p可能不是因为靶基因种子区的结合而抑制CLTC基因的表达,推测可能存在其他的靶作用位点。截至目前,有关miR-129-5p对病毒侵入细胞的研究鲜有报道,前期研究发现虾miR-1通过抑制CLTC基因的表达调控细胞的吞噬作用[23],推测本研究鉴定出的miR-129-5p可能在病毒侵入细过程中发挥重要作用。
本研究成功构建了包含猪CLTC基因3′UTR的双荧光酶报告基因载体和不同miRNA种子区域所对应的点突变报告载体,成功筛选出靶向猪CLTC基因的miR-1和miR-129-5p,利用点突变实验最终确定miR-1通过种子区靶向结合猪CLTC基因并抑制其表达,研究结果为探究miRNA-CLTC基因-clathrin胞吞通路在抵抗病毒侵入宿主细胞的研究打下基础,也为抗病候选基因的筛选提供了新的素材。
Prediction and validation of miRNA targeting the porcine CLTC gene
-
摘要: 网格蛋白介导的胞吞是病毒侵入细胞的重要途径,网格蛋白重链(clathrin heavy chain,CLTC)是形成网格蛋白小窝结构的重要组成部分。针对CLTC基因的转录后调控特别是调控猪Sus scrofa CLTC的miRNA目前还不太清楚。本研究旨在筛选出调控猪CLTC基因的miRNA。利用生物信息学方法预测出6个靶向猪CLTC基因的miRNA,将猪CLTC基因3'UTR克隆至双荧光素酶报告基因载体psiCHECK2中获得双荧光素酶报告基因重组载体psiCHECK2-CLTC-3'UTR。将预测得到的miRNA分别和重组载体psiCHECK2-CLTC-3'UTR共转染到细胞中,以乱序序列作为阴性对照(NC),检测miRNA对重组质粒荧光素酶活性的影响。结果发现miR-205,miR-1,miR-129-5p和miR-206均能够显著抑制荧光素酶活性(P < 0.05)。在猪肾上皮细胞系PK15细胞中超表达miR-1和miR-129-5p后,定量PCR(q-PCR)结果显示:猪CLTC基因的表达量显著下调。突变了psiCHECK2-CLTC-3'UTR载体中这4个miRNA的种子序列的结合位点发现:miR-1对突变质粒中的荧光素酶无显著抑制作用。表明miR-1与猪CLTC基因有直接的靶向关系,并通过其种子序列抑制CLTC基因的表达。
-
关键词:
- 猪 /
- CLTC /
- miRNA /
- 胞吞 /
- 双荧光素酶报告基因载体
Abstract: Clathrin heavy chain (CLTC), an important component of clathrin-coated pits, plays an important role with virus invasion of cells; however, post-transcriptional gene regulation of the CLTC gene, especially porcine CLTC gene regulation by miRNA has not yet been clearly elucidated. This study aimed to screen miRNAs that target the CLTC gene. First, bioinformatics predicted that miR-205, miR-1, miR-129-5p, miR-206, miR-19a, and miR-19b targeted the porcine CLTC gene. Then porcine CLTC 3'UTR was cloned into the psiCHECK2 vector, and the dual luciferase reporter recombinant vector psiCHECK2-3'UTR was constructed. The prediction of miRNA and the recombinant vector psiCHECK2-3'UTR were co-transfected into cells, respectively, with the scramble sequence of miRNA as a negative control (NC); then the luciferase activity was detected. A quantitative PCR (q-PCR) was also used to determine the expression of CLTC mRNA levels. Then to verify whether miRNA regulated the porcine CLTC gene through seed sequences, binding sites of psiCHECK2-3'UTR with seed sequence were mutated. Results showed that miR-205, miR-1, miR-129-5p, and miR-206 were able to significantly inhibit luciferase activity (P < 0.05). At the same time there was an overexpression of miR-1 and miR-129-5p in PK15 cells, and the q-PCR showed that the expression of CLTC mRNA level was significantly reduced (P < 0.05). Verification of whether the four miRNA (miR-205, miR-1, miR-129-5p, and miR-206) regulated the porcine CLTC gene through seed sequences showed that miR-1 mutant plasmid did not inhibit the luciferase activity. Thus, the results demonstrated that miR-1 inhibited porcine CLTC gene expression through its seed sequence binding with CLTC 3'UTR.-
Key words:
- porcine /
- CLTC /
- miRNA /
- endocylosis /
- dual luciferase reporter gene vector
-
图 1 CLTC-3′UTR双荧光素酶报告基因载体的构建图谱
Figure 1 Constructing of luciferase reporter gene vectors or porcine CLTC-3′UTR vectors
图 2 菌液PCR验证猪CLTC-3'UTR双荧光素酶报告基因载体
Figure 2 Microbial validation of luciferase report gene vector
图 3 菌液PCR鉴定猪CLTC-3'UTR双荧光素酶靶位点突变载体
Figure 3 Microbial validation of porcine CLTC-3'UTR mutation vectors
图 4 miRNA模拟物与psiCHECK2-CLTC-3′UTR共转染BHK-21细胞的相对荧光素酶活性
**表示P < 0.01,差异极显著;*表示P< 0.05,差异显著。
Figure 4 Relative luciferase activity of psiCHECK2-CLTC-3′UTR reporter cotransfected with miRNA mimics in BHK-21 cells
图 5 定量PCR检测miRNA模拟物转染PK15细胞后CLTC基因的表达量
*表示P < 0.05,差异显著。
Figure 5 CLTC gene expression was detected by q-PCR method after transfecting with miRNA mimics in PK15 cells
图 6 种子区突变前后4种miRNA模拟物对荧光素酶表达的影响
*表示P < 0.05,差异显著。
Figure 6 Relative luciferase activity of different reporters in the presence or absence of miRNA seed sequence in BHK-21 cell
表 1 构建含猪CLTC-3′UTR突变序列的双荧光素酶载体的突变引物
Table 1. Primers for constructing luciferase reporter gene vectors containing porcine CLTC mutation 3′UTR area
引物名称 引物序列(5'-3') 对应的miRNA MT1-R CGAAAAGTAGTCCAAGTAGAAATAAAGGTTACAAGAACA miR-205 MT1-F TTTCTACTTGGACTACTTTTCGTTTCTAACTGTAAAACTTGGA MT2-F TCTGTAACCGTATCATTTTAGAATTTATTTTCAAAGGG miR-1/206 MT2-R TAAAATGATACGGTTACAGAAATAAGCTTTTAACATAGGT MT3-F TAAACGTTTAATATTGGTATGTGACCATGCAAGACTGT miR-129-5p MT3-R CATACCAATATTAAACGTTTAGCTTTTCTTTGAATAAAAG Xhol I-F CCCTCGAGGACGGGAAGCTGATCCTGTAGT Not I-R ATTTGCGGCCGCTTCCACAAACAAAACTGAAGAACAG  下载: 导出CSV
下载: 导出CSV
表 2 与猪CLTC基因3′UTR可能结合的miRNA
Table 2. Potential miRNA targeting 3′UTR of porcine CLTC gene
miRNA模拟物 CLTC-3'UTR碱基位置 表达特异性 miRNA成熟序列 (5'—3‘) miR-205 111-119 胸腺 UCCUUCAUUCCACCGGAGUCUG miR-1 173-181 肌肉 UGGAAUGUAAAGAAGUAUGUA miR-206 173-181 肌肉 UGGAAUGUAAGGAAGUGUGUGA miR-129-5p 437-443 肌肉 CUUUUUGCGGUCUGGGCUUGC miR-19a 303-310 脾脏 UGUGCAAAUCUAUGCAAAACUGA miR-19b 303-310 卵巢 UGUGCAAAUCCAUGCAAAACUGA
下载: 导出CSV
-
[1] WANG Yang, STRICKER H M, GUO Deming, et al. MicroRNA:past and present[J]. Front BioSci, 2007, 12(6):2316-2329. [2] MOORE M J, SCHEEL T K, LUNA J M, et al. miRNA-target chimeras reveal miRNA 3'-end pairing as a major determinant of Argonaute target specificity[J]. Nat Commun, 2015, 6:8864. doi:10. 1038/ncomms 9864. [3] ROY-CHAUDHURI B, VALDMANIS P N, ZHANG Y, et al. Regulation of microRNA-mediated gene silencing by microRNA precursors[J]. Nat Struct Mol Biol, 2014, 21(9):825-832. [4] KETTING R F. MicroRNA biogenesis and function:an overview[J]. Adv Exp Med Biol, 2011, 700:1-14. [5] BRODSKY F M. Diversity of clathrin function:new tricks for an old protein[J]. Annu Rev Cell Dev Biol, 2012, 28(1):309-336. [6] PEARSE B M. Clathrin:a unique protein associated with intracellular transfer of membrane by coated vesicles[J]. Proc Nat Acad Sci, 1976, 73(4):1255-1259. [7] McMAHON H T, BOUCROT E. Molecular mechanism and physiological functions of clathrin-mediated endocytosis[J]. Nat Rev Mol Cell Biol, 2011, 12(8):517-533. [8] ACOSTA E G, CASTILLA V, DAMONTE E B. Functional entry of dengue virus into Aedes albopictus mosquito cells is dependent on clathrin-mediated endocytosis[J]. J Gen Virol, 2008, 89(Pt 2):474-484. [9] PICCINI L E, CASTILLA V, DAMONTE E B. Dengue-3 virus entry into vero cells:role of clathrin-mediated endocytosis in the outcome of infection[J]. PLoS One, 2015, 10(10):e0140824. doi:10. 1371/journal.pone. 0140824. [10] CAI Yingyun, POSTNIKOVA E N, BERNBAUM J G, et al. Simian hemorrhagic fever virus cell entry is dependent on CD163 and uses a clathrin-mediated endocytosis-like pathway[J]. J Virol, 2015, 89(1):844-856. [11] BLAISING J, LÉVY P L, GONDEAU C, et al. Silibinin inhibits hepatitis C virus entry into hepatocytes by hindering clathrin-dependent trafficking[J]. Cell Microbiol, 2013, 15(11):1866-1882. [12] BENEDICTO I, GONDAR V, MOLINA-JIMÉNEZ F, et al. Clathrin mediates infectious hepatitis C virus particle egress[J]. J Virol, 2015, 89(8):4180-4190. [13] BLANCHARD E, BELOUZARD S, GOUESLAIN L, et al. Hepatitis C virus entry depends on clathrin-mediated endocytosis[J]. J Virol, 2006, 80(14):6964-6972. [14] YANG Songbai, HE Minhui, LIU Xiangdong, et al. Japanese encephalitis virus infects porcine kidney epithelial PK15 cells via clathrin-and cholesterol-dependent endocytosis[J]. Virol J, 2013, 10(1):258. doi:10. 1186/1743-422X-10-258. [15] HUANG Li, ZHANG Yuanpeng, YU Yaling, et al. Role of lipid rafts in porcine reproductive and respiratory syndrome virus infection in MARC-145 cells[J]. Biochem Biophys Res Commun, 2011, 414(3):545-550. [16] HUSSAIN K M, LEONG K L, NG M M, et al. The essential role of clathrin-mediated endocytosis in the infectious entry of human enterovirus 71[J]. J Biol Chem, 2011, 286(1):309-321. [17] HUNTZINGER E, IZAURRALDE E. Gene silencing by microRNAs:contributions of translational repression and mRNA decay[J]. Nat Rev Genet, 2011, 12(2):99-110. [18] 姜雪鸥, 钟金城. miRNA的研究进展及其展望[J].中国草食动物, 2011, 31(4):63-67. JIANG Xueou, ZHONG Jincheng. Progress in miRNA research and its prospect[J]. China Herbivore Sci, 2011, 31(4):63-67. [19] 戴丽荷, 褚晓红, 路伏增, 等.靶向猪ATGL基因的miRNA预测及鉴定[J].畜牧兽医学报, 2015, 46(8):1281-1289. DA Lihe, CHU Xiaohong, LU Fuzeng, et al. Prediction and validation of miRNA targeting porcine ATGL gene[J]. Acta Vet Zootech Sin, 2015, 46(8):1281-1289. [20] SONG Liping, LIU He, GAO Shijuan, et al. Cellular microRNAs inhibit replication of the H1N1 influenza A virus in infected cells[J]. J Virol, 2010, 84(17):8849-8860. [21] OTSUKA M, JING Qing, GEORGEL P, et al. Hypersusceptibility to vesicular stomatitis virus infection in Dicer1-deficient mice is due to impaired miR24 and miR93 expression[J]. Immunity, 2007, 27(1):123-134. [22] SHIRASAKI T, HONDA M, SHIMAKAMI T, et al. MicroRNA-27a regulates lipid metabolism and inhibits hepatitis C virus replication in human hepatoma cells[J]. J Virol, 2013, 87(9):5270-5286. [23] LIU Cuilian, WANG Jiajia, ZHANG Xiaobo. The involvement of miR-1-clathrin pathway in the regulation of phagocytosis[J]. PLoS One, 2014, 9(6):e98747. doi:10. 1371/joumal.pone. 0098747. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2017.03.002
 点击查看大图
点击查看大图

计量
- 文章访问数: 4048
- HTML全文浏览量: 1039
- PDF下载量: 567
- 被引次数: 0
