




-
凋落物是森林生态系统中植被代谢的产物。凋落物分解对森林土壤有机质形成和植被群落组成起着决定性作用[1-2],是维持森林生态系统物质循环和能量流动的关键因素[3-5]。凋落物分解包括从新鲜凋落物到粗腐殖质,粗腐殖质进一步分解为腐殖质2个阶段。新鲜凋落物分解为腐殖质的过程中会逐步释放营养元素,使处于不同分解阶段的凋落物养分含量及化学计量比不同。凋落叶作为凋落物的主要组成成分,分解速度快,在养分循环中占据重要的地位[6]。据统计,森林土壤中至少有90%的氮和磷、60%的其他矿质元素来自凋落叶的分解归还[7],因此,凋落叶分解对土壤性质产生直接影响[8],对土壤有机质提升和养分积累具有重要作用。同时,土壤中碳氮磷养分元素及化学计量比也在不同程度上影响凋落叶的分解[9-10],尤其是氮磷元素在诸多生态系统中会对凋落叶分解产生复杂的交互作用。此外,环境因子也与凋落叶分解密切相关,其中温度和湿度被认为是影响凋落叶分解的主要因子。温度和湿度可以直接或通过改变凋落叶质量、土壤酶活性、土壤微生物活性及植被群落组成间接影响凋落叶分解[11]。因此,研究凋落叶不同分解阶段化学计量特征及影响因素,可深入了解其在分解过程中养分归还状况。
黔西南地区地势西高东低,大部分地区为山地与丘陵,石漠化问题严重,是典型的喀斯特生境和生态脆弱区[12]。该区森林类型丰富,不仅分布有马尾松Pinus massoniana林、毛竹Phyllostachys edulis林、杉木Cunninghamia lanceolata林等人工林,还有以滇青冈Cyclobalanopsis glaucoides、麻栎Quercus acutissima、安顺润楠Machilus cavaleriei等为优势种的天然林。为了更好地了解喀斯特地区森林养分循环规律,为喀斯特地区生态系统恢复重建和科学管理提供理论指导,研究者探究了植被与土壤间碳氮磷化学计量特征[13-16]及不同林分类型凋落物的养分释放规律和归还特征[17-19]。由于对凋落物养分释放特征的研究时长较短,忽略了凋落物在长期分解过程中的营养元素释放规律和迁移特征对森林生态系统养分循环产生的影响。本研究利用空间代替时间的方法,以马尾松林、毛竹林、杉木林以及天然林为对象,根据凋落叶分解状态,将凋落叶分为未分解层、半分解层和已分解层[20-22],采集并测定了处于3种分解阶段凋落叶的全碳、全氮和全磷质量分数,同时,测定了不同林分土壤环境因子,并对凋落叶化学计量特征与环境因子间的相关关系进行了分析,以期为喀斯特地区森林生态系统养分循环以及生态系统功能研究奠定基础。
-
研究区位于贵州省西南部的兴义市、望谟县、平坝县和普定县境内(24°42′45″~26°31′36″N,104°38′39″~106°14′08″E),属亚热带湿润季风气候,平均海拔为1 443.5 m,年平均气温为14.8 ℃,年平均降水量为1 100~1 400 mm。该区分布有马尾松、毛竹、杉木、麻栎、大叶青冈Cyclobalanopsis jenseniana、云山青冈C. sessilifolia、滇青冈、安顺润楠等林分类型。林下植被以悬钩子属Rubus、狗脊Woodwardia japonica、里白Diplopterygium glaucum、芒萁Dicranopteris pedata、紫萁Osmunda japonica等为主。选取该区4种典型林分类型(马尾松林、毛竹林、杉木林和以麻栎、安顺润楠和滇青冈为优势种组成的天然林),每种林分分别设置3个20 m×20 m的样地。样地基本信息如表1所示。
表 1 样地信息表
Table 1. Information table of sample plots
林分类型 纬度(N) 经度(E) 海拔/m 坡位 坡向 坡度/(°) 郁闭度/% 干扰程度 经营措施 马尾松林 24°42′45″ 104°38′39″ 1 050 中 南偏西7° 25.0 50 中 去除林下植被、无施肥 26°29′59″ 106°12′15″ 1 480 上 南偏东75° 20.0 30 重 去除林下植被、无施肥 26°29′54″ 106°12′13″ 1 444 下 东南 9.0 58 重 去除林下植被、无施肥 毛竹林 26°31′36″ 106°14′08″ 1 462 下 东 7.5 80 重 去除林下植被、无施肥 26°31′36″ 106°14′08″ 1 500 中 南偏西60° 12.0 50 重 去除林下植被、无施肥 26°31′36 106°14′05″ 1 476 中 东南 8.0 85 重 去除林下植被、无施肥 杉木林 24°59′48″ 104°48′44″ 1 890 中 北偏西15° 16.0 40 重 去除林下植被、无施肥 25°22′09″ 106°08′02″ 1 530 中 北偏东60° 45.0 70 轻 去除林下植被、无施肥 25°14′01″ 106°08′40″ 1 250 上 南偏东60° 15.0 70 中 去除林下植被、无施肥 天然林 25°22′00″ 106°08′02″ 1 490 下 北偏西60° 40.0 70 轻 无经营措施 25°06′15″ 105°05′25″ 1 330 上 北 30.0 80 轻 无经营措施 26°14′44″ 105°45′36″ 1420 中上 北偏东58° 39.0 60 中 无经营措施 -
2020年11月5—20日采集样地中的凋落叶。具体做法是:在每个样地内随机选取3个面积为1 m2的小样方,将每个小样方中的凋落叶分为未分解(可看清凋落叶形状,多为新鲜叶片)、半分解(凋落叶进入分解发酵阶段,形状不完整,包含杂质,与未分解凋落叶有明显的区别)和已分解(看不出凋落叶形状,为腐殖质状态)3种类型,分别进行采集,采集量为2.28~4.36 t·hm−2。采集完成后将同一个样地中的3个小样方对应层次的凋落叶混合均匀,共取得36个样品。同时,采集样方中0~10 cm土壤,并混合均匀,并用土壤温度测试仪测定并记录土壤温度。
将凋落叶和土壤装入自封袋带回实验室,凋落叶在65 ℃下烘干至恒量,全自动研磨仪磨碎,过100目筛;土壤自然风干过100目筛。测定凋落叶和土壤全碳(TC)、全氮(TN)和全磷(TP)质量分数。全碳和全氮采用元素分析仪(PE 2400Ⅱ ,美国)进行测定,全磷采用碱熔-钼锑抗比色法测定[23]。土壤pH采用玻璃电极法测定(土水比为1.0∶2.5);土壤含水率采用鲜土烘干法测定。
-
利用Excel 2010对数据进行处理,采用SPSS 24.0对凋落叶化学计量特征进行双因素(林分类型和凋落叶分解阶段)方差分析,并在林分类型和分解阶段下对化学计量特征进行单因素方差分析(one-way ANOVA),采用LSD法(P<0.05)对各指标在不同处理间的差异进行显著性分析,采用Pearson检验分析凋落叶化学计量特征与环境因子的相关性,用Origin 2018作图。
-
双因素(林分类型和凋落叶分解阶段)方差分析表明:林分类型对全碳、全氮和全磷质量分数均产生显著影响(P<0.05),凋落叶分解阶段对全碳和全磷质量分数有显著影响(P<0.05)。随着凋落叶分解程度的加剧,全碳质量分数呈现降低趋势;全氮质量分数呈现升高趋势;全磷质量分数除天然林呈升高趋势外,其余呈现降低趋势。两者交互效应对全碳、全氮和全磷质量分数均无显著影响。
由图1可见:各林分类型凋落叶各分解阶段全碳质量分数总和从大到小为天然林(1 466.37 mg·g−1)、马尾松林(1 465.07 mg·g−1)、杉木林(1 455.93 mg·g−1)、毛竹林(1 246.53 mg·g−1)。随着凋落叶分解程度的加剧,4种林分凋落叶全碳质量分数均呈现降低趋势,其中,马尾松林和杉木林凋落叶已分解阶段全碳质量分数均显著低于未分解和半分解阶段(P<0.05),其余2种林分在不同分解阶段无显著差异。不同林分类型凋落叶全碳质量分数具有明显差异,毛竹林凋落叶全碳质量分数在各分解阶段下均显著低于其余3种林分(P<0.05)。

图 1 不同林分类型和分解阶段的凋落叶全碳、全氮和全磷质量分数
Figure 1. Carbon, nitrogen and phosphorus contents of leaf litters in different decomposition stages in four forest types
各林分类型凋落叶各分解阶段全氮质量分数总和从大到小依次为毛竹林(47.53 mg·g−1)、天然林(42.4 mg·g−1)、杉木林(32.57 mg·g−1)、马尾松林(31.9 mg·g−1)。随着凋落叶分解程度的加剧,马尾松林和天然林凋落叶全氮质量分数先减少后增加;毛竹林逐渐增加;杉木林先增加后减少,且未分解阶段显著低于半分解和已分解阶段(P<0.05)。在不同林分类型凋落叶中,全氮质量分数在各分解阶段均无显著差异。
各林分类型凋落叶各分解阶段全磷质量分数总和从大到小依次为天然林(8.20 mg·g−1)、杉木林(4.49 mg·g−1)、马尾松林(4.39 mg·g−1)、毛竹林(4.17 mg·g−1)。随着凋落叶分解程度的加剧,杉木林凋落叶全磷质量分数呈现逐渐降低趋势;马尾松林、毛竹林和天然林凋落叶全磷质量分数呈现先降低后升高的趋势,其中,天然林半分解阶段显著低于未分解和已分解阶段(P<0.05)。对不同林分类型进行分析,在半分解阶段中,马尾松林和毛竹林凋落叶全磷质量分数均显著低于天然林(P<0.05);已分解阶段,马尾松林、杉木林和毛竹林凋落叶全磷质量分数均显著低于天然林(P<0.05)。
-
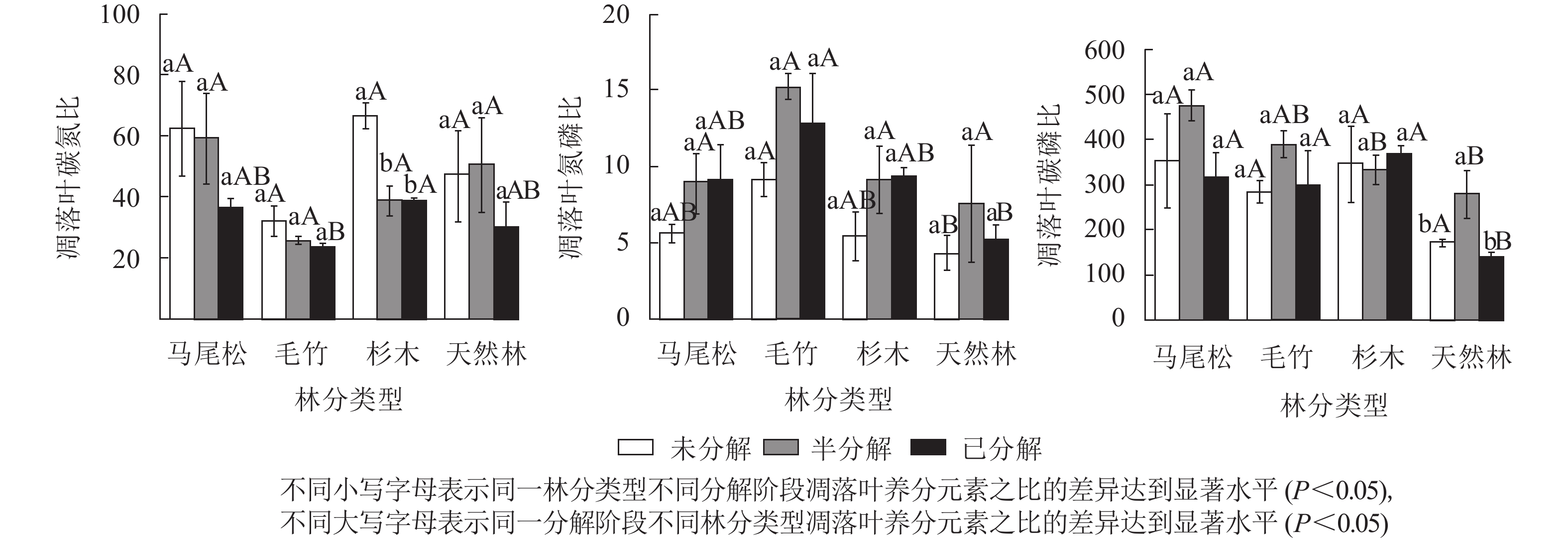
双因素(林分类型和凋落叶分解阶段)方差分析表明:林分类型和凋落叶分解阶段对其化学计量比均产生显著影响(P<0.05):随着凋落叶分解程度的加剧,凋落叶碳氮比和氮磷比分别呈现出降低和升高的趋势;对于凋落叶碳磷比而言,马尾松林和天然林呈降低趋势,毛竹林和杉木林呈升高趋势。两者交互效应对凋落叶碳氮比、氮磷比和碳磷比均无显著影响。
4种林分类型不同分解阶段凋落叶碳氮比、氮磷比和碳磷比如图2所示。马尾松林、毛竹林、杉木林和天然林凋落叶碳氮比平均值分别为52.72、27.09、48.11、42.72,凋落叶碳氮比随凋落叶逐渐分解大体呈现出降低趋势,杉木林半分解和已分解阶段碳氮比显著低于未分解阶段(P<0.05)。对不同林分类型进行分析,杉木林已分解阶段碳氮比显著高于毛竹林(P<0.05)。

图 2 不同林分类型和不同分解阶段下的凋落叶碳、氮、磷化学计量特征
Figure 2. Carbon, nitrogen and phosphorus stoichiometry characteristicsof leaf litters in different decomposition stages in four forest types
马尾松林、毛竹林、杉木林和天然林凋落叶氮磷比平均值分别为7.87、12.39、7.99、7.97。随着凋落叶分解程度的加剧,马尾松林和杉木林凋落叶氮磷比呈现逐渐升高的趋势;毛竹林和天然林凋落叶氮磷比呈现先升高后降低的趋势,且各分解阶段之间无显著差异。对不同林分类型进行分析,在未分解和已分解阶段中,天然林氮磷比显著低于毛竹林(P<0.05),半分解阶段中无显著差异。
马尾松林、毛竹林、杉木林和天然林凋落叶碳磷比平均值分别为381.35、324.66、349.26、227.82。随着凋落叶分解程度的加剧,马尾松林、毛竹林和天然林凋落叶碳磷比呈现先升高后降低的趋势;杉木林凋落叶碳磷比呈先降低后升高的趋势;天然林半分解阶段凋落叶碳磷比显著高于未分解和已分解阶段(P<0.05)。对不同林分类型进行分析,在半分解阶段中,杉木和天然林凋落叶碳磷比均显著低于马尾松林(P<0.05),已分解阶段中,天然林凋落叶碳磷比显著低于其余3种林分(P<0.05)。
-
凋落叶全碳、全氮、全磷质量分数及化学计量特征与环境因子相关分析如表2所示。凋落叶全碳质量分数与土壤温度和坡度极显著正相关(P<0.01),与林分郁闭度显著负相关(P<0.05);凋落叶全氮质量分数与坡位和土壤含水率显著负相关(P<0.05);凋落叶全磷质量分数与土壤碳氮比显著负相关(P<0.05),与土壤氮磷比、土壤碳磷比、土壤温度、土壤pH、坡度显著正相关(P<0.05, P<0.01)。
表 2 凋落叶化学计量特征与环境因子的相关分析
Table 2. Pearson correlations between stoichiometric characteristics of leaf litters and environmental factors
项目 凋落叶全碳 凋落叶全氮 凋落叶全磷 凋落叶碳氮比 凋落叶氮磷比 凋落叶碳磷比 土壤全碳 0.125 −0.047 0.130 0.136 −0.041 −0.093 土壤全氮 0.110 −0.022 0.265 0.124 −0.122 −0.243 土壤全磷 0.052 −0.074 −0.229 0 0.104 0.217 土壤碳氮比 0.073 −0.029 −0.336* 0.022 0.262 0.386* 土壤氮磷比 0.231 −0.220 0.377* 0.296 −0.341* −0.343* 土壤碳磷比 0.230 −0.222 0.332* 0.296 −0.305 −0.295 土壤温度 0.439** −0.193 0.565** 0.267 −0.556** −0.418* 土壤含水率 −0.007 −0.340* −0.318 0.251 0.013 0.253 土壤pH 0.167 −0.224 0.461** 0.216 −0.462** −0.453** 海拔 0.015 0.013 −0.275 −0.075 0.190 0.239 郁闭度 −0.419* 0.197 −0.192 −0.212 0.371* 0.086 坡度 0.450** −0.096 0.469** 0.187 −0.456** −0.358* 坡位 0.282 −0.386* 0.256 0.328 −0.411* −0.217 说明:*和**分别表示在P<0.05和P<0.01下显著相关 凋落叶全碳、全氮、全磷化学计量与环境因子的关系是:凋落叶碳氮比除与海拔和林分郁闭度呈负相关关系外,与其余环境因子均呈正相关关系,但相关性不显著;对凋落叶氮磷比而言,除受土壤氮磷比影响外(P<0.05),还与土壤温度、土壤pH、坡位、坡度呈显著负相关关系(P<0.05, P<0.01),与林分郁闭度呈显著正相关(P<0.05);凋落叶碳磷比与土壤碳氮比和土壤氮磷比分别呈显著正相关和负相关(P<0.05),还与土壤温度、土壤pH、坡度呈显著负相关(P<0.05, P<0.01)。
-
凋落叶在分解过程中养分释放大致分为3种模式:①直接释放,养分含量出现延续性减少状态;②淋溶—富集—释放,养分含量先减少后增多再减少,这一过程中养分含量变化具有较大的波动性;③富集—释放,为了满足微生物参与凋落叶分解过程的需要,养分含量随时间变化先升高后降低[24]。本研究中,不同养分元素的释放规律有所差异。4种林分凋落叶全碳质量分数呈直接释放规律,从大到小均表现为未分解层、半分解层、已分解层。这与赵畅等[25]研究结果类似,凋落叶分解过程中可溶性糖和有机质的损失,以及分解微生物的消耗都可能造成碳的减少。对氮元素而言,马尾松林和天然林呈现淋溶—富集规律,表明凋落叶分解过程中为满足微生物生命活动的需要而吸收了一定量的氮。SULKAVA等[26]研究指出:马尾松林凋落物在分解过程中氮呈现出的富集规律,杉木林和毛竹林也呈现富集规律。这是由于氮的释放滞后于凋落叶的质量损失。杉木林凋落叶中全磷为直接释放,表明杉木林更易受到磷元素的限制。凋落叶碳氮比、碳磷比和氮磷比反映其自身分解速率及植物生长过程中受何种元素的限制[27]。随着凋落叶逐渐分解,杉木林碳氮比显著降低,与王书丽等[27]研究的研究结果一致,表明其分解初期受微生物影响分解迅速;天然林凋落叶碳磷比未分解阶段显著高于其分解阶段,这可能是由于土壤微生物对磷的固持效应相关[28];4种林分凋落叶分解后期氮磷比均有增大趋势,证明林分易受到氮或磷元素的限制。
-
林分类型可以直接或通过改变土壤微生物群落结构间接影响凋落叶养分及化学计量特征[29-30],这与本研究得出的林分类型对其影响显著的结果一致。含氮量较高的凋落叶会降低分解惰性碳的微生物活性[31],其碳的留存量会升高。本研究得出毛竹林高氮低碳的结论,证明了林分类型影响凋落物养分释放,通常与林分自身性质相关。本研究发现毛竹林全磷质量分数低于马尾松林,这可能是由于该地区土壤微环境不同而导致。马尾松林、毛竹林、杉木林、天然林凋落叶全氮平均质量分数分别为10.63、15.84、10.86、14.13 mg·g−1,全磷平均质量分数分别为1.46、1.39、1.50、2.73 mg·g−1,均高于全球森林凋落叶氮(10.0 mg·g−1)和磷(0.7 mg·g−1)质量分数[32]。这可能与氮、磷元素重吸收程度有关。本研究得出所有林分凋落叶碳氮比、碳磷比和氮磷比均小于全球森林的平均水平[33],碳磷比也低于亚热带常绿阔叶林和亚热带人工林凋落叶平均值[34],这与该地区富含大量磷元素有关。氮磷比较小除与该区富含磷元素有关,还可能受叶片结构的影响。
-
凋落叶分解主要受非生物因素和生物因素共同影响[35]。非生物因素包括环境因子、凋落叶自身质量等,生物因素包括土壤微生物丰富度和多样性。土壤化学计量比、土壤pH、土壤温度和土壤湿度通过改变土壤微生物群落结构间接影响凋落叶的养分释放[36]。在本研究中,土壤化学计量比、土壤温湿度、土壤pH、林分郁闭度、坡度对凋落叶养分质量分数均有显著影响。较高的土壤湿度改变了土壤微生物的生长活性,影响凋落叶中氮的释放,进而改变了凋落叶碳氮比。本研究得出土壤含水率、土壤温度、土壤pH、坡度与凋落叶碳氮比相关不显著,尤其是土壤含水率对凋落叶化学计量比之间的相关均不显著,这是因为该区气温较高、林分郁闭度大的特点影响了某些土壤微生物群落对凋落叶的分解活动。坡度通过改变土壤表面径流影响凋落叶养分分布规律;郁闭度改变了由水热变化引起的一系列生物化学过程从而导致凋落叶的养分迁移;海拔可以通过改变气温和降水间接影响凋落叶分解。海拔对凋落叶养分及其化学计量比的影响不显著的可能原因是本研究没有在大范围尺度上研究海拔与凋落叶养分之间的关系。
-
林分类型对凋落叶养分质量分数及化学计量特征均有显著影响,凋落叶分解阶段对全碳和全磷质量分数及化学计量特征均有显著影响。随着凋落叶分解程度的加剧,4种林分凋落叶全碳质量分数均呈现出释放规律,马尾松林和杉木林已分解阶段全碳质量分数均显著低于未分解和半分解阶段;杉木林全氮质量分数呈现富集—释放的规律;天然林全磷质量分数呈现淋溶—富集的规律。杉木林未分解阶段凋落叶碳氮比显著高于半分解和已分解阶段;天然林碳磷比先升高后降低,且其半分解阶段碳磷比显著高于未分解和已分解阶段。
Stoichiometric characteristics of leaf litter at different decomposition stages in 4 forest types
-
摘要:
目的 研究黔西南地区不同林分凋落叶不同分解阶段的化学计量特征,深入了解喀斯特地区不同森林生态系统养分循环规律。 方法 选取黔西南地区4种典型林分类型,包括马尾松Pinus massoniana林、毛竹Phyllostachys edulis林、杉木Cunninghamia lanceolata林,以及以麻栎Quercus acutissima、安顺润楠Machilus cavaleriei和滇青冈Cyclobalanopsis glaucoides为优势种的天然林,采集处于不同分解阶段的森林凋落叶,并测定其全碳、全氮和全磷化学计量特征。 结果 ①毛竹林各分解阶段全碳质量分数均显著低于其余林分(P<0.05),马尾松林和杉木林已分解阶段凋落叶全碳质量分数均显著低于未分解和半分解阶段(P<0.05);杉木林未分解阶段全氮质量分数显著低于半分解和已分解阶段(P<0.05);天然林已分解阶段全磷质量分数显著高于其余林分(P<0.05),且其半分解阶段全磷质量分数显著低于未分解和已分解阶段(P<0.05)。②杉木林已分解阶段碳氮比显著高于毛竹林(P<0.05),且其未分解阶段碳氮比显著高于半分解和已分解阶段(P<0.05);毛竹林未分解和已分解阶段氮磷比显著高于天然林(P<0.05);杉木林和天然林半分解阶段碳磷比均显著低于马尾松林(P<0.05),天然林已分解阶段碳磷比显著低于其余林分(P<0.05)。 结论 林分类型和分解阶段对凋落叶全碳、全氮和全磷质量分数及化学计量特征均有显著影响。图2表2参36 Abstract:Objective This research aims to study the stoichiometric characteristics of leaf litter at different decomposition stages in different stands in southwest Guizhou Province, so as to better understand the nutrient cycling law of different forest ecosystems in karst areas. Method 4 typical forest types were selected in southwest Guizhou, including Pinus massoniana forest, Phyllostachys edulis forest, Cunninghamia lanceolata forest, and natural forest dominated by Quercus acutissima, Machilus cavaleriei and Cyclobalanopsis glaucoides. Leaf litter at different decomposition stages was collected and stoichiometric characteristics of C, N and P were measured. Result (1) The total C content of leaf litter in each decomposition stage of P. edulis forest was significantly lower than that in other stands (P<0.05), and the total C content of leaf litter at decomposed stage of P. massoniana and C. lanceolata forests was significantly lower than that at undecomposed and semi-decomposed stages (P<0.05). The total N content of leaf litter in C. lanceolata forest at undecomposed stage was significantly lower than that at semi-decomposed and decomposed stages (P<0.05). The total P content of leaf litter at decomposed stage of natural forest was significantly higher than that in other stands (P< 0.05), and the total P content at semi-decomposed stage of natural forest was significantly lower than that at undecomposed and decomposed stages (P< 0.05). (2) The C/N ratio of C. lanceolata forest at decomposed stage was significantly higher than that of Ph. edulis forest (P<0.05), and the C/N ratio at undecomposed stage was significantly higher than that at semi-decomposed and decomposed stages (P<0.05). The N/P ratio at undecomposed and decomposed stages of Ph. edulis forest was significantly higher than that of natural forest (P<0.05). The C/P ratio of leaf litter at semi-decomposed stage of C. lanceolata and natural forests was significantly lower than that of P. massoniana forest (P< 0.05), and the C/P ratio at decomposed stage of natural forest was significantly lower than that of other stands (P<0.05). Conclusion Both forest type and decomposition stage have significant effects on contents of total C, N and P, as well as stoichiometric characteristics of leaf litter. [Ch, 2 fig. 2 tab. 36 ref.] -
Key words:
- forest type /
- leaf litter /
- decomposition stage /
- stoichiometry
-
图 1 不同林分类型和分解阶段的凋落叶全碳、全氮和全磷质量分数
Figure 1 Carbon, nitrogen and phosphorus contents of leaf litters in different decomposition stages in four forest types
图 2 不同林分类型和不同分解阶段下的凋落叶碳、氮、磷化学计量特征
Figure 2 Carbon, nitrogen and phosphorus stoichiometry characteristicsof leaf litters in different decomposition stages in four forest types
表 1 样地信息表
Table 1. Information table of sample plots
林分类型 纬度(N) 经度(E) 海拔/m 坡位 坡向 坡度/(°) 郁闭度/% 干扰程度 经营措施 马尾松林 24°42′45″ 104°38′39″ 1 050 中 南偏西7° 25.0 50 中 去除林下植被、无施肥 26°29′59″ 106°12′15″ 1 480 上 南偏东75° 20.0 30 重 去除林下植被、无施肥 26°29′54″ 106°12′13″ 1 444 下 东南 9.0 58 重 去除林下植被、无施肥 毛竹林 26°31′36″ 106°14′08″ 1 462 下 东 7.5 80 重 去除林下植被、无施肥 26°31′36″ 106°14′08″ 1 500 中 南偏西60° 12.0 50 重 去除林下植被、无施肥 26°31′36 106°14′05″ 1 476 中 东南 8.0 85 重 去除林下植被、无施肥 杉木林 24°59′48″ 104°48′44″ 1 890 中 北偏西15° 16.0 40 重 去除林下植被、无施肥 25°22′09″ 106°08′02″ 1 530 中 北偏东60° 45.0 70 轻 去除林下植被、无施肥 25°14′01″ 106°08′40″ 1 250 上 南偏东60° 15.0 70 中 去除林下植被、无施肥 天然林 25°22′00″ 106°08′02″ 1 490 下 北偏西60° 40.0 70 轻 无经营措施 25°06′15″ 105°05′25″ 1 330 上 北 30.0 80 轻 无经营措施 26°14′44″ 105°45′36″ 1420 中上 北偏东58° 39.0 60 中 无经营措施  下载: 导出CSV
下载: 导出CSV
表 2 凋落叶化学计量特征与环境因子的相关分析
Table 2. Pearson correlations between stoichiometric characteristics of leaf litters and environmental factors
项目 凋落叶全碳 凋落叶全氮 凋落叶全磷 凋落叶碳氮比 凋落叶氮磷比 凋落叶碳磷比 土壤全碳 0.125 −0.047 0.130 0.136 −0.041 −0.093 土壤全氮 0.110 −0.022 0.265 0.124 −0.122 −0.243 土壤全磷 0.052 −0.074 −0.229 0 0.104 0.217 土壤碳氮比 0.073 −0.029 −0.336* 0.022 0.262 0.386* 土壤氮磷比 0.231 −0.220 0.377* 0.296 −0.341* −0.343* 土壤碳磷比 0.230 −0.222 0.332* 0.296 −0.305 −0.295 土壤温度 0.439** −0.193 0.565** 0.267 −0.556** −0.418* 土壤含水率 −0.007 −0.340* −0.318 0.251 0.013 0.253 土壤pH 0.167 −0.224 0.461** 0.216 −0.462** −0.453** 海拔 0.015 0.013 −0.275 −0.075 0.190 0.239 郁闭度 −0.419* 0.197 −0.192 −0.212 0.371* 0.086 坡度 0.450** −0.096 0.469** 0.187 −0.456** −0.358* 坡位 0.282 −0.386* 0.256 0.328 −0.411* −0.217 说明:*和**分别表示在P<0.05和P<0.01下显著相关
下载: 导出CSV
-
[1] WAN Songze, FU Shenglei, ZHANG Chenlu, et al. Effects of understory removal and litter addition on leaf and twig decomposition in a subtropical Chinese fir plantation [J]. Land Degradation &Development, 2021, 32(17): 5004 − 5011. [2] 岳新建, 叶功富, 高伟, 等. 海岸沙地主要森林的凋落物分解及生态化学计量特征[J]. 水土保持研究, 2021, 28(4): 77 − 83. YUE Xinjian, YE Gongfu, GAO Wei, et al. Characteristics of litter decomposition and ecological stoichiometry of different forests on coastal sandy land in Fujian Province [J]. Research of Soil and Water Conservation, 2021, 28(4): 77 − 83. [3] SHEN Ying, TIAN Dashuan, HOU Jihua, et al. Forest soil acidification consistently reduces litter decomposition irrespective of nutrient availability and litter type [J]. Functional Ecology, 2021, 35(12): 2753 − 2762. [4] 李汶倬, 高扬, 杨柳, 等. 东北地区落叶松新鲜针叶凋落物碳氮磷化学计量特征对环境变化的响应[J]. 生态学杂志, 2020, 39(9): 2832 − 2841. LI Wenzhuo, GAO Yang, YANG Liu, et al. Carbon, nitrogen, and phosphorus stoichiometry of recently senesced larch leaves in response to environmental factors across an entire growing season [J]. Chinese Journal of Ecology, 2020, 39(9): 2832 − 2841. [5] 杨关吕, 黎建强, 左嫚, 等. 滇中高原磨盘山云南松林凋落物输入动态及养分归还量研究[J]. 生态环境学报, 2019, 28(11): 2158 − 2164. YANG Guanlü, LI Jianqiang, ZUO Man, et al. Litter production variation and nutrient return of Pinus yunnanensis forest in Mopan mountain in central Yunnan plateau [J]. Ecology and Environmental Sciences, 2019, 28(11): 2158 − 2164. [6] 李非凡, 孙冰, 裴男才, 等. 粤北3种林分凋落叶-根系-土壤生态化学计量特征[J]. 浙江农林大学学报, 2020, 37(1): 18 − 26. LI Feifan, SUN Bing, PEI Nancai, et al. Characteristics of litter-root-soil ecological stoichiometry of three forest stands in northern Guangdong [J]. Journal of Zhejiang A&F University, 2020, 37(1): 18 − 26. [7] CHAPIN Ⅲ F, SMATSON P A, MOONEY H A. Principles of Terrestrial Ecosystem Ecology[M]. New York: Springer, 2011: 183 − 228. [8] 马文济, 赵延涛, 张晴晴, 等. 浙江天童常绿阔叶林不同演替阶段地表凋落物的C∶N∶P化学计量特征[J]. 植物生态学报, 2014, 38(8): 833 − 842. MA Wenji, ZHAO Yantao, ZHANG Qingqing, et al. C∶N∶P toichiometry in forest floor litter of evergreen broad-leaved forests at different successional stages in Tiantong, Zhejiang, eastern China [J]. Chinese Journal of Plant Ecology, 2014, 38(8): 833 − 842. [9] NAVE L E, VANCE E D, SWANSTON C W, et al. Impacts of elevated N inputs on north temperate forest soil C storage, C/N, and net N-mineralization [J]. Geoderma, 2009, 153(1/2): 231 − 240. [10] 王新源, 赵学勇, 李玉霖, 等. 环境因素对干旱半干旱区凋落物分解的影响研究进展[J]. 应用生态学报, 2013, 24(11): 3300 − 3310. WANG Xinyuan, ZHAO Xueyong, LI Yulin, et al. Effects of environmental factors on litter decomposition in arid and semi-arid regions: a review [J]. Chinese Journal of Applied Ecology, 2013, 24(11): 3300 − 3310. [11] CORNELISSEN J H C. An experimental comparison of leaf decomposition rates in a wide range of temperate plant species and types [J]. Journal of Ecology, 1996, 84(4): 573 − 582. [12] 武亚楠, 喻理飞, 张丽敏, 等. 喀斯特高原区植被恢复过程中土壤碳特征及其影响因素[J]. 生态环境学报, 2020, 29(10): 1935 − 1942. WU Yanan, YU Lifei, ZHANG Limin, et al. Characteristics and influencing factors of soil carbon poolduring vegetation restoration in karst plateau [J]. Ecology and Environmental Sciences, 2020, 29(10): 1935 − 1942. [13] 胡忠良, 潘根兴, 李恋卿, 等. 贵州喀斯特山区不同植被下土壤C、N、P含量和空间异质性[J]. 生态学报, 2009, 29(8): 4187 − 4195. HU Zhongliang, PAN Genxing, LI Lianqing, et al. Changes in pools and heterogeneity of soil organic carbon, nitrogen and phosphorus under different vegetation types in karst mountainous area of central Guizhou Province, China [J]. Acta Ecologica Sinica, 2009, 29(8): 4187 − 4195. [14] 刘娜, 喻理飞, 赵庆, 等. 喀斯特高原石漠化区次生林叶片—枯落物—土壤连续体碳氮磷生态化学计量特征[J]. 应用与环境生物学报, 2020, 26(3): 681 − 688. LIU Na, YU Lifei, ZHAO Qing, et al. C∶N∶P stoichiometry of leaf-litter-soil continuum in secondary forests of the rocky desert regions of the karst plateau [J]. Chinese Journal of Applied and Environmental Biology, 2020, 26(3): 681 − 688. [15] 吴鹏, 崔迎春, 赵文君, 等. 喀斯特森林植被自然恢复过程中土壤化学计量特征[J]. 北京林业大学学报, 2019, 41(3): 80 − 92. WU Peng, CUI Yingchun, ZHAO Wenjun, et al. Characteristics of soil stoichiometric in natural restoration process of Maolan karst forest vegetation, southwestern China [J]. Journal of Beijing Forestry University, 2019, 41(3): 80 − 92. [16] 喻阳华, 李一彤, 王俊贤, 等. 贵州白云岩地区植物群落叶片-凋落物-土壤化学计量与碳氮同位素特征[J]. 生态学报, 2022, 42(8): 1 − 10. YU Yanghua, LI Yitong, WANG Junxian, et al. Leaf-litter-soil stoichiometry and carbon and nitrogen isotopes of plant communities in dolomite district in Guizhou Province [J]. Acta Ecologica Sinica, 2022, 42(8): 1 − 10. [17] 曾昭霞, 刘孝利, 王克林, 等. 桂西北喀斯特区原生林与次生林凋落物及养分归还特征比较[J]. 生态环境学报, 2010, 19(1): 146 − 151. ZENG Zhaoxia, LIU Xiaoli, WANG Kelin, et al. Comparison of litterfall and nutrients return properties of primary and secondary forest ecosystems, the karst region of northwest Guangxi [J]. Ecology and Environmental Sciences, 2010, 19(1): 146 − 151. [18] 卢晓强, 杨万霞, 丁访军, 等. 茂兰喀斯特地区原始林凋落物量动态与养分归还[J]. 生态与农村环境学报, 2014, 30(5): 614 − 619. LU Xiaoqiang, YANG Wanxia, DING Fangjun, et al. Dynamics of litterfall and nutrient recycling in virgin forest in Maolan karst region [J]. Journal of Ecology and Rural Environment, 2014, 30(5): 614 − 619. [19] 杨艳鲜, 冯光恒, 潘志贤, 等. 干热河谷罗望子人工林凋落物分解及养分释放[J]. 干旱区资源与环境, 2013, 27(1): 102 − 107. YANG Yanxian, FENG Guangheng, PAN Zhixian, et al. Decomposition dynamics and nutrient release of litters of Tamarindus indica Linn. artificial forests in dry-hot valler, China [J]. Journal of Arid Land Resources and Environment, 2013, 27(1): 102 − 107. [20] BERG B. Plant Litter: Decomposition, Humus Formation, Carbon Sequestration[M]. Berlin: Springer Verlag, 2005: 269 − 271. [21] 马祥庆, 刘爱琴, 何智英. 杉木幼林生态系统凋落物及其分解作用研究[J]. 植物生态学报, 1997, 21(6): 77 − 83. MA Xiangqing, LIU Aiqin, HE Zhiying. The litter and its decomposition in young Chinese fir plantation ecosystem [J]. Chinese Journal of Plant Ecology, 1997, 21(6): 77 − 83. [22] 施昀希, 陈奇伯, 黎建强. 滇中高原森林凋落物不同分解阶段C、N、P的化学计量特征及种间差异[J]. 中南林业科技大学学报, 2020, 40(1): 114 − 122. SHI Yunxi, CHEN Qibo, LI Jianqiang. C, N and P stoichiometry at different stages of litter decomposition and interspecies comparison in central Yunnan plateau [J]. Journal of Central South University of Forestry &Technology, 2020, 40(1): 114 − 122. [23] 喻林华, 方晰, 项文化, 等. 亚热带4种林分类型枯落物层和土壤层的碳氮磷化学计量特征[J]. 林业科学, 2016, 52(10): 10 − 21. YU Linhua, FANG Xi, XIANG Wenhua, et al. Stoichiometry of carbon, nitrogen, andphosphorus in litter and soil of four types of subtropical stand [J]. Scientia Silvae Sinicae, 2016, 52(10): 10 − 21. [24] 龙健, 赵畅, 张明江, 等. 不同坡向凋落物分解对土壤微生物群落的影响[J]. 生态学报, 2019, 39(8): 2696 − 2704. LONG Jian, ZHAO Chang, ZHANG Mingjiang, et al. Effect of litter decomposition on soil microbes on different slopes [J]. Acta Ecologica Sinica, 2019, 39(8): 2696 − 2704. [25] 赵畅, 龙健, 李娟, 等. 茂兰喀斯特原生林不同坡向及分解层的凋落物现存量和养分特征[J]. 生态学杂志, 2018, 37(2): 295 − 303. ZHAO Chang, LONG Jian, LI Juan, et al. Litter stock and nutrient characteristics of decomposing litter layers in Maolan karst primary forest in different slope directions [J]. Chinese Journal of Ecology, 2018, 37(2): 295 − 303. [26] SULKAVA P, HUHTA V. Effects of hard frost and freeze-thaw cycles on decomposer communities and N mineralisation in boreal forest soil [J]. Applied Soil Ecology, 2003, 22(3): 225 − 239. [27] 王书丽, 黄立君, 袁希, 等. 氮添加和升温对杉木林凋落物分解及碳氮磷化学计量特征的影响[J]. 生态学杂志, 2020, 39(9): 2842 − 2850. WANG Shuli, HUANG Lijun, YUAN Xi, et al. Effects of nitrogen addition and warming on litter decomposition and C, N and P stoichiometric characteristics of Chinese fir [J]. Chinese Journal of Ecology, 2020, 39(9): 2842 − 2850. [28] 刘瑞龙, 杨万勤, 谭波, 等. 土壤动物对川西亚高山和高山森林凋落叶第1年不同分解时期N和P元素动态的影响[J]. 植物生态学报, 2013, 37(12): 1080 − 1090. LIU Ruilong, YANG Wanqin, TAN Bo, et al. Effects of soil fauna on N and P dynamics at different stages during the first year of litter decomposition in subalpine and alpine forests of western Sichuan [J]. Chinese Journal of Plant Ecology, 2013, 37(12): 1080 − 1090. [29] 谌贤, 刘洋, 邓静, 等. 川西亚高山森林凋落物不同分解阶段碳氮磷化学计量特征及种间差异[J]. 植物研究, 2017, 37(2): 216 − 226. SHEN Xian, LIU Yang, DENG Jing, et al. C, N and P stoichiometry at different stages of litter decomposition insubalpine forest of western Sichuan Province and interspecies comparison [J]. Bulletin of Botanical Research, 2017, 37(2): 216 − 226. [30] 张建利, 严令斌, 喻理飞, 等. 草海流域3种优势树种凋落物叶分解历程中的水文特征[J]. 水土保持研究, 2014, 21(6): 61 − 66. ZHANG Jianli, YAN Lingbin, YU Lifei, et al. Research for leaf-litter hydrographic effect in the decomposition process of three dominant three species in the Caohai wetland watershed [J]. Research of Soil and Water Conservation, 2014, 21(6): 61 − 66. [31] CRAINE J M, MORROW C, FIERER N. Microbial nitrogen limitation increases decomposition [J]. Ecology, 2007, 88(8): 2105 − 2113. [32] YUAN Zhiyou, CHEN H Y H. Meta-analysis global trends in senesced-leaf nitrogen and phosphorus [J]. Global Ecology and Biogeography, 2009, 18(5): 532 − 542. [33] MCGRODDY M E, DAUFRESNE T, HEDIN L O. Scaling of C∶N∶P stoichiometry in forests worldwide: implications of terrestrial redfield-type ratios [J]. Ecology, 2004, 85(9): 2390 − 2401. [34] 王晶苑, 王绍强, 李纫兰, 等. 中国4种森林类型主要优势植物的C∶N∶P化学计量学特征[J]. 植物生态学报, 2011, 35(6): 587 − 595. WANG Jingyuan, WANG Shaoqiang, LI Renlan, et al. C∶N∶P stoichiometric characteristics of four forest types’ dominant tree species in China [J]. Chinese Journal of Plant Ecology, 2011, 35(6): 587 − 595. [35] VEEN G F, SUNDQVIST M K, WARDLE D A. Environmental factors and traits that drive plant litter decomposition do not determine home-field advantage effects [J]. Functional Ecology, 2015, 29(7): 981 − 991. [36] 赵溪, 李君剑, 李洪建. 关帝山不同植被恢复类型对土壤碳、氮含量及微生物数量的影响[J]. 生态学杂志, 2010, 29(11): 2102 − 2110. ZHAO Xi, LI Junjian, LI Hongjian. Effects of vegetation restoration type on soil carbon, nitrogen, and microbial quantity in Guandi Mountain [J]. Chinese Journal of Ecology, 2010, 29(11): 2102 − 2110. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20220289
 点击查看大图
点击查看大图

计量
- 文章访问数: 1882
- HTML全文浏览量: 1104
- PDF下载量: 89
- 被引次数: 0
