







-
Wuschel-related homeobox (WOX)是植物特有的新型转录因子,属于Homepbox (HOX)超家族,包含由60~65个氨基酸组成的螺旋-环-螺旋-转角-螺旋的保守结构域。WOX家族基因分为3个独立进化支,即现代进化支(modern clade,WUS),中间进化支(intermediate clade)和远古进化支(ancient clade)[1-3],其中WUS是最早发现的WOX家族成员[4]。WOXs蛋白在植物胚胎形成[5]、干细胞维持[6]和花发育[7]等方面发挥重要作用。拟南芥Arabidopsis thaliana中WOX家族有15个成员,分别是AtWUS和AtWOX1~AtWOX14[4],其中,AtWOX10、AtWOX13和AtWOX14蛋白属于远古进化分支,AtWUS和AtWOX1~7等8个蛋白质属于WUS分支,AtWOX8、AtWOX9、AtWOX11和AtWOX12蛋白属于中间进化分支。AtWUS在胚珠、花药和茎尖分生组织中表达,是维持中央分生组织的关键基因,AtWOX11和AtWOX12参与新生根器官发生,在顶端分生组织发育阶段,AtWUS参与干细胞稳态维持[8]。超表达AtWUS促进棉花Gossypium hirsutum体细胞胚胎发育和器官发生[9]。水稻Oryza sativa中,OsWOX11激活冠根的萌发和生长,过表达OsWOX11可促进雌蕊增加[10];OsWOX3A参与水稻叶片、小穗、分蘖和侧根的发育[11];在茎顶端分生组织和腋分生组织中OsWOX4正调控干细胞[12]。超表达WOX11 (PeWOX11a和PeWOX11b)或WOX11/12a增加转基因植株不定根数量[13-14]。在小麦Triticum aestivum中超表达TaWUS影响外花轮状器官发育,TaWOX9促进转基因拟南芥根的发育[15]。
WOXs转录因子不仅调控植物生长发育,而且参与胁迫响应。在水稻中OsWOX12A和OsWOX12B等基因的表达受干旱、寒冷和盐胁迫差异调控,超表达OsWOX11通过促进根毛生长发育提高转基因植株干旱胁迫耐受性[16-17]。84K杨树Populus alba×P. glandulosa中,干旱胁迫诱导PagWOX11/12a基因强烈表达,促进根系伸长和生物量生长,上游调控因子PagERF35激活PagWOX11/12a表达[18],PagWOX11/12a通过调控PagCYP736A12基因表达,调节活性氧(reactive oxygen species,ROS )清除,提高杨树耐盐性[19]。
杜仲Ecommia ulmoides是杜仲科Eucommiaceae杜仲属Eucommia的落叶乔木,为中国二级保护植物,叶片、树皮和果皮中富含杜仲胶,是重要的胶用和药用经济树种,具有极高的开发利用价值[20]。杜仲叶片中含有绿原酸、黄酮类、木脂素类、环烯醚萜类、α-亚麻酸等药用成分,具有抗疲劳、抗衰老、抗肿瘤、增强免疫力等重要作用[21-22]。鉴于WOX基因在拟南芥、水稻、玉米Zea mays、杨树、油菜Brassica napus、铁皮石斛Dendrobium officinale等中的作用,推测WOX家族基因可能在杜仲叶芽的形成和激活过程中起关键作用。本研究以杜仲基因组数据为基础,对杜仲WOX家族基因进行了全基因组鉴定和生物信息学分析,基于转录组分析WOX在杜仲叶片不同发育时期以及杜仲胶形成中的表达模式,利用实时荧光定量PCR(RT-qPCR)检测杜仲WOX基因(EuWOXs)在‘紫叶’杜仲‘Ziye’叶片发育中的表达水平,以期为EuWOXs功能的深入研究奠定基础。
-
自西北农林科技大学苗圃(陕西杨凌),取生长正常长势一致的2年生‘紫叶’杜仲幼苗的叶芽(茎尖)、生长叶(3 cm长叶片)、幼叶(完全展开的新叶),用液氮迅速处理,置于−80 ℃冰箱保存。
-
拟南芥WOX蛋白序列下载于TAIR数据库,根据Pfam号(PF00046)在杜仲基因组数据库中筛选出WOX家族基因候选序列,利用美国国立生物技术信息中心(National Center for Biotechnology Information,NCBI)的保守结构域搜索服务(Conserved Domain Search Service,CD Search)检测蛋白质保守结构域,筛选出具有完整WOX结构域的蛋白质作为EuWOX家族成员,利用生物信息学方法[23]分析EuWOXs的理化性质。
-
通过杜仲基因组数据库搜索WOX基因在染色体上的位置及每条染色体长度,利用MapGene2Chromosome v2 (
http://mg2c.iask.in/mg2c_v2.0/ )软件绘制WOX家族基因染色体定位。利用DNAMAN进行蛋白序列比对,通过Clustal X 1.83对杜仲、拟南芥、毛果杨、水稻和玉米WOXs蛋白进行多序列比对,利用MEGA 6.0邻接法(neighbor-joining),重复次数设置为1 000次[24],构建系统发育树,根据拟南芥同源基因对EuWOXs蛋白命名。 -
利用GSDS (
http://gsds.gao-lab.org/index.php )软件分析EuWOXs的内含子和外显子分布。利用MEME (http://meme-suite.org/ )软件分析EuWOXs基序,参数设置为:any number of Repetitions,maximum number of Motifs=20,minimum width≥6,and maximum width≤50。分离EuWOXs启动子(ATG)上游2 000 bp序列,利用Plant CARE 分析EuWOXs启动子顺式作用元件。 -
从NCBI的Short Read Arshive (SRA)数据库中下载‘秦仲1号’‘Qinzhong No.1’叶片不同发育时期(叶芽、初生叶、幼叶、老叶)(版本号:SRP218063)[25]及高产胶杜仲品种‘秦仲2号’‘Qinzhong No.2’、低产胶杜仲品种‘小叶’‘Xiaoye’(版本号:SRP158357)[26]的转录组数据,使用每1百万个映射上的碱基中映射到外显子的每1千个碱基上的碱基个数(fragments per kilobase million,FPKM)值表示EuWOXs相对表达丰度,取对数(log2)进行统计分析,利用TBtools工具绘制基因表达图谱[27]。
使用Trizol(天根DP424)提取RNA,反转录成cDNA,通过Primer 3.0软件设计EuWOXs特异性引物(引物序列见表1),通过Quant Studio 6(新加坡Life Technologies公司),All-in-One SYBR Premix EX TaqTM kit(美国Gene Copoeia公司)进行实时荧光定量PCR(RT-qPCR)反应,10 μL反应体系:2× mix 5.00 μL、正向引物/反向引物各0.25 μL、cDNA 2.00 μL、ROX 0.20 μL、双蒸水2.30 μL。反应程序:95 ℃预变性5 min,95 ℃变性10 s,60 ℃退火10 s,72 ℃延伸20 s,45个循环。以UBCE2为内参基因[26],通过$2^{-\Delta \Delta C_t} $法对3次生物学重复进行数据分析。
表 1 引物序列
Table 1. Primer sequences
基因名 上游引物(5′→3′) 下游引物(5′→3′) EuWOX1 ATGGTGGGTGACCAGCTTAG TTCTCTGGCCTTGTGGTTCT EuWOX2 ACCGTACCCCAACCTACTCC ACTTCCCGTTGGATGAAGTG EuWOX4-1 GGAACCCTACGCAAGAACAG GCGCTTCTGCTTTTGTCTCT EuWOX4-2 TAGAGCAGATCACGGCACAG CTAGGGTCGGATGTTGGAGA EuWOX5 GACGGAGCAAGTGAGAGTCC TCTCCCGTGCCTTATGATTC EuWOX11 ACTCGAGTTTTGTGGCCTGT AATTGGAGGCATCTGGATTG EuWOX13-1 GGTCTGAGGGCATGTGTTTT TTGGAGATATGGGTGGTGGT EuWOX13-2 GGGTTGTTCGTCAAGGTCAT GTTGGAATCCACCGTTGTCT UBCE2 AGTGGGTGGTGCTGTAGTCC AACTCCCGTTTCGTTTGTTG -
通过STRING软件(
https://string-db.org/ )上传EuWOXs蛋白质序列,利用拟南芥数据库,根据拟南芥WOXs蛋白已知互作关系,预测EuWOXs互作蛋白,通过Cytoscape 3.7.0软件对EuWOXs蛋白质互作信息进行评估和预测[28]。 -
由表2可知:从杜仲基因组中共鉴定到8个EuWOXs,分布在8条染色体上(图1);均含有HD保守结构域,其中EuWOX1序列最长,编码352个氨基酸,EuWOX13-2序列最短,编码191个氨基酸。EuWOXs分子量为22.12~40.36 kDa,EuWOX11等电点最小,为5.62,EuWOX4-1等电点最大,为9.04;亚细胞定位预测结果显示:EuWOXs均定位在细胞核中,均为亲水性蛋白。
表 2 EuWOXs蛋白质序列特征及亚细胞定位
Table 2. Sequence characteristics and subcellular location of E. ulmoides WOX proteins
基因号 基因名 拟南芥
同源基因染色体 位置 CDS长度/
bp氨基酸数/
个分子量/
kDa等电点 亚细胞
定位EUC13591-RA EuWOX1 AT3G18010.1 Super-Scaffold_235 3540694-3544292 1059 352 40.36 5.78 细胞核 EUC12552-RA EuWOX2 AT5G59340.1 Scaffold912_obj 156744-159059 810 269 30.16 8.11 细胞核 EUC15721-RA EuWOX4-1 AT1G46480.1 Super-Scaffold_242 604979-606289 618 205 23.48 9.04 细胞核 EUC21176-RA EuWOX4-2 AT1G46480.1 Scaffold272_obj 37477-39280 642 213 24.31 8.82 细胞核 EUC18832-RA EuWOX5 AT3G11260.1 Super-Scaffold_117 336319-340482 549 182 20.70 6.92 细胞核 EUC00362-RA EuWOX11 AT3G03660.1 Super-Scaffold_154 68808-70507 765 254 27.67 5.62 细胞核 EUC00756-RA EuWOX13-1 AT4G35550.1 Super-Scaffold_233 319468-325733 810 269 30.42 6.22 细胞核 EUC02503-RA EuWOX13-2 AT4G35550.1 Super-Scaffold_71 6332599-6364092 576 191 22.11 6.54 细胞核 
图 1 EuWOXs染色体位点
Figure 1. Chromosome site of EuWOXs genes
-
利用DNAMAN软件对8个EuWOXs及12个拟南芥WOXs蛋白(AtWOXs)保守结构域进行序列分析。结果(图2)显示:WOXs蛋白质HD结构域氨基酸及其分布具有显著的相似性,均包含由60个氨基酸组成的螺旋-环-螺旋-转角-螺旋,螺旋较环和转角保守。谷氨酰胺(Q)、亮氨酸(L)和脯氨酸(Pro)是螺旋Ⅰ (Helix Ⅰ)结构域的保守氨基酸,脯氨酸、异亮氨酸(Ile)和亮氨酸是螺旋Ⅱ (Helix Ⅱ)结构域的保守氨基酸,相比之下,螺旋Ⅲ (Helix Ⅲ)结构域最为保守,其中保守氨基酸有天冬酰胺(N)、缬氨酸(V)、色氨酸(W)、苯丙氨酸(F)、谷氨酰胺、天冬酰胺和精氨酸(R)。EAR-like仅存在于EuWOX1、EuWOX2、EuWOX4-1、EuWOX4-2和EuWOX5中,属于WUS,暗示EuWOXs在进化过程中具有保守性。
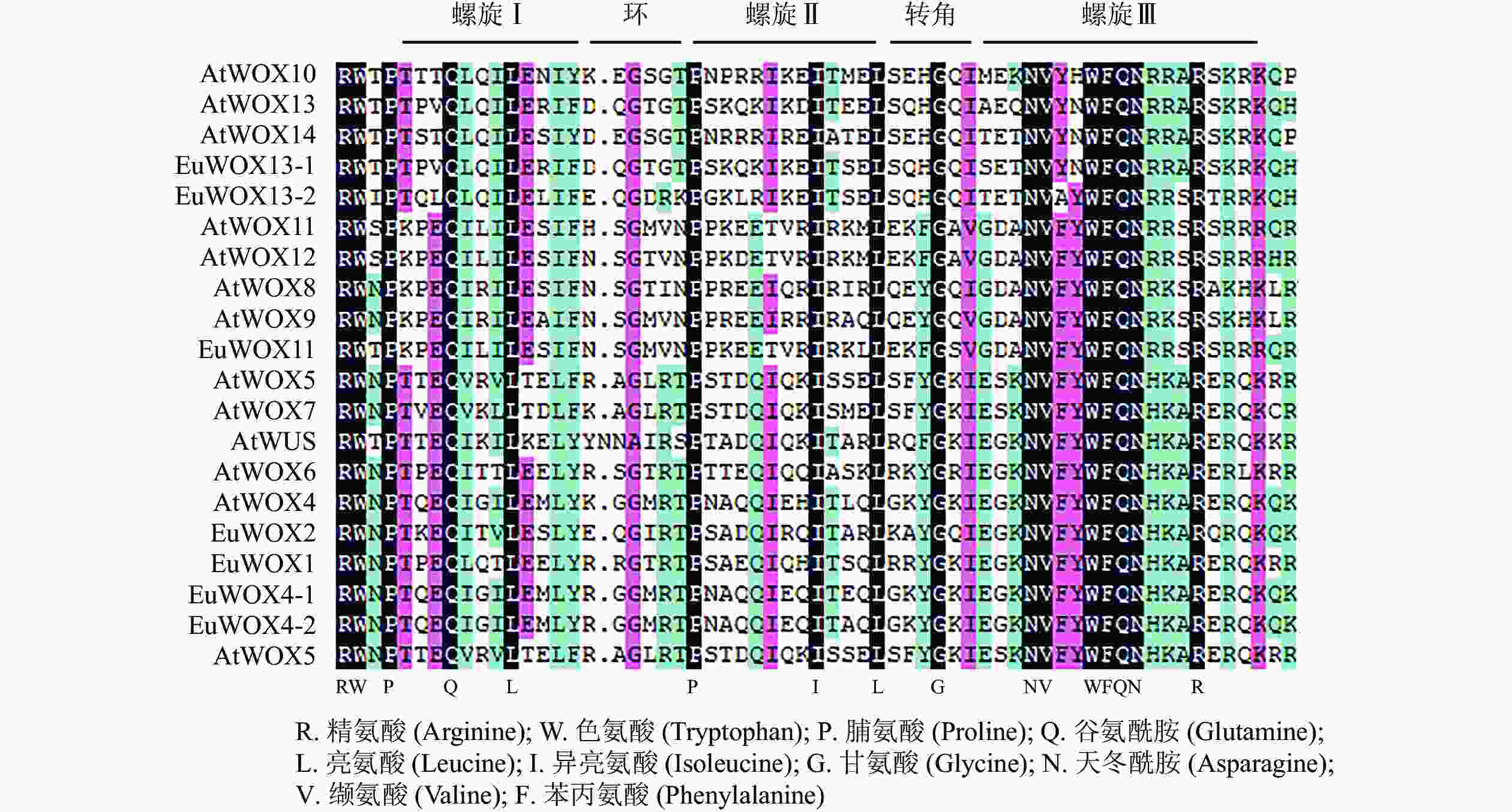
图 2 拟南芥和杜仲WOXs的蛋白质同源结构域序列分析
Figure 2. Sequence analysis of WOX proteins homeo domain in A. thaliana and E. ulmoides
-
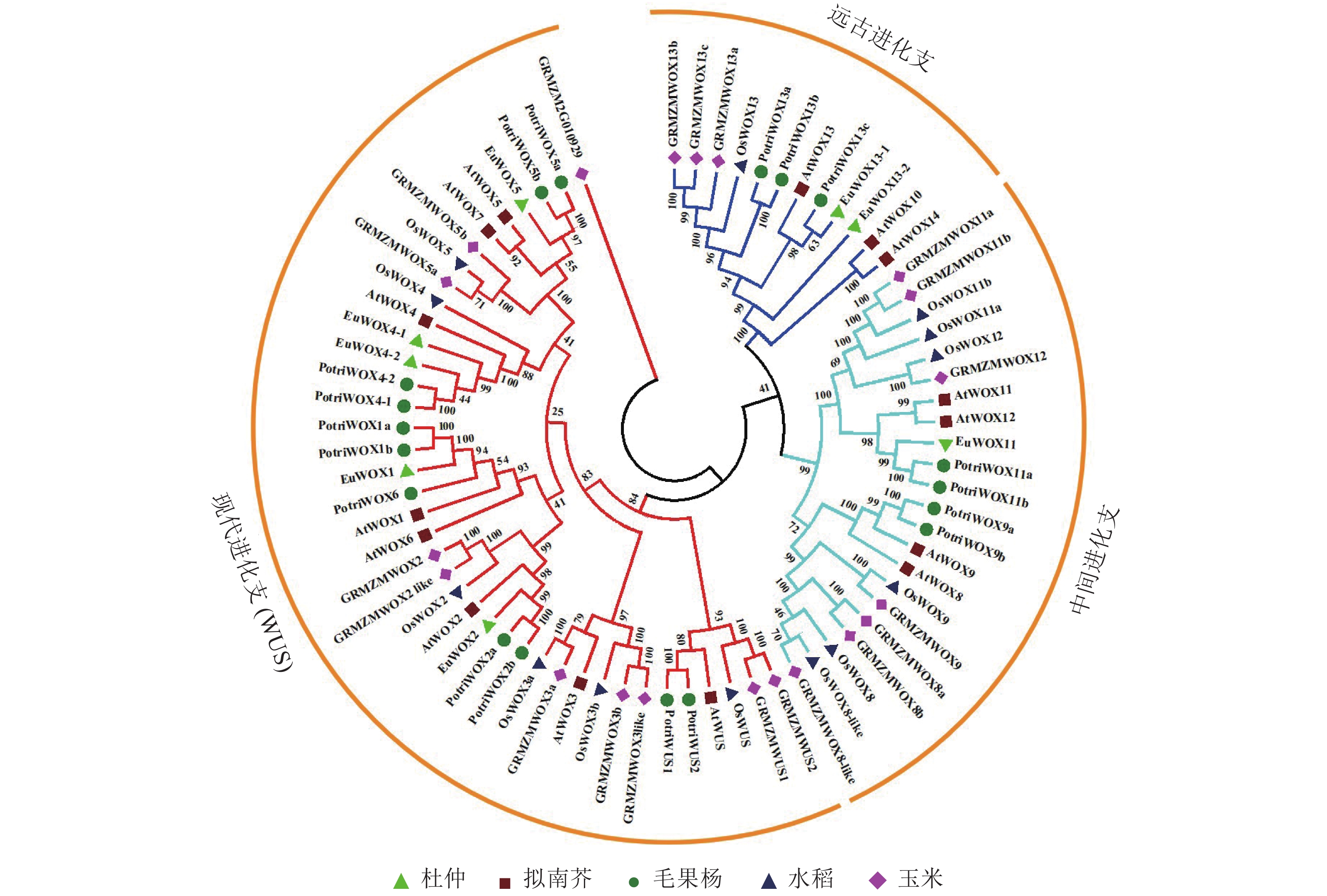
对8个杜仲EuWOXs、15个拟南芥AtWOXs、18个毛果杨Populus trichocarpa WOX蛋白(PotriWOXs)、13个水稻OsWOXs、20个玉米ZmWOXs的蛋白质序列进行多重比对,构建无根系统发育树。结果如 图3所示:74个WOXs蛋白共分为3组[远古进化支、中间进化支和现代进化支(WUS)],其中远古进化支含有12个WOXs蛋白,中间进化支含22个WOXs蛋白,现代进化支包含的蛋白数量最多,共有40个。8个EuWOXs中,EuWOX13-1和EuWOX13-2属于远古进化支,EuWOX11属于中间进化支,EuWOX1、EuWOX2、EuWOX5、EuWOX4-1和EuWOX4-2等5个蛋白质属于WUS。进化结果显示:杜仲与毛果杨亲缘关系最近。
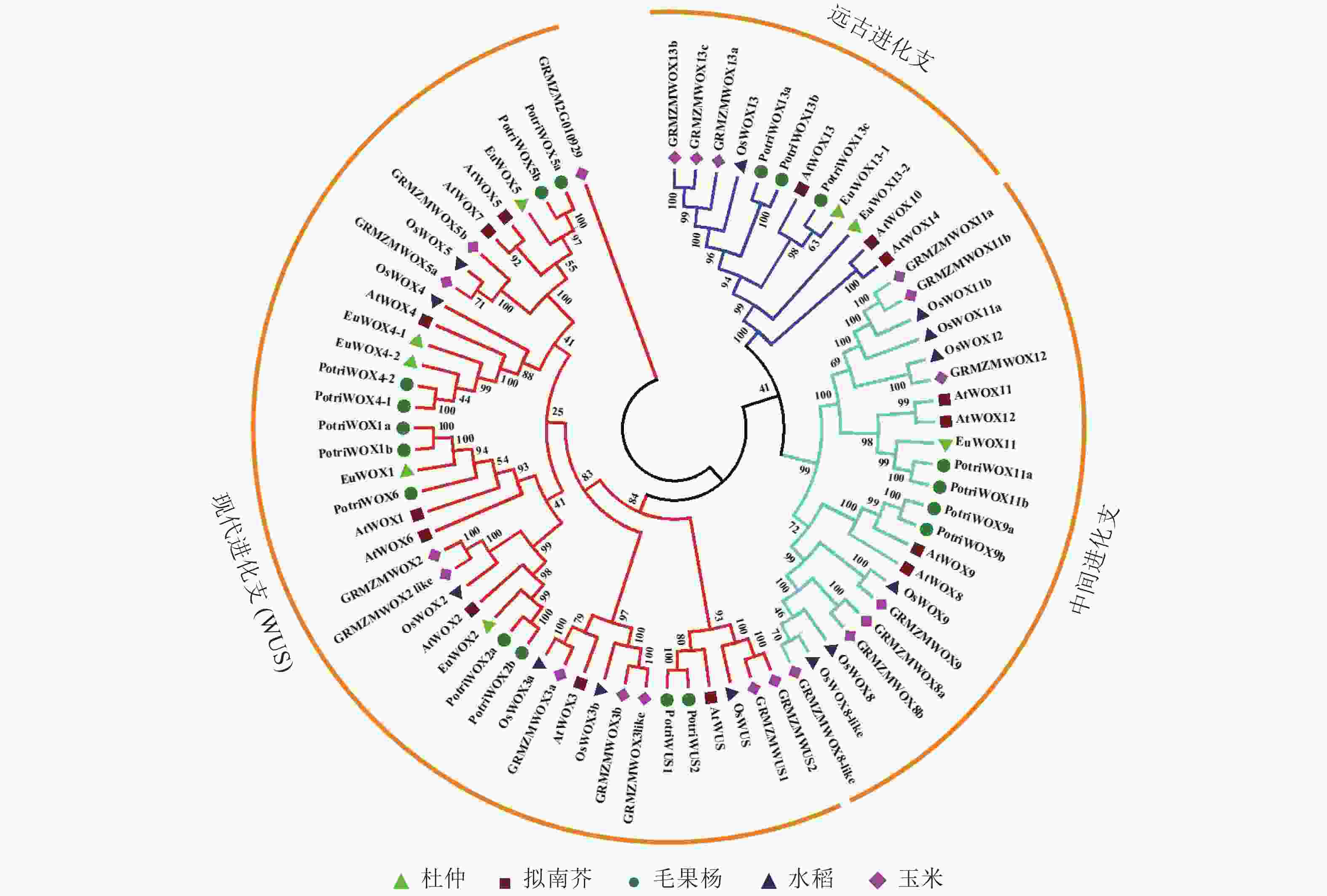
图 3 杜仲、拟南芥、毛果杨、水稻和玉米WOXs蛋白系统发育树
Figure 3. WOX proteins phylogenetic trees of E. ulmoides, A. thaliana, P. trichocar, O. sativa and Z. mays
-
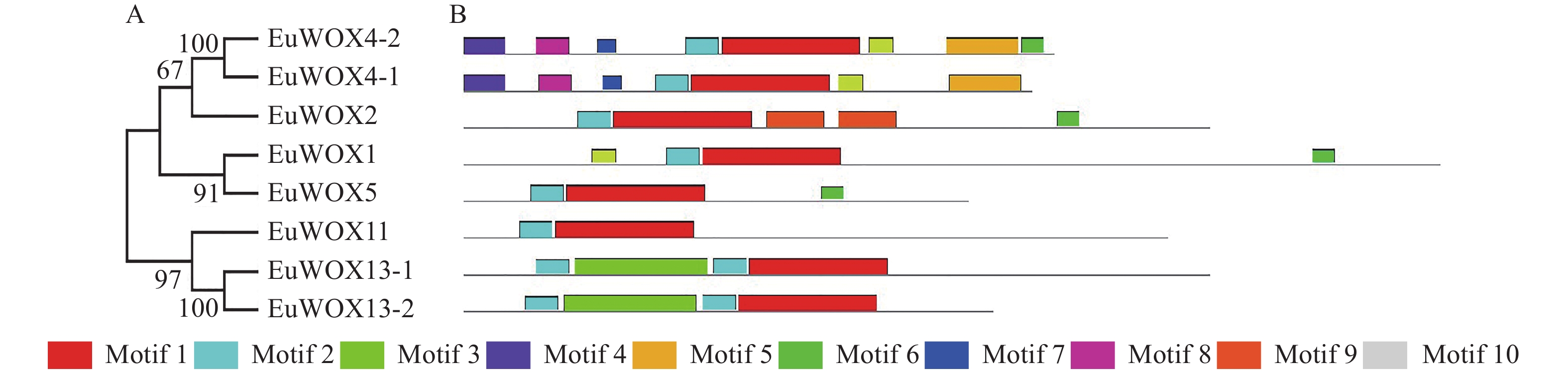
利用GSDS软件构建EuWOXs基因内含子-外显子结构图,结果如图4显示:EuWOXs含有1~3个内含子,EuWOX13-2、EuWOX2和EuWOX5基因含有2个外显子,EuWOX11、EuWOX13-1、EuWOX4-1和EuWOX4-2含有3个外显子,EuWOX1含有4个外显子。不同进化分支基因结构差异显著,同一分支基因结构也存在差异。属于中间进化支的EuWOX11含有3个外显子,同属远古进化支的EuWOX13-1和EuWOX13-2分别含有3个和2个外显子,在WUS中,EuWOX2和EuWOX5含有2个外显子,EuWOX4-1和EuWOX4-2含有3个外显子,而EuWOX1含有4个外显子。
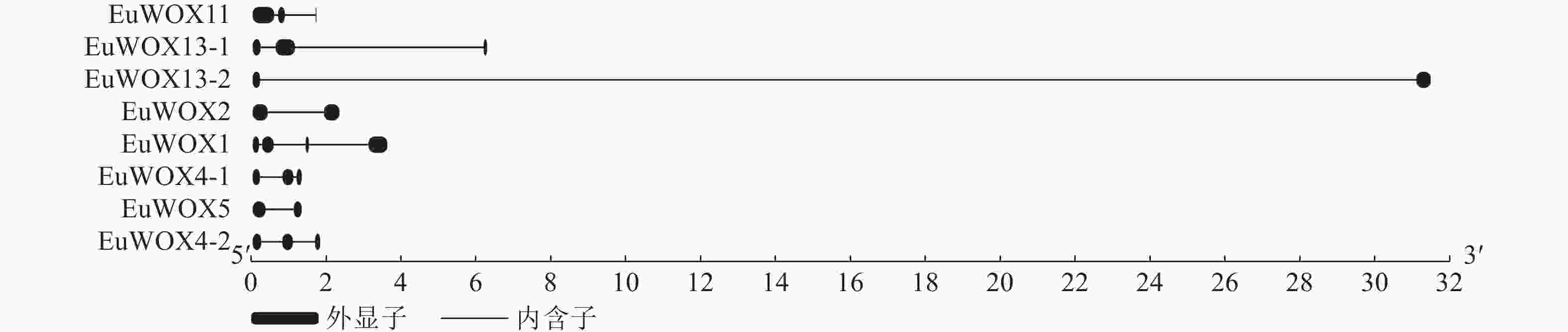
图 4 杜仲WOX家族基因的结构分析
Figure 4. Structural analysis of WOX gene family in E. ulmoides
蛋白质保守基序分析显示:EuWOXs含有10个保守基序,分别命名为Motif 1~Motif 10 (图5),其中Motif 1和Motif 2最为保守,是WOX的核心基序,存在于所有EuWOXs中。Motif 6较为保守,存在于4个EuWOXs蛋白质(EuWOX4-2、EuWOX2、EuWOX1和EuWOX5)中。相同分支EuWOXs含有相似的保守基序,不同分支EuWOXs基序之间存在显著差异,Motif 4~Motif 10只存在于现代进化分支,Motif 3只在EuWOX13-1和EuWOX13-2蛋白质中存在。

图 5 杜仲WOX基因家族保守基序分析
Figure 5. Conserved motifs analysis of E. ulmoides WOX gene family
-
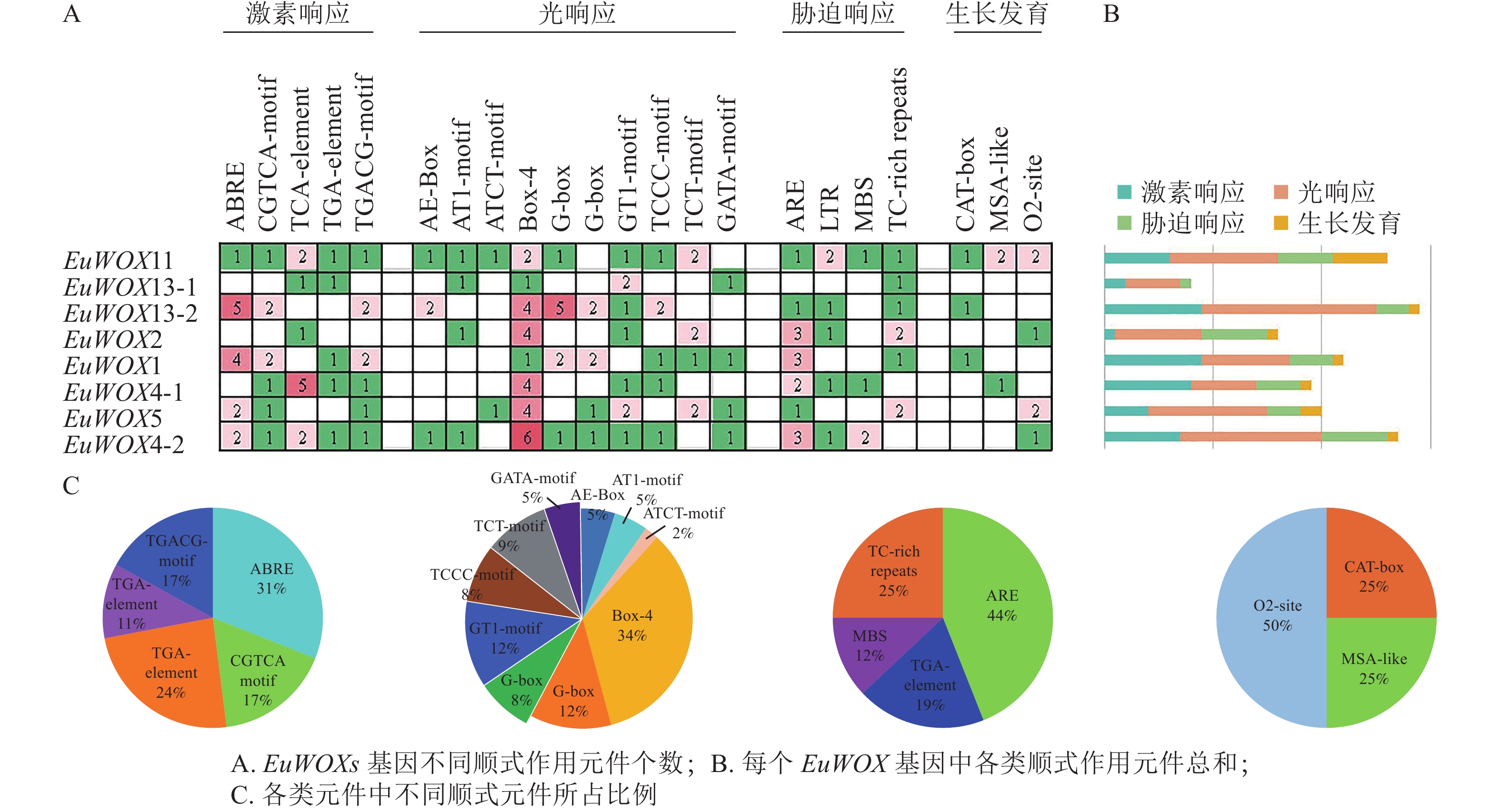
顺式作用元件分析结果(图6)显示:EuWOXs启动子中主要包括脱落酸(ABRE)和水杨酸反应元件(TCA-element)、厌氧响应元件(ARE)、光响应元件(Box 4)及玉米蛋白代谢调节元件(O2-site)。所有顺式作用元件中光响应元件最多,达77个,其中Box 4元件有26个,所占比例是34%;G-box和GT1-motif元件均有9个,占比为12%,表明EuWOXs基因表达可能与光合作用有关。EuWOXs共含有46个激素响应元件,32个胁迫响应元件,其中ABRE和ARE元件数量最多,均含有14个,所占比例分别为31%和44%,暗示EuWOXs参与杜仲激素及胁迫响应。此外EuWOXs共含有12个生长发育调控相关元件,其中O2-site有6个,占50%。
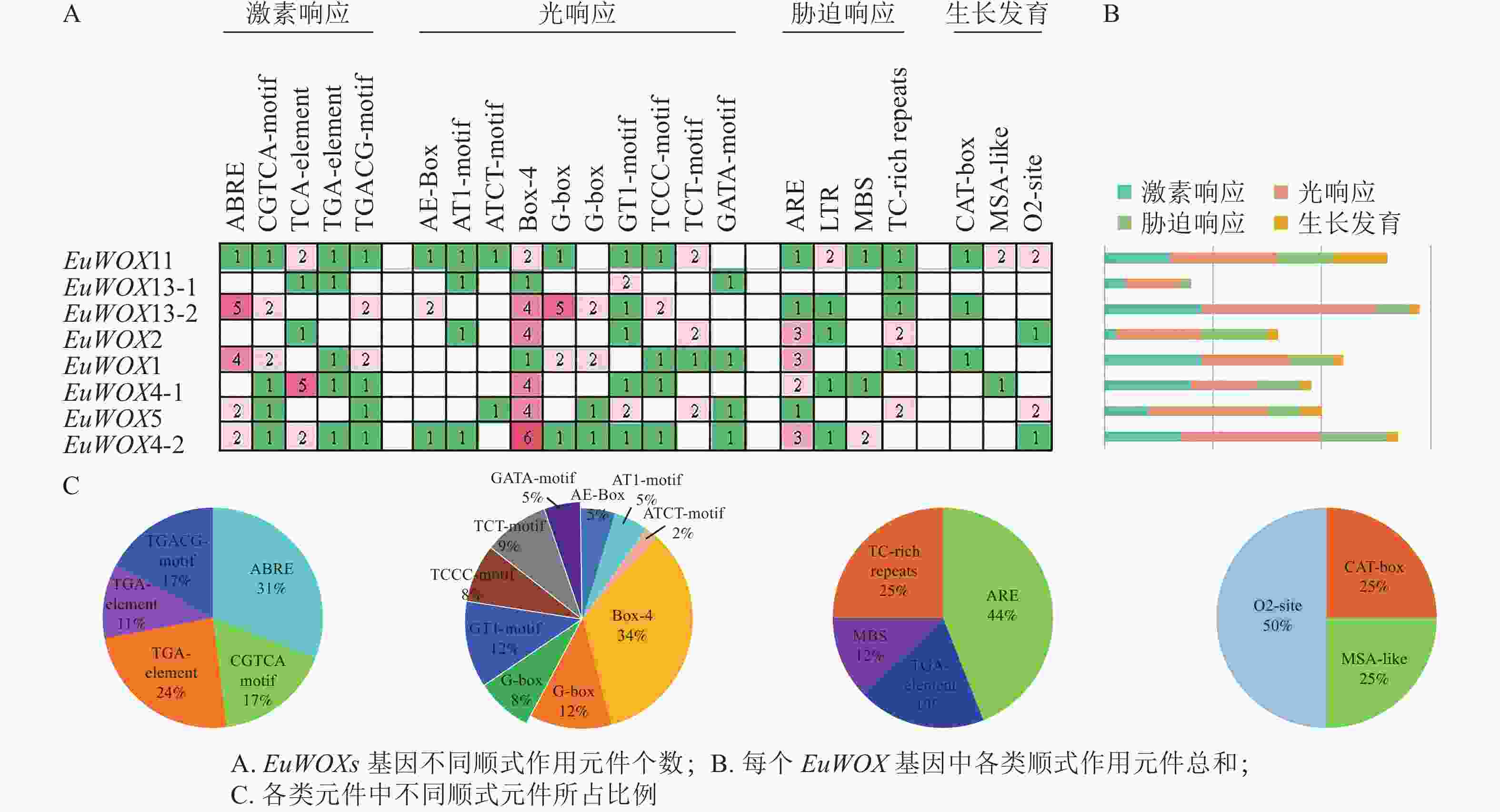
图 6 EuWOXs基因启动子顺式作用元件分析
Figure 6. Cis-element analysis of EuWOX genes promoter
-
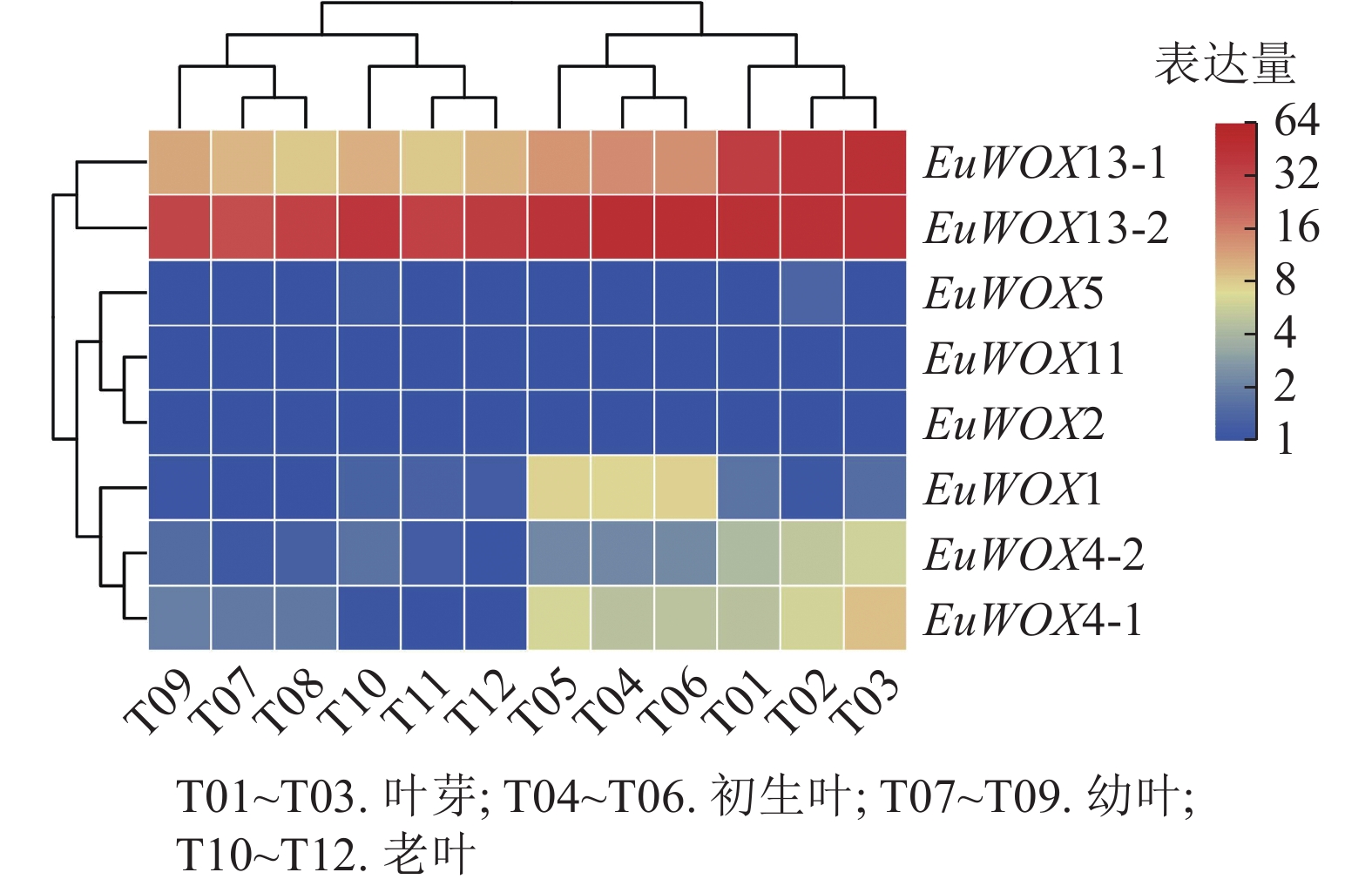
利用‘秦仲1号’叶片不同发育时期转录组数据对EuWOXs基因的表达模式进行分析。结果(图7)可见:EuWOXs在叶片不同发育时期表达丰度存在显著差异,EuWOX11和EuWOX2在杜仲叶芽、初生叶、幼叶、老叶时期均不表达,EuWOX5仅在叶芽和老叶中低表达,EuWOX13-2在4个时期中的FPKM值均大于20,推测EuWOX13-2参与杜仲叶片的整个发育过程;EuWOX13-1和EuWOX4-1在叶芽中表达丰度最高,随着叶片发育表达水平逐渐降低,表明EuWOX13-1和EuWOX4-1主要在叶芽中发挥作用;EuWOX1在生长叶中表达量相对较高,其余EuWOXs基因表达丰度较低,FPKM值小于5。
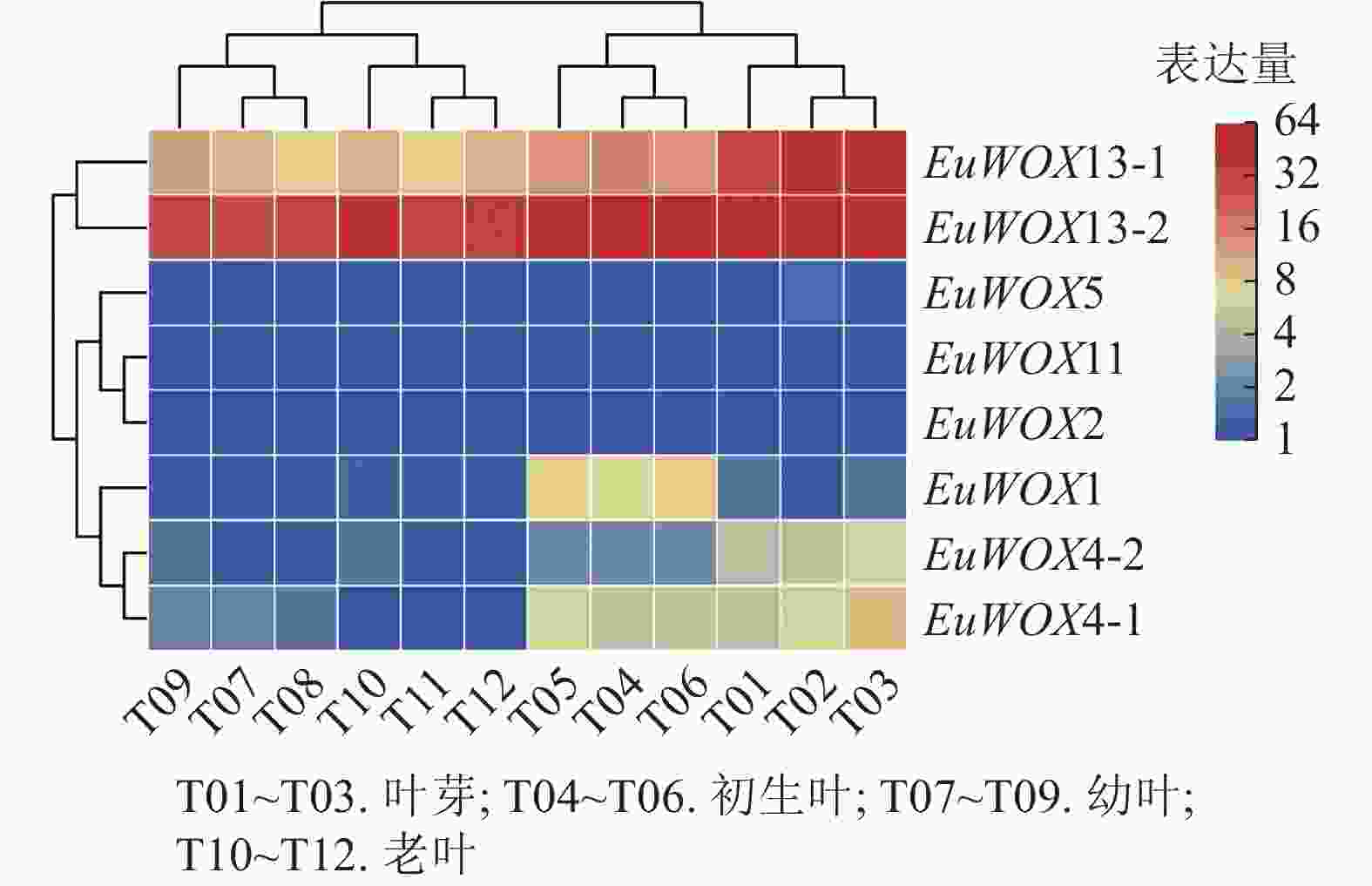
图 7 EuWOXs在杜仲叶片不同发育阶段的表达模式
Figure 7. Expression patterns of EuWOXs at different developmental stages in E. ulmoides leaves
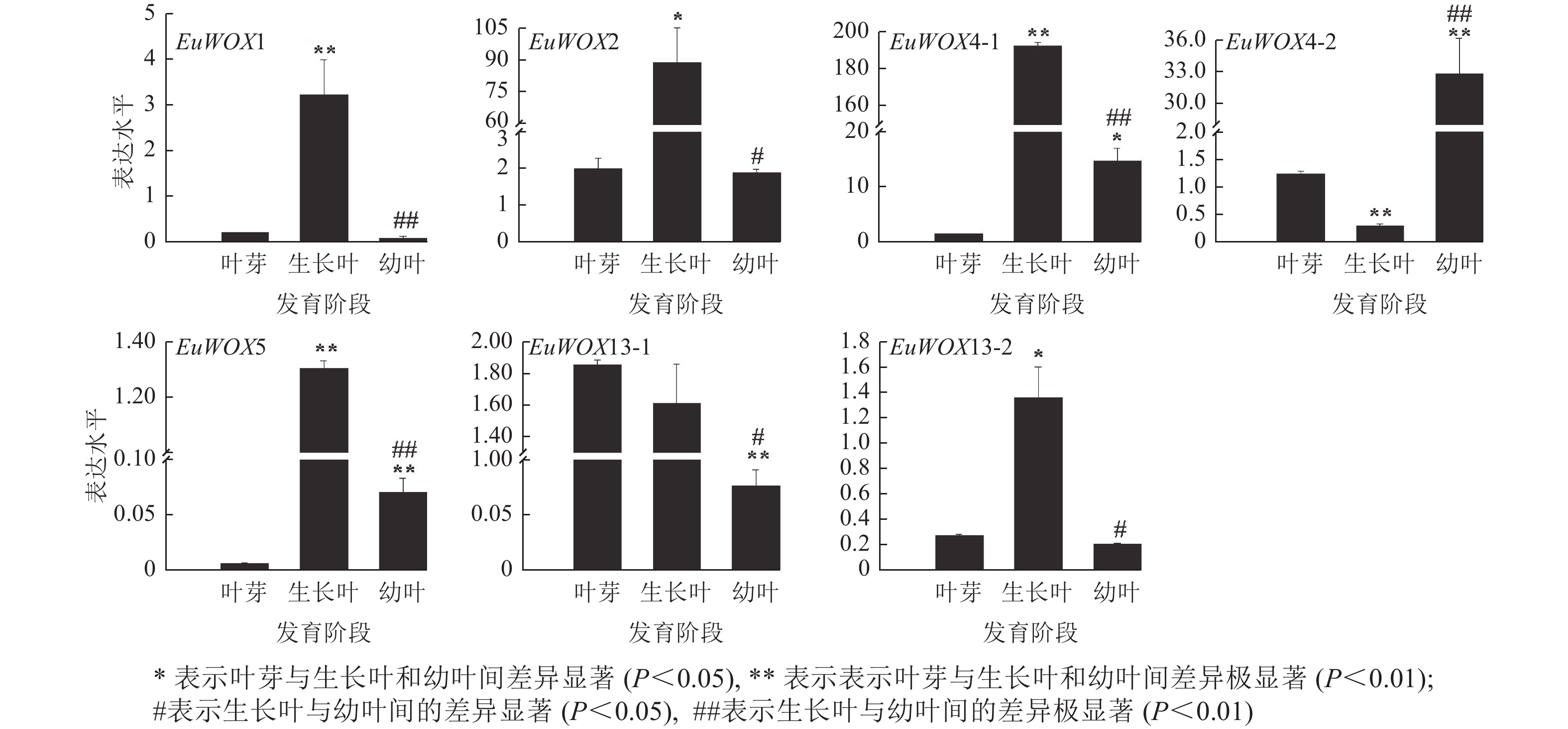
利用RT-qPCR检测EuWOXs在‘紫叶’杜仲叶片不同发育阶段(叶芽、生长叶、幼叶)的表达水平。结果(图8)可见:EuWOX1、EuWOX2、EuWOX4-1、EuWOX5和EuWOX13-2在生长叶中表达量最高,随着叶片发育表达水平呈先升高后降低趋势,EuWOX4-2在幼叶中表达量最高,EuWOX13-1在叶芽中表达量最高,随着叶片发育,表达量逐渐降低,暗示EuWOX13-1在叶片发育的起始阶段发挥重要作用。EuWOX1、EuWOX13-1和EuWOX13-2在‘紫叶’杜仲叶片中的表达趋势与‘秦仲1号’一致。
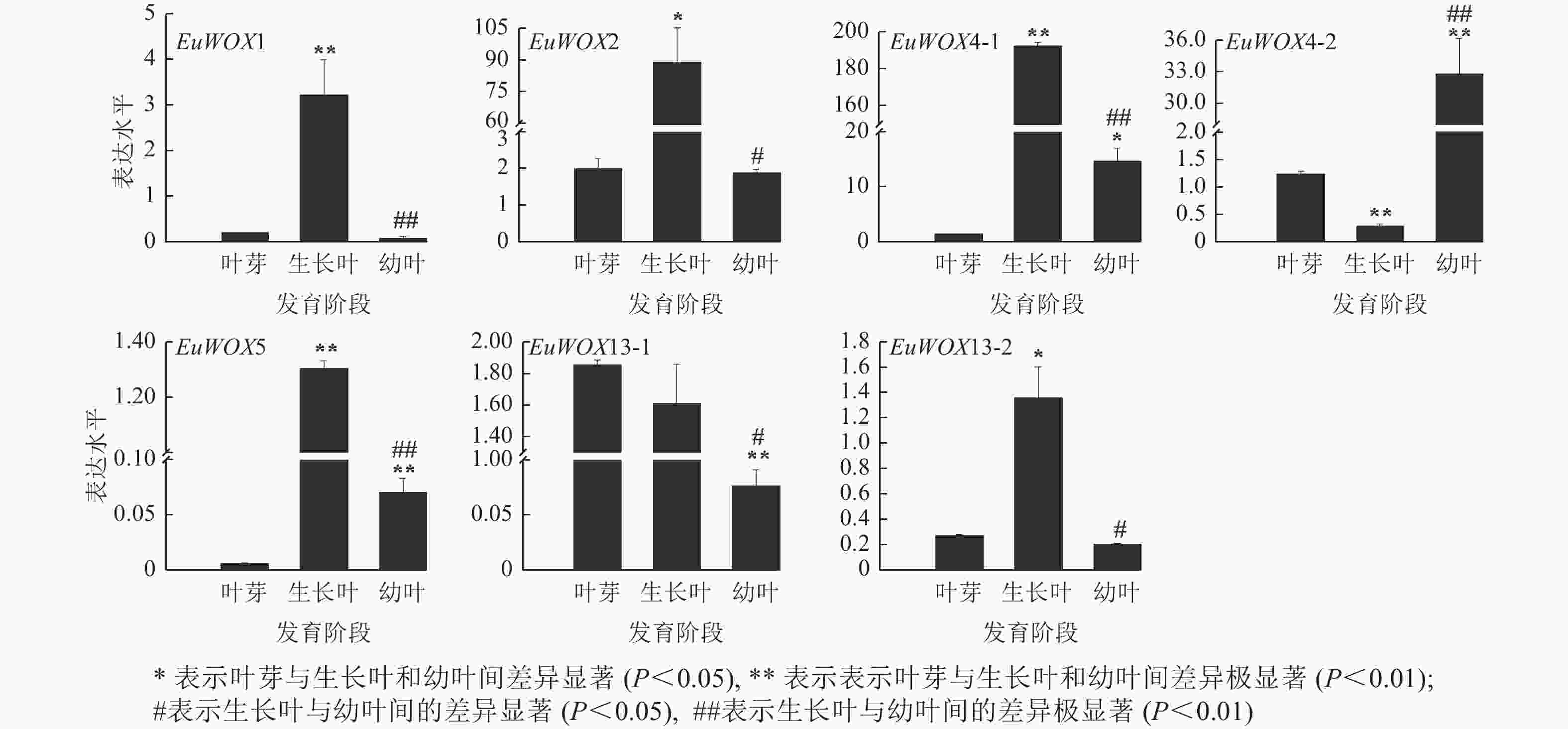
图 8 杜仲EuWOXs基因在杜仲叶片不同发育时期的表达模式
Figure 8. Expression pattern of EuWOX genes in E. ulmoides leaves at different developmental stages
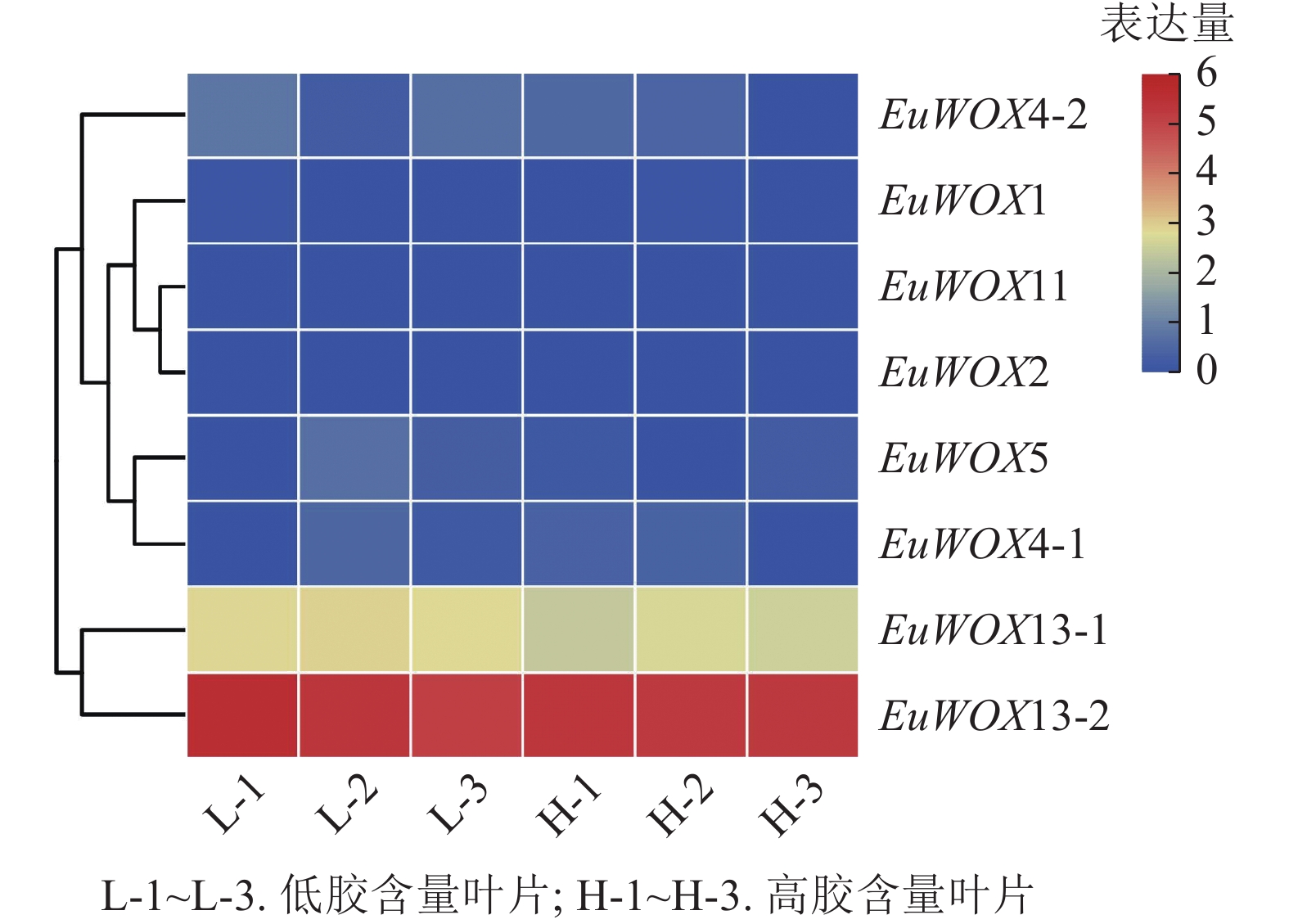
利用‘秦仲2号’和‘小叶’杜仲成熟叶片转录组数据检测EuWOXs的表达模式。由如图9可见:大部分EuWOXs转录水平较低,其中有6个EuWOXs基因几乎不表达,EuWOX13-2表达水平最高,FPKM值>40,不同胶含量样品之间无显著差异,推测EuWOXs在杜仲胶形成过程中发挥作用较小。

图 9 EuWOXs在杜仲胶形成中的表达模式
Figure 9. Expression patterns of EuWOX genes in the form of Eu-rubber
-
植物WOXs蛋白由多基因家族编码,蛋白质之间可能存在相互作用。利用STRING数据库,构建EuWOXs蛋白相互作用网络。图10显示:该网络包含21个节点(互作蛋白)和82条边(相互作用组合)。EuWOX4-1可与20个蛋白质互作,其中包含干细胞分化抑制因子(CLE41和CLE44),细胞增殖和愈伤组织形成蛋白(CLV1、CLV3和ACT7),胚胎发育相关蛋白(TPL、BBM和APM1),维管组织发育蛋白(PXY、HB-8、ATHB-15、MOL和RUL1),细胞程序化死亡调控因子(LOL1),参与DNA的复制和延伸(MCM1、POLD2)以及信号转导蛋白(F14K14),花药发育关键调控因子(RPK2),参与DNA的复制植物发育相关转录因子GRAS(HAM3和SCL27)等,推测EuWOXs全面参与了杜仲的生长发育。
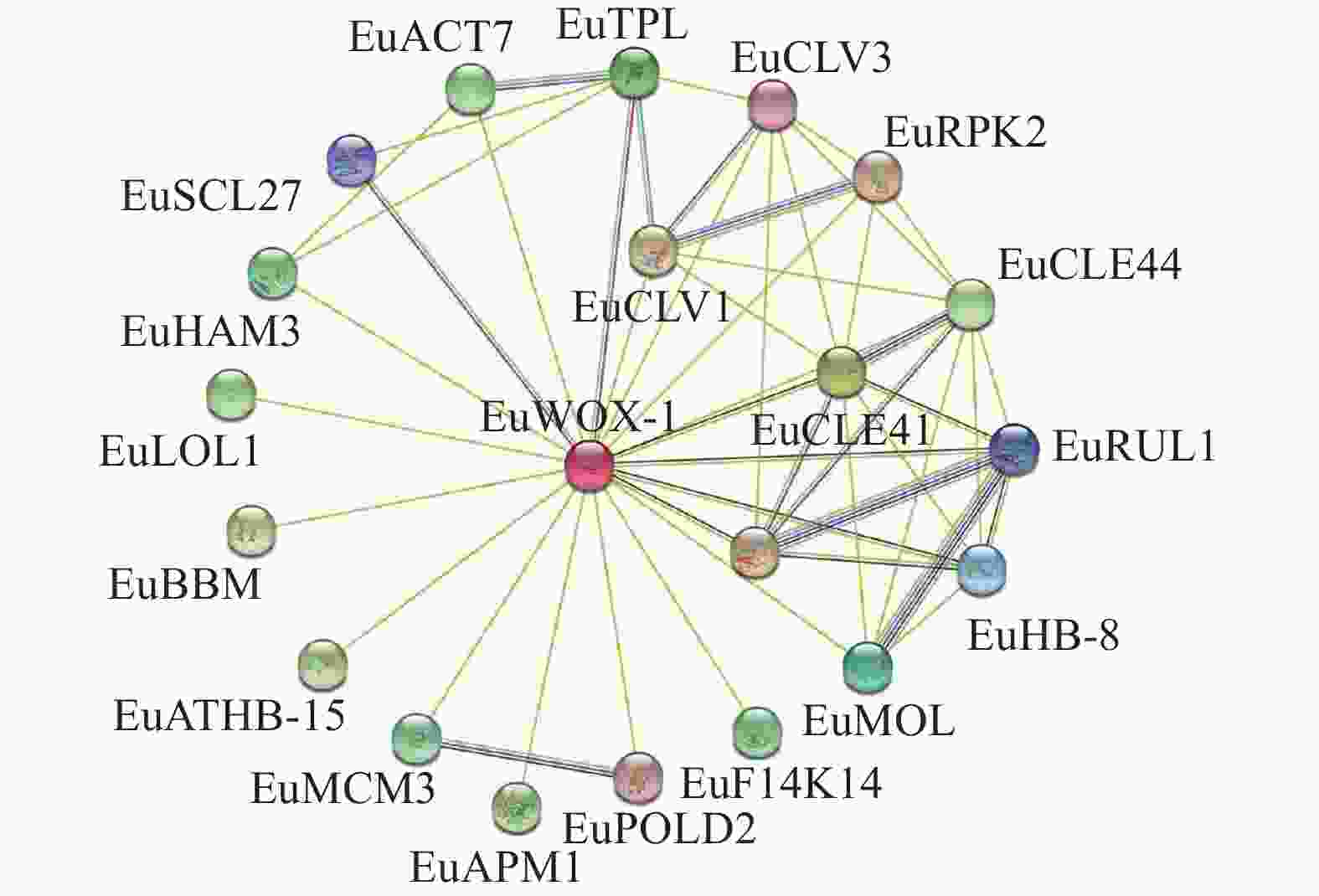
图 10 EuWOXs蛋白互作网络预测
Figure 10. Prediction of interaction network between EuWOX proteins
-
WOX蛋白是植物特有的高度保守的一类转录因子,广泛参与植物的生长发育、干细胞维持、组织器官发生和形成等多种生物学过程。到目前为止,WOX家族基因已在多个物种进行了研究报道,如拟南芥中含有15个、毛果杨18个、水稻中有13个、玉米中有20个、毛竹Phyllostachys edulis 中存在27个[29],小麦中有26个[30]、茶树Camellia sinensis中包含18个[31],黄瓜Cucumis sativus中有11个[3],陆地棉Gossypium hirsutum中含有38个[32]。小麦(17 Gb)[33]、玉米(2 300 Mb)[34]和毛竹(2 021 Mb)[35]基因组大于杜仲(1.2 Gb)[36],拟南芥(164 Mb)[37]、水稻(441 Mb) [38]和毛果杨(392.3 Mb)[39]基因组小于杜仲。杜仲WOX数量低于拟南芥、毛果杨、水稻、玉米、小麦和毛竹,表明WOXs基因的丰富程度与基因组大小无关,这可能与基因重复有关。
杜仲基因组中共鉴定出8个EuWOXs,分布在8条染色体上。EuWOXs均为核定位蛋白,在现代进化支(WUS)、中间进化支和远古进化支分别含有5、1和2个成员,其系统发育模式与拟南芥、水稻、陆地棉等类似[29,31-32]。EuWOXs基因启动子中含有多种生长发育、激素响应、非生物胁迫以及光周期响应元件。在水稻中,WUS的OsWOX5和中间进化支的OsWOX11、OsWOX12A和OsWOX12B基因表达受生长素、细胞分裂素和赤霉素调节,超表达OsWOX11可提高水稻抗旱性[16-17]。细胞分裂素强烈促进苹果Malus pumila WOX1和WOX3基因表达,生长素诱导黄瓜CsWOX1b和CsWOX3基因表达[3]。在拟南芥中,生长素反应因子5 (AUXIN RESPONSE FACTOR,ARF5)上调AtWOX1和PRS (AtWOX3)基因的表达,ARF2、ARF3和ARF4抑制AtWOX1和PRS的表达[40]。OsWOX3A参与水稻器官发育、叶片横向轴伸长、颖花外稃形态发生以及分蘖和侧根发育[10],MtWOX1的同源基因STENOFOLIA是蒺藜苜蓿Medicago truncatula叶片生长和维管组织形成的必须基因[41],PttWOX4在杨树形成层中特异表达,PttWOX4a/b RNAi干扰后导致维管形成层宽度缩小,次生生长减弱[42]。推测EuWOXs可能在杜仲生长发育、激素和胁迫响应等生物学过程中发挥重要作用。
WOX家族基因参与叶片发育。属于中间支的AtWOX9/STIMPY过表达导致拟南芥叶缘波浪化[43],SlLAM1主要在番茄Solanum lycopersicum叶片、花和果实中表达,SlLAM1缺失导致叶片变窄,次生小叶数量减少[44],超表达黄瓜CsWOX9导致转基因拟南芥角果变短,莲座叶和分枝数目增加[3]。来源于WUS的AtWOX3是拟南芥侧托叶发育的必需基因,Atwox1和Atwox3缺失突变体导致叶片和花器官变窄,影响叶片横向扩张和花瓣融合[45-46];GhWOX9_At,GhWUSa_At和GhWUSb_Dt主要在棉花幼叶中高量表达[47]。远古进化分支中的OsWOX13在水稻叶、茎、根维管组织中表现为空间表达调控,在花和发育中的种子中是时间表达调控[48]。在杜仲中,EuWOX13-1在叶芽中表达量最高,随着叶片发育,转录水平逐渐降低,暗示EuWOX13-1主要在杜仲叶片发育的早期阶段发挥作用。EuWOX13-2在生长叶中表达量较高,在叶片发育过程中呈现先升高后降低的趋势。EuWOX13-1和EuWOX13-2是一对重复基因,其表达水平的差异可能与基因结构不同有关,也可能是EuWOX13-1和EuWOX13-2在重复后发生了功能分化。在甘蓝型油菜Brassica napus中,BnCWOX13a与BnCWOX13c互为同源基因,然而它们的表达趋势完全不同[49],在拟南芥中,AtWOX13在初生根、侧根、雌蕊和胚发育中动态表达,而AtWOX13的直系同源基因AtWOX14只在侧根形成的早期阶段和发育的花药中特异表达[50],由此推测EuWOX13-2可能只获得了EuWOX13-1基因的部分功能,具体功能还需要进一步研究。
Identification of WOX gene family and their expression in the leaf development of Eucommia ulmoides
-
摘要:
目的 Wuschel (WUS)相关的同源异型盒(Wuschel-related homeobox,WOX)转录因子家族在植物生长发育中发挥重要作用。本研究旨在探索WOX转录因子在杜仲Eucommia ulmoides中的分布及表达特征。 方法 以杜仲基因组数据库为基础,利用生物信息学方法对杜仲WOX家族进行全基因组鉴定;基于转录组数据分析EuWOXs在叶片发育及杜仲胶形成中的表达特征,通过实时荧光定量PCR (RT-qPCR)检测EuWOXs在‘紫叶’杜仲‘Ziye’叶片不同发育时期的表达模式。 结果 杜仲基因组中共鉴定出8条EuWOXs,分布于8条染色体;EuWOXs蛋白质长度为182~352个氨基酸,理论等电点为5.10~6.47,分子量为20.7~40.4 kDa;亚细胞定位预测EuWOXs均定位在细胞核中,均为亲水性蛋白。根据系统进化关系,杜仲WOX家族包括3个亚家族,分别含2、1和5个EuWOXs基因。EuWOXs均含有内含子,并包含多个基序,启动子中富含激素、胁迫和光周期响应元件。大部分EuWOXs在杜仲叶片中表达量较低,EuWOX13-1随叶片发育表达量逐渐降低,EuWOX13-2在生长叶中表达量最高。 结论 杜仲中有8个EuWOXs基因,EuWOX13-1和EuWOX13-2可能在杜仲叶片发育中发挥重要作用。图10表2参50 Abstract:Objective This study is aimed to explore the distribution and expression characteristics of WOX (Wuschel-related homeobox) transcription factors in Eucommia ulmoides as they play an important role in plant growth and development. Method First, bioinformatics method was employed to identify the genome-wide of EuWOX gene family on the basis of E. ulmoides genome database. Then an analysis was conducted of the expression levels of EuWOXs in E. ulmoides leaf development and Eu-rubber formation based on transcriptomic data. At last, the expression patterns of EuWOXs at different developmental stages of E. ulmoides ‘Ziye’ leaves were detected by real-time quantitative PCR (RT-qPCR). Result Eight EuWOXs were identified in E. ulmoides genome, distributing in eight chromosomes and they were composed of 182−352 amino acid residues with their isoelectric points being 5.10−6.47 and molecular weight between 20.7 to 40.4 kDa. According to the subcellular localization prediction, EuWOXs were all hydrophilic proteins and located in the nucleus. EuWOX gene family was divided into three subfamilies, containing two, one and five EuWOX genes respectively. All EuWOX genes had introns, and each EuWOX protein contains multiple motifs. EuWOXs promoters contained multiple hormones, stress and photoperiod response elemens. The expression level of most EuWOXs in E. ulmoides leaves was low and the expression of EuWOX13-1 gradually decreased with leaf development whereas the EuWOX13-2 expression was the highest in growing leaves. Conclusion Of the eight EuWOXs genes in E. ulmoides, EuWOX13-1 and EuWOX13-2 may play important roles in the leaf development of E. ulmoides. [Ch, 10 fig. 2 tab. 50 ref.] -
Key words:
- Eucommia ulmoides /
- WOX gene family /
- bioinformatics analysis /
- gene expression /
- leaf development
-
图 2 拟南芥和杜仲WOXs的蛋白质同源结构域序列分析
Figure 2 Sequence analysis of WOX proteins homeo domain in A. thaliana and E. ulmoides
图 3 杜仲、拟南芥、毛果杨、水稻和玉米WOXs蛋白系统发育树
Figure 3 WOX proteins phylogenetic trees of E. ulmoides, A. thaliana, P. trichocar, O. sativa and Z. mays
图 7 EuWOXs在杜仲叶片不同发育阶段的表达模式
Figure 7 Expression patterns of EuWOXs at different developmental stages in E. ulmoides leaves
图 8 杜仲EuWOXs基因在杜仲叶片不同发育时期的表达模式
Figure 8 Expression pattern of EuWOX genes in E. ulmoides leaves at different developmental stages
图 9 EuWOXs在杜仲胶形成中的表达模式
Figure 9 Expression patterns of EuWOX genes in the form of Eu-rubber
图 10 EuWOXs蛋白互作网络预测
Figure 10 Prediction of interaction network between EuWOX proteins
表 1 引物序列
Table 1. Primer sequences
基因名 上游引物(5′→3′) 下游引物(5′→3′) EuWOX1 ATGGTGGGTGACCAGCTTAG TTCTCTGGCCTTGTGGTTCT EuWOX2 ACCGTACCCCAACCTACTCC ACTTCCCGTTGGATGAAGTG EuWOX4-1 GGAACCCTACGCAAGAACAG GCGCTTCTGCTTTTGTCTCT EuWOX4-2 TAGAGCAGATCACGGCACAG CTAGGGTCGGATGTTGGAGA EuWOX5 GACGGAGCAAGTGAGAGTCC TCTCCCGTGCCTTATGATTC EuWOX11 ACTCGAGTTTTGTGGCCTGT AATTGGAGGCATCTGGATTG EuWOX13-1 GGTCTGAGGGCATGTGTTTT TTGGAGATATGGGTGGTGGT EuWOX13-2 GGGTTGTTCGTCAAGGTCAT GTTGGAATCCACCGTTGTCT UBCE2 AGTGGGTGGTGCTGTAGTCC AACTCCCGTTTCGTTTGTTG  下载: 导出CSV
下载: 导出CSV
表 2 EuWOXs蛋白质序列特征及亚细胞定位
Table 2. Sequence characteristics and subcellular location of E. ulmoides WOX proteins
基因号 基因名 拟南芥
同源基因染色体 位置 CDS长度/
bp氨基酸数/
个分子量/
kDa等电点 亚细胞
定位EUC13591-RA EuWOX1 AT3G18010.1 Super-Scaffold_235 3540694-3544292 1059 352 40.36 5.78 细胞核 EUC12552-RA EuWOX2 AT5G59340.1 Scaffold912_obj 156744-159059 810 269 30.16 8.11 细胞核 EUC15721-RA EuWOX4-1 AT1G46480.1 Super-Scaffold_242 604979-606289 618 205 23.48 9.04 细胞核 EUC21176-RA EuWOX4-2 AT1G46480.1 Scaffold272_obj 37477-39280 642 213 24.31 8.82 细胞核 EUC18832-RA EuWOX5 AT3G11260.1 Super-Scaffold_117 336319-340482 549 182 20.70 6.92 细胞核 EUC00362-RA EuWOX11 AT3G03660.1 Super-Scaffold_154 68808-70507 765 254 27.67 5.62 细胞核 EUC00756-RA EuWOX13-1 AT4G35550.1 Super-Scaffold_233 319468-325733 810 269 30.42 6.22 细胞核 EUC02503-RA EuWOX13-2 AT4G35550.1 Super-Scaffold_71 6332599-6364092 576 191 22.11 6.54 细胞核
下载: 导出CSV
-
[1] ZHANG Xin, ZONG Jie, LIU Jianhua, et al. Genome-wide analysis of WOX gene family in rice, sorghum, maize, Arabidopsis and poplar [J]. Journal of Integrative Plant Biology, 2010, 52(11): 1016 − 1026. [2] HAO Qingnan, ZHANG Ling, YANG Yanyan, et al. Genome-wide analysis of the WOX gene family and function exploration of GmWOX18 in soybean [J/OL]. Plants, 2019, 8(7): 215[2021-11-01]. doi: 10.3390/plants8070215. [3] GU Ran, SONG Xiaofei, LIU Xiaofeng, et al. Genome-wide analysis of CsWOX transcription factor gene family in cucumber (Cucumis sativus L. ) [J/OL]. Scientific Reports, 2020, 10: 6216[2021-11-01]. doi: 10.1038/s41598-020-63197-z. [4] HAECKER A, GROß-HARDT R, GEIGES B, et al. Expression dynamics of WOX genes mark cell fate decisions during early embryonic patterning in Arabidopsis thaliana [J]. Development, 2004, 131(3): 657 − 668. [5] BREUNINGER H, RIKIRSCH E, HERMANN M, et al. Differential expression of WOX genes mediates apical-basal axis formation in the Arabidopsis embryo [J]. Developmental Cell, 2008, 14(6): 867 − 876. [6] DOLZBLASZ A, NARDMANN J, CLERICI E, et al. Stem cell regulation by Arabidopsis WOX genes [J]. Molecular Plant, 2016, 9(7): 1028 − 1039. [7] COSTANZO E, TREHIN C, VANDENBUSSCHE M. The role of WOX genes in flower development [J]. Annals of Botany, 2014, 114(7): 1545 − 1553. [8] LAUX T, MAYER K F, BERGER J, et al. The WUSCHEL gene is required for shoot and floral meristem integrity in Arabidopsis [J]. Development, 1996, 122(1): 87 − 96. [9] BOUCHABKÉ-COUSSA O, OBELLIANNE M, LINDERME D, et al. Wuschel overexpression promotes somatic embryo genesis and induces organo genesis in cotton (Gossypium hirsutum L. ) tissues cultured in vitro [J]. Plant Cell Reports, 2013, 32(5): 675 − 686. [10] ZHAO Yu, HU Yongfeng, DAI Mingqiu, et al. The WUSCHEL-related homeobox gene WOX11 is required to activate shoot-borne crown root development in rice [J]. The Plant Cell, 2009, 21(3): 736 − 748. [11] CHO S H, YOO S C, ZHANG Haitao, et al. The rice narrow leaf2 and narrow leaf3 loci encode WUSCHEL-related homeobox 3A (OsWOX3A) and function in leaf, spikelet, tiller and lateral root development [J]. New Phytologist, 2013, 198(4): 1071 − 1084. [12] OHMORI Y, TANAKA W, KOJIMA M, et al. WUSCHEL-RELATED HOMEOBOX4 is involved in meristem maintenance and is negatively regulated by the CLE gene FCP1 in rice [J]. The Plant Cell, 2013, 25: 229 − 241. [13] LIU Bobin, WANG Lin, ZHANG Jin, et al. WUSCHEL-related Homeobox genes in Populus tomentosa: diversified expression patterns and a functional similarity in adventitious root formation [J/OL]. BMC Genomics, 2014, 15: 296[2021-11-01]. doi: 10.1186/1471-2164-15-296. [14] XU Meng, XIE Wenfan, HUANG Minren. Two WUSCHEL-related HOMEOBOX genes, PeWOX11a and PeWOX11b, are involved in adventitious root formation of poplar [J]. Physiologia Plantarum, 2015, 155(4): 446 − 456. [15] LI Zheng, LIU Dan, XIA Yu, et al. Identification of the WUSCHEL-related Homeobox (WOX) gene family, and interaction and functional analysis of TaWOX9 and TaWUS in wheat [J/OL]. International Journal of Molecular Sciences, 2020, 21(5): 1581[2021-11-01]. doi: 10.3390/ijms21051581. [16] CHENG Saifeng, HUANG Yulan, ZHU Ning, et al. The rice WUSCHEL-related homeobox genes are involved in reproductive organ development, hormone signaling and abiotic stress response [J]. Gene, 2014, 549(2): 266 − 274. [17] CHENG Saifeng, ZHOU Daoxiu, ZHAO Yu. WUSCHEL-related homeobox gene WOX11 increases rice drought resistance by controlling root hair formation and root system development [J/OL]. Plant Signaling & Behavior, 2016, 11(2): e1130198[2021-11-01]. doi: 10.1080/15592324.2015.1130198. [18] WANG Liuqiang, LI Zhen, WEN Shuangshuang, et al. WUSCHEL-related homeobox gene PagWOX11/12a responds to drought stress by enhancing root elongation and biomass growth in poplar [J]. Journal of Experimental Botany, 2020, 71(4): 1503 − 1513. [19] WANG Liuqiang, WEN Shuangshuang, WANG Rui, et al. PagWOX11/12a activates PagCYP736A12 gene that facilitates salt tolerance in poplar [J]. Plant Biotechnology Journal, 2021, 19(11): 2249 − 2260. [20] 杜红岩. 中国杜仲图志[M]. 北京: 中国林业出版社, 2014. DU Hongyan. Chinese Eucommia Pictorial [M]. Beijing: China Forestry Publishing House, 2014. [21] LI Zhenyu, GU Juan, YAN Jin, et al. Hypertensive cardiac remodeling effects of lignan extracts from Eucommia ulmoides Oliv. bark: a famous traditional Chinese medicine [J]. The American Journal of Chinese Medicine, 2013, 41(4): 801 − 815. [22] WU Dong, YU Danmeng, ZHANG Yujia, et al. Metabolite profiles, bioactivity, and HPLC fingerprint of different varieties of Eucommia ulmoides Oliv. : Towards the utilization of medicinal andcommercial chinese endemic tree [J/OL]. Molecules, 2018, 23(8): 1898[2021-11-01]. doi: 10.3389/molecules.23081898. [23] 刘俊, 陈玉龙, 刘燕, 等. 杜仲TIFY转录因子鉴定与表达分析[J]. 中国实验方剂学杂志, 2021, 27(19): 165 − 174. LIU Jun, CHEN Yulong, LIU Yan, et al. Identification and expression analysis of TIFY transcription factor in Eucommia ulmoides [J]. Chinese Journal of Experemental Traditional Medical Formulae, 2021, 27(19): 165 − 174. [24] TAMURA K, STECHER G, PETERSON D, et al. MEGA6: molecular evolutionary genetics analysis version 6.0 [J]. Molecular Biology &Evolution, 2013, 30(12): 2725 − 2729. [25] LI Long, LIU Minhao, SHI Kan, et al. Dynamic changes in metabolite accumulation and the transcriptome during leaf growth and development in Eucommia ulmoides [J/OL]. International Journal of Molecular Sciences, 2019, 20(16): 4030[2021-11-01]. doi: 10.3390/ijms20164030. [26] YE Jing, HAN Wenjing, FAN Ruisheng, et al. Integration of transcriptomes, small RNAs, and degradome sequencing to identify putative miRNAs and their targets related to eu-rubber biosynthesis in Eucommia ulmoides [J/OL]. Genes, 2019, 10(8): 623[2021-11-01]. doi: 10.3390/genes10080623. [27] CHEN Chengjie, CHEN Hao, ZHANG Yi, et al. TBtools: an integrative toolkit developed for interactive analyses of big biological data [J]. Molecular Plant, 2020, 13(8): 1194 − 1202. [28] LIU Jun, CHENG Zhanchao, XIE Lihua, et al. Multifaceted role of PheDof12-1 in the regulation of flowering time and abiotic stress responses in moso bamboo (Phyllostachys edulis) [J/OL]. International Journal of Molecular Sciences, 2019, 20(2): 424[2021-11-01]. doi: 10.3390/ijms20020424. [29] LI Xiangyu, LI Juan, CAI Miaomiao, et al. Identification and evolution of the WUSCHEL related homeobox protein family in Bambusoideae [J/OL]. Biomolecules, 2020, 10(5): 739[2021-11-01]. doi: 10.3390/biom10050739. [30] 武强强, 张凤洁, 董浩欢, 等. 小麦WOX转录因子基因的全基因组鉴定与分析[J]. 激光生物学报, 2021, 30(1): 67 − 74. WU Qiangqiang, ZHANG Fengjie, DONG Haohuan, et al. Genome-wide identification and analyses of WOX transcription factor genes in wheat [J]. Acta Laser Biology Sinica, 2021, 30(1): 67 − 74. [31] WANG Pengjie, GUO Yongchun, CHEN Xuejin, et al. Genome-wide identification of WOX genes and their expression patterns under different hormone and abiotic stress treatments in tea plant (Camellia sinensis) [J]. Trees, 2019, 33(4): 1129 − 1142. [32] YANG Zhaoen, GONG Qian, QIN Wenqiang, et al. Genome-wide analysis of WOX genes in upland cotton and their expression pattern under different stresses [J/OL]. BMC Plant Biology, 2017, 17(1): 113[2021-11-01]. doi: 10.1186/s12870-017-2015-8. [33] International Wheat Genome Sequencing Consortium. A chromosomebased draft sequence of the hexaploid bread wheat (Triticum aestivum) genome [J/OL]. Science, 2014, 345(6194): 1251788[2021-11-01]. doi: 10.1126/science.1251788. [34] WANG Guifeng, ZHONG Mingyu, WANG Jiajia, et al. Genome-wide identification, splicing, and expression analysis of the myosin gene family in maize (Zea mays) [J]. Journal of Experimental Botany, 2014, 65(4): 923 − 938. [35] PENG Zhenhua, LU Ying, LI Lubin, et al. The draft genome of the fast-growing non-timber forest species moso bamboo (Phyllostachys heterocycla) [J]. Nature Genetics, 2013, 45(4): 456 − 461. [36] WUYUN Tana, WANG Lin, LIU Huimin, et al. The hardy rubber tree genome provides insights into the evolution of polyisoprene biosynthesis [J]. Molecular Plant, 2018, 11(3): 429 − 442. [37] The International Brachypodium Initiative. Genome sequencing and analysis of the model grass Brachypodium distachyon [J]. Nature, 2009, 463: 763 − 768. [38] BURR B. Mapping and sequencing the rice genome [J]. The Plant Cell, 2002, 14(3): 521 − 523. [39] HOFMEISTER B T, DENKENA J, COLOMÉ-TATCHÉ M, et al. A genome assembly and the somatic genetic and epigenetic mutation rate in a wild long-lived perennial Populus trichocarpa [J/OL]. Genome Biology, 2020, 21: 259[2021-11-01]. doi: 10.1186/s13059-020-02162-5. [40] GUAN Chunmei, WU Binbin, YU Ting, et al. Spatial auxin signaling controls leaf flattening in Arabidopsis [J]. Current Biology, 2017, 27(19): 2940 − 2950. [41] TADEGE M, LIN Hao, BEDAIR M, et al. STENOFOLIA regulates blade outgrowth and leaf vascular patterning in Medicago truncatula and Nicotiana sylvestris [J]. The Plant Cell, 2011, 23(6): 2125 − 2142. [42] KUCUKOGLU M, NILSSON J, ZHENG Bo, et al. WUSCHEL-RELATED HOMEOBOX4 (WOX4)-like genes regulate cambial cell division activity and secondary growth in Populus trees [J]. New Phytologist, 2017, 215(2): 642 − 657. [43] BLEIN T, PAUTOT V, LAUFS P. Combinations of mutations sufficient to alter Arabidopsis leaf dissection [J]. Plants, 2013, 2: 230 − 247. [44] WANG Chaoqun, ZHAO Baolin, HE Liangliang, et al. The WOX family transcriptional regulator SlLAM1 controls compound leaf and floral organ development in Solanum lycopersicum [J]. Journal of Experimental Botany, 2021, 72(5): 1822 − 1835. [45] VANDENBUSSCHE M, HORSTMAN A, ZETHOF J, et al. Differential recruitment of WOX transcription factors for lateral development and organ fusion in Petunia and Arabidopsis [J]. The Plant Cell, 2009, 21(8): 2269 − 2283. [46] NAKATA M, MATSUMOTO N, TSUGEKI R, et al. Roles of the middle domain-specifific WUSCHEL-RELATED HOMEOBOX genes in early development of leaves in Arabidopsis [J]. The Plant Cell, 2012, 24(2): 519 − 535. [47] HE Peng, ZHANG Yuzhou, LIU Hao, et al. Comprehensive analysis of WOX genes uncovers that WOX13 is involved in phytohormone-mediated fiber development in cotton [J/OL]. BMC Plant Biology, 2019, 19: 312[2021-11-01]. doi: 10.1186/s12870-019-1892-x. [48] MINH-THU P T, KIM J S, CHAE S, et al. A WUSCHEL homeobox transcription factor, OsWOX13, enhances drought tolerance and triggers early flowering in rice [J]. Moleculer Cells, 2018, 41(8): 781 − 798. [49] LI Mengdi, WANG Ruihua, LIU Zhengyi, et al. Genome-wide identification and analysis of the WUSCHEL-related homeobox (WOX) gene family in allotetraploid Brassica napus reveals changes in WOX genes during polyploidization [J/OL]. BMC Genomics, 2019, 20: 317[2021-11-01]. doi: 10.1186/s12864-019-5684-3. [50] DEVEAUX Y, TOFFANO-NIOCHE C, CLAISSE G, et al. Genes of the most conserved WOX clade in plants affect root and flower development in Arabidopsis [J/OL]. BMC Evolutionary Biology, 2008, 8: 291[2021-11-01]. doi: 10.1186/1471-2148-8-291. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20210725
 点击查看大图
点击查看大图

计量
- 文章访问数: 1784
- HTML全文浏览量: 559
- PDF下载量: 158
- 被引次数: 0
