




 下载:
下载:

-
桂花Osmanthus fragrans为木犀科Oleaceae园林观赏植物,是中国十大传统名花之一,桂花树形优美、叶色青翠、香味馥郁,在园林中应用广泛[1]。目前,桂花在食品、化妆品和药品中的应用已经逐渐成熟[2],新兴的桂花香水、乳液、香熏和精油等高档化妆品也逐渐打开市场[3−5]。然而,桂花花期较短,最佳观赏期和采收期仅2~3 d,极大地限制了其观赏价值与经济价值[6−7]。
已有研究证明:OfABFs基因可能参与调控桂花花瓣衰老[8];蒋琦妮等[9]研究发现:桂花OfAP1基因在桂花成花转变、花芽分化和发育中有重要作用。向其柏等[10]研究发现:许多经济采收价值较高的桂花品种对乙烯敏感,非授粉诱导的内源乙烯跃变是其衰老的重要调控因子。朱诚等[11]研究表明:盛花末期乙烯释放量的迅速增加及膜脂过氧化程度加剧是导致‘薄叶金桂’O. fragrans‘Baoye Jingui’衰老的主要生理原因。ZHOU等[12]发现:外源乙烯利处理明显加速‘厚瓣金桂’O. fragrans‘Houban Jingui’和‘柳叶金桂’O. fragrans‘Liuye Jingui’的衰老,而乙烯抑制剂硫代硫酸银则延长了其观赏寿命。ZOU等[6]发现‘柳叶金桂’内源乙烯跃变峰的出现与花瓣褐化、脱落等衰老特征同时发生,外源乙烯处理不仅明显加速了切花花瓣的萎蔫和脱落,还导致花瓣细胞中央大液泡破裂、各细胞器扭曲挤缩变形,加剧了 DNA 断裂,降低了抗氧化酶活性,增加了超氧自由基的激发和膜脂过氧化程度。由此可见:桂花是乙烯敏感型花卉,乙烯参与了其花瓣衰老过程中花瓣脱落和萎蔫、细胞结构变化、氧化还原系统及核酸降解等多个过程的调节,是桂花衰老的重要调控因子。然而,尚不清楚该过程中内源乙烯合成途径。
氨基环丙烷羧酸氧化酶(1-aminocyclopropane-1-carbox-ylate oxidase, ACO)作为乙烯生物合成途径中的最后一个关键酶,直接催化乙烯的合成,被认为是高等植物中乙烯应答的主要标志[13]。ACO早期被ADAMS等[14]发现,并命名为乙烯形成酶,后来发现需要抗坏血酸和氧作为辅助底物,因此称为ACC氧化酶。目前,ACO基因家族在多个物种中都有研究,SORNCHAI 等[15]通过导入石斛Dendrobium CpACO 基因延长了其花期;有研究表明:杨树Populus ACO基因参与调节林木茎的生长发育[16]。植物ACO基因的表达受到生长素、干旱以及盐胁迫的抑制[17];番茄Lycopersicon esculentum、花椰菜Brassica oleracea var. botrytis的ACO基因表达受脱落酸、外部机械损伤和低温的诱导[18]。在拟南芥Arabidopsis thaliana中[19],ACO1受多种信号调控,影响植株的乙烯产量。然而,目前桂花中还未见相关基因的报道。本研究以‘柳叶金桂’为试材,利用生物信息学方法和工具对桂花OfACOs家族进行鉴定、进化分析、基因结构分析、表达模式分析等,为进一步探索桂花花瓣衰老机制以及提高桂花园林赏花价值与经济价值提供指导。
-
‘柳叶金桂’花瓣采自华中农业大学校园内。分别采集不同开花阶段的桂花花瓣,称量后密封,保存在液氮中,立即运送回实验室,置于−80 ℃超低温冰箱备用。参考CHEN等[20]对桂花开花级数的划分体系,将‘柳叶金桂’开花级数划分为:①铃梗期(花苞期,S1),花朵呈紧闭的花苞状,未展开;②初花期(S2),花朵微张,呈半开放状态;③盛花初期(S3),花瓣进一步展开,夹角为45°~90°;④盛花期(S4),花瓣完全展开,花药膨大浅黄色;⑤盛花后期(S5),花瓣完全展开,花药萎缩变为深褐色;⑥脱落期(S6),花瓣失水,部分从树体上自然脱落。
-
ACO基因家族成员大多含有DIOX-N (PF14226)和2OG-FeII-Oxy (PF03171)保守结构域[21]。对‘柳叶金桂’基因进行蛋白质家族数据库(pfam)保守结构域注释,以注释到PF03171和PF14226保守结构域的基因作为OfACOs基因家族候选基因,再通过蛋白结构域搜索(NCBI Conserved Domain Search Service) (
https://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi )[22]进一步验证候选基因的PF03171和PF14226结构域。 -
登录拟南芥官网 (www.arabidopsis.org)下载拟南芥AtACO基因序列信息。利用MUSCLE [23]比对各基因多序列,采用邻接算法进行系统进化树聚类分析。
-
对‘柳叶金桂’基因进行pfam保守结构域注释,获取OfACOs成员序列信息及其在各染色体上的定位信息,然后使用Mapchart (2.32)[24]绘制染色体上的ACO基因。
-
利用 MEME软件( Suite version 5.0.2,
http://meme-suite.org/ ) 对 OfACOs保守基序进行预测分析,设置Motif个数为15个。根据基因注释的gff文件,利用TBtools (v1.098775)[25]对基因结构、Motif进行可视化。 -
为分析OfACOs基因家族成员的重复关系,通过使用blast (2.11.0+)和MCScanX[26]对家族成员进行共线性分析,使用默认参数输出数据,结果使用circos (0.69)圈图展示。此外,下载了拟南芥ACO基因家族成员的基因注释文件,通过使用blast (2.11.0+)和MCScanX 对家族成员进行共线性分析,使用默认参数输出数据,结果使用circos (0.69)圈图展示。
-
从美国国家生物技术信息中心(NCBI)数据库下载桂花在根、茎、叶及不同开花阶段等组织部位的转录组测序数据(PRJNA679852),以基因表达量量化指标 (FPKM)计算基因表达水平值,用 TBtools [25]对数据可视化。
-
取S1~S6不同开花阶段的桂花花瓣,按HiPure Universal RNA Kit提取试剂盒说明提取桂花花瓣RNA。用StarScript II RT Kit反转录试剂盒将RNA逆转录为cDNA,稀释10倍备用。利用染料法(SYBR)预混2×RealStar Fast在ABI 7500 系统(Thermo Fisher Scientific, Inc.)进行实时荧光定量PCR分析(RT-qPCR)。RT-qPCR体系为10.0 μL 2×RealStar Fast SYBR qPCR Mix,上下游引物各0.8 μL,2.0 μL cDNA,6.4 μL ddH2O。RT-qPCR的反应程序为:94 ℃ 30 s;40个循环:94 ℃ 10 s,60 ℃ 30 s,72 ℃ 10 s。以 ACTIN 作为内参,采用 2−ΔΔCt法计算基因的相对表达量,每个组织每个基因设3个重复。
-
每个试验的每个处理进行3次生物学重复。数据为平均值 ± 标准误。所有数据用SAS v.8.0进行方差分析和多重比较分析,在0.05水平上进行邓肯氏多重比较。
-
通过pfam数据库PF03171和PF14226保守结构域注释分析,从桂花基因组中共鉴定到122个OfACOs基因家族成员(图1)。这122个OfACOs家族成员主要分布于除19号染色体以外的其他22条染色体上,其中,1号14条、2号4条、3号13条、4号4条、5号2条、6号6条、7号和8号各1条、9号7条、10号8条、11号2条、12号5条、13号和14号各3条、15号4条、16号7条、17号3条、18号6条、20号8条、21号和23号各1条、22号11条。其中1号染色体上基因最多,7号、8号、21号和23号染色体上分布最少。
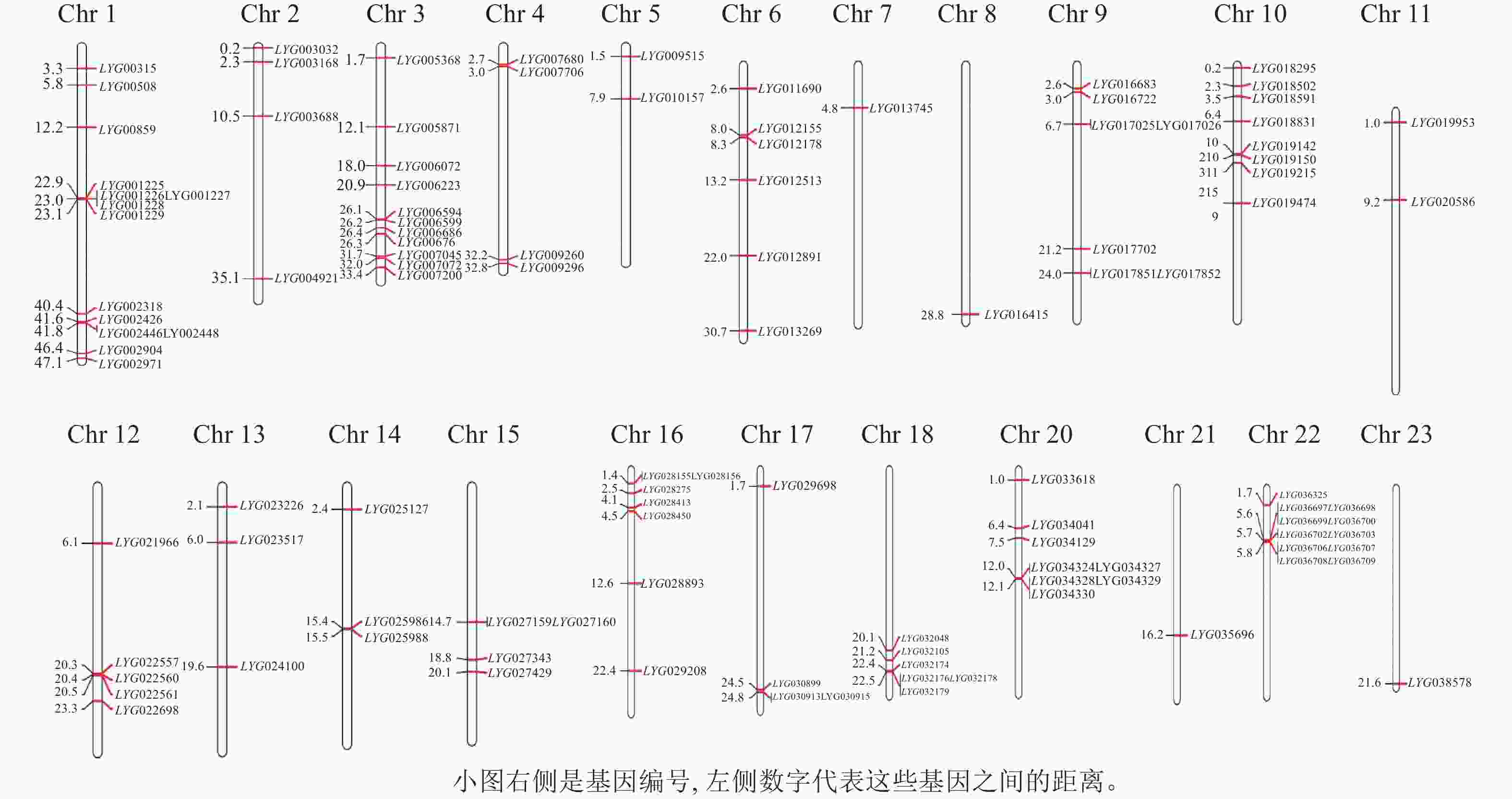
图 1 桂花OfACOs基因家族的染色体定位
Figure 1. Chromosomal localization of OfACOs gene family in O. fragrans
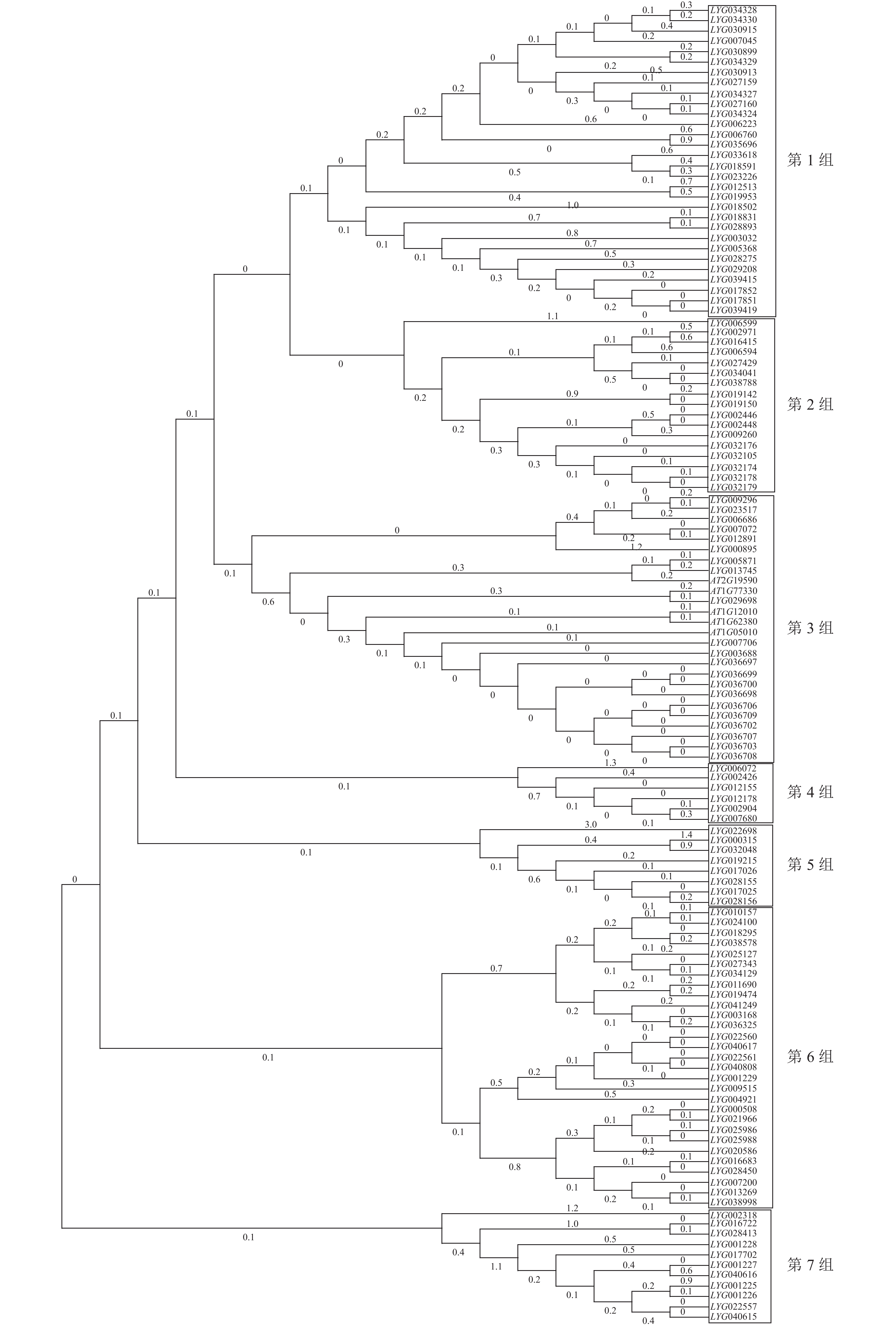
用桂花OfACOs和拟南芥AtACOs构建进化树,结果如图2所示。通过系统聚类,大致将OfACOs分为7组,其中拟南芥基因AT1G05010、AT1G62380、AT1G77330、AT2G19590和AT1G12010与桂花LYG005871、LYG00776等21个OfACOs共存于第3组中。其他组不存在拟南芥AtACOs基因。通过拟南芥官网查找发现这5个基因都具有促进乙烯生成的功能。
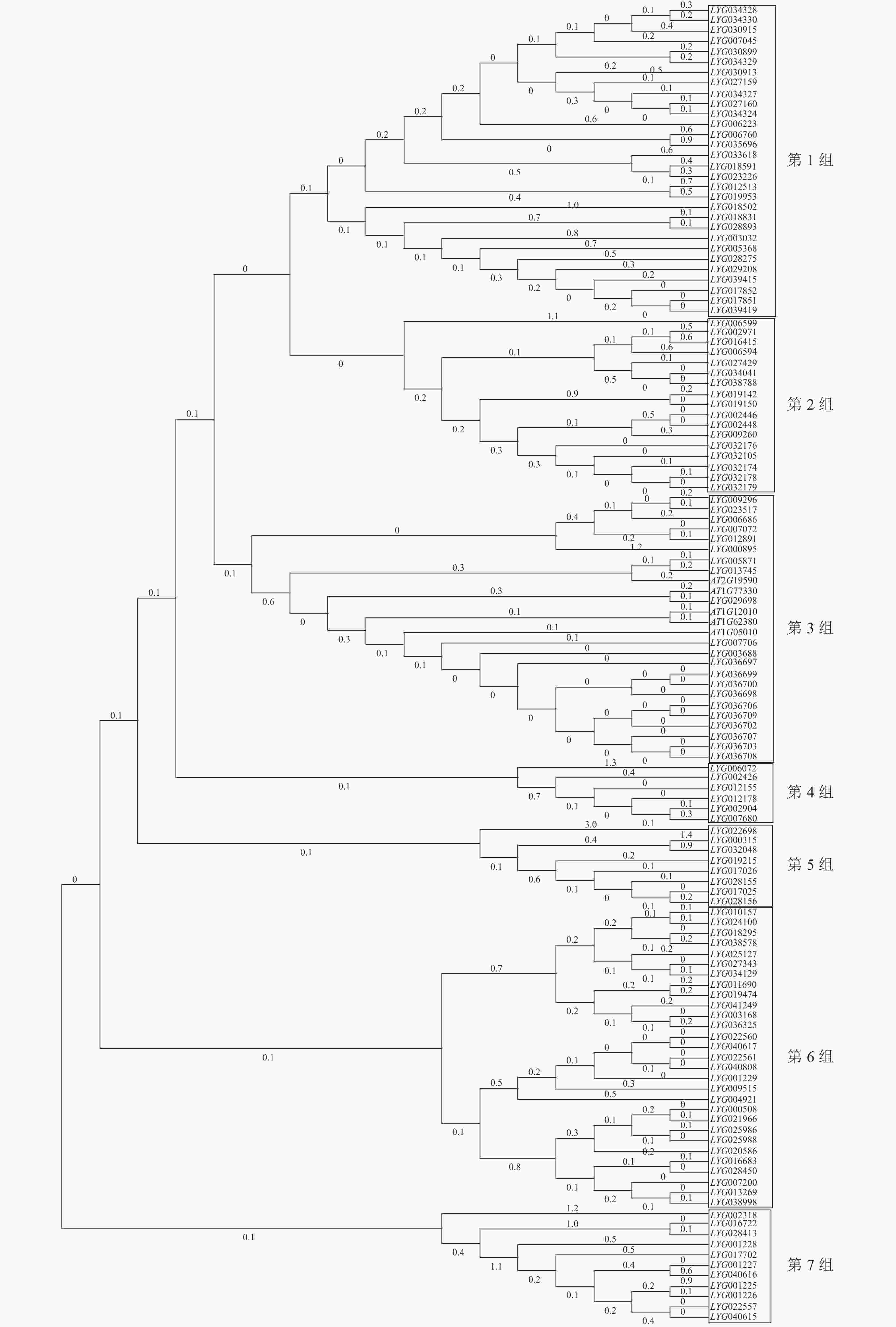
图 2 桂花与拟南芥ACOs基因进化分析
Figure 2. Phylogenetic analysis of ACOs in O. fragrans and A. thaliana
-
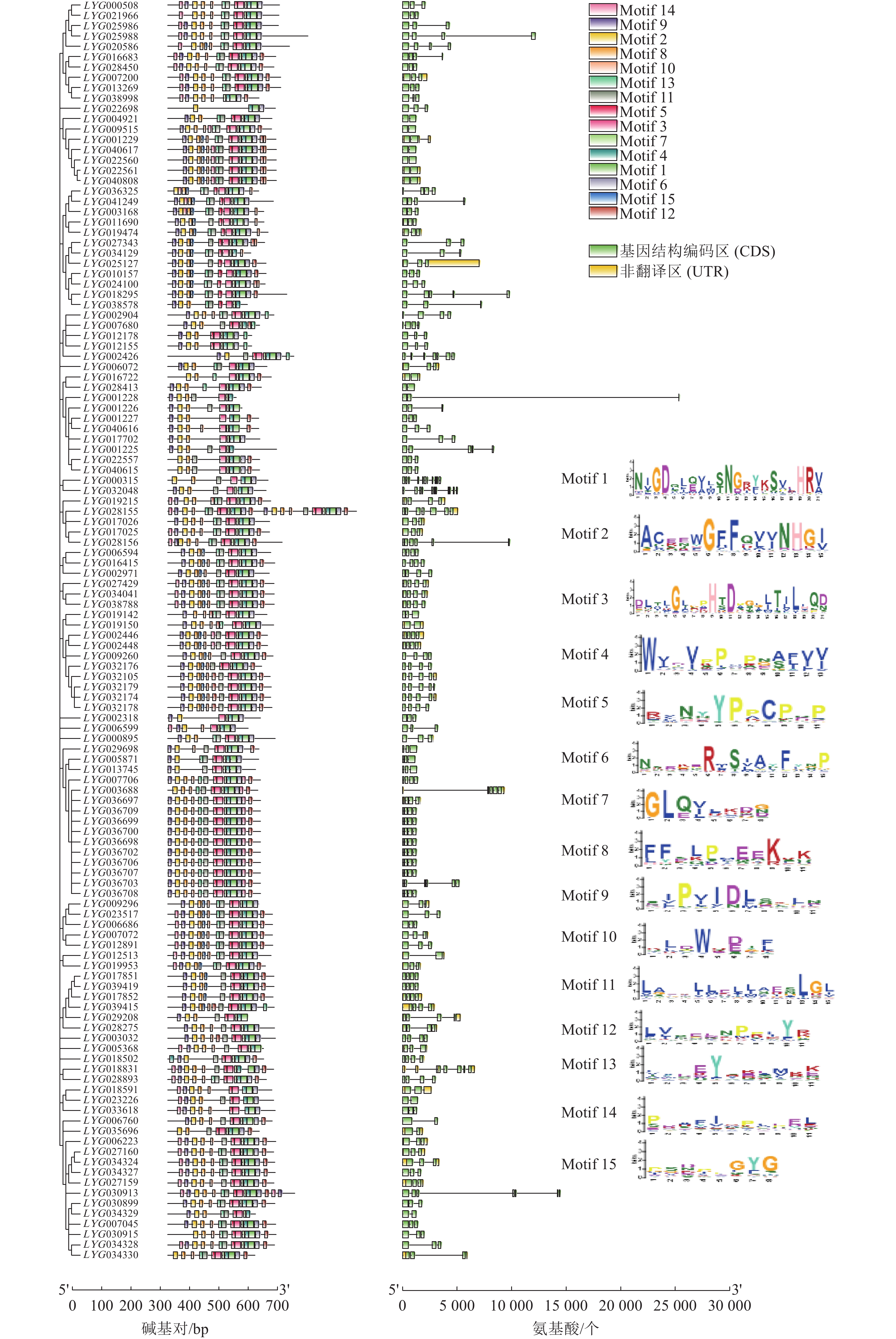
进一步对桂花122个OfACOs进行基因结构和保守基序分析(图3)。基因结构编码区(CDS)、非翻译区(UTR)分析结果表明:OfACOs基因的CDS数量为2~11个,其中52个成员含有3个CDS区(42.6%),41个成员含有4个CDS区(33.6%)。其中LYG025988、LYG018295、LYG001228、LYG001225、LYG028156和LYG030913的CDS区分布较散。LYG000315是CDS区最多的基因。
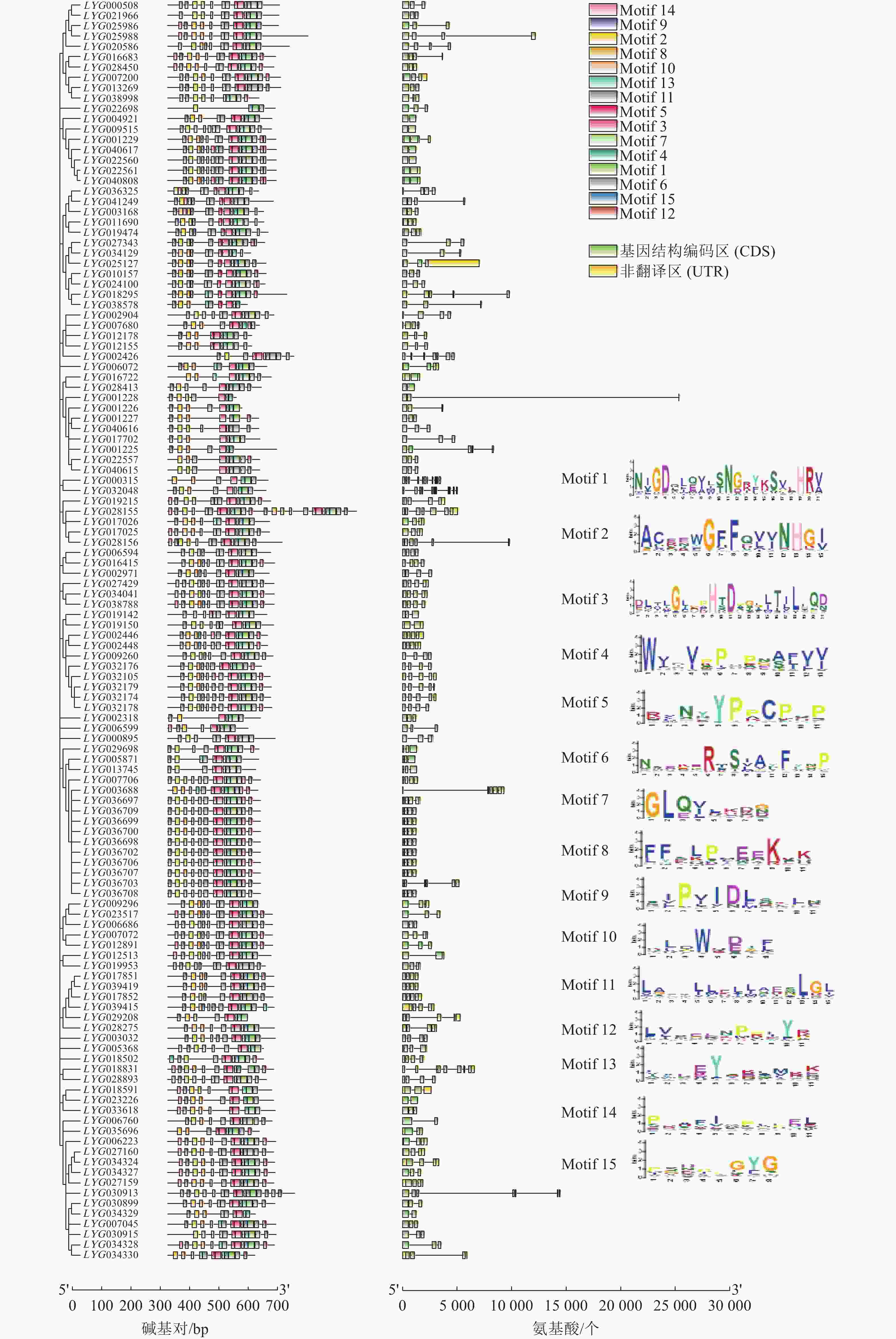
图 3 桂花OfACOs基因结构及蛋白质保守结构域
Figure 3. Gene structure and conserved motifs analysis of OfACOs in O. fragrans
对122个OfACOs的氨基酸序列进行保守基序分析表明:OfACOs的Motif为4~25个,其中,LYG022698为最少,只有4个Motif,而LYG028155有25个Motif;24个OfACOs成员含有14个Motif,且其中有11个的Motif构成完全相同,都位于第3组中;几乎所有的OfACOs成员中都含有Motif 1、Motif 2、Motif 3、Motif 4和Motif 7,表明在桂花OfACOs家族中这5个Motif的保守性更高。
-
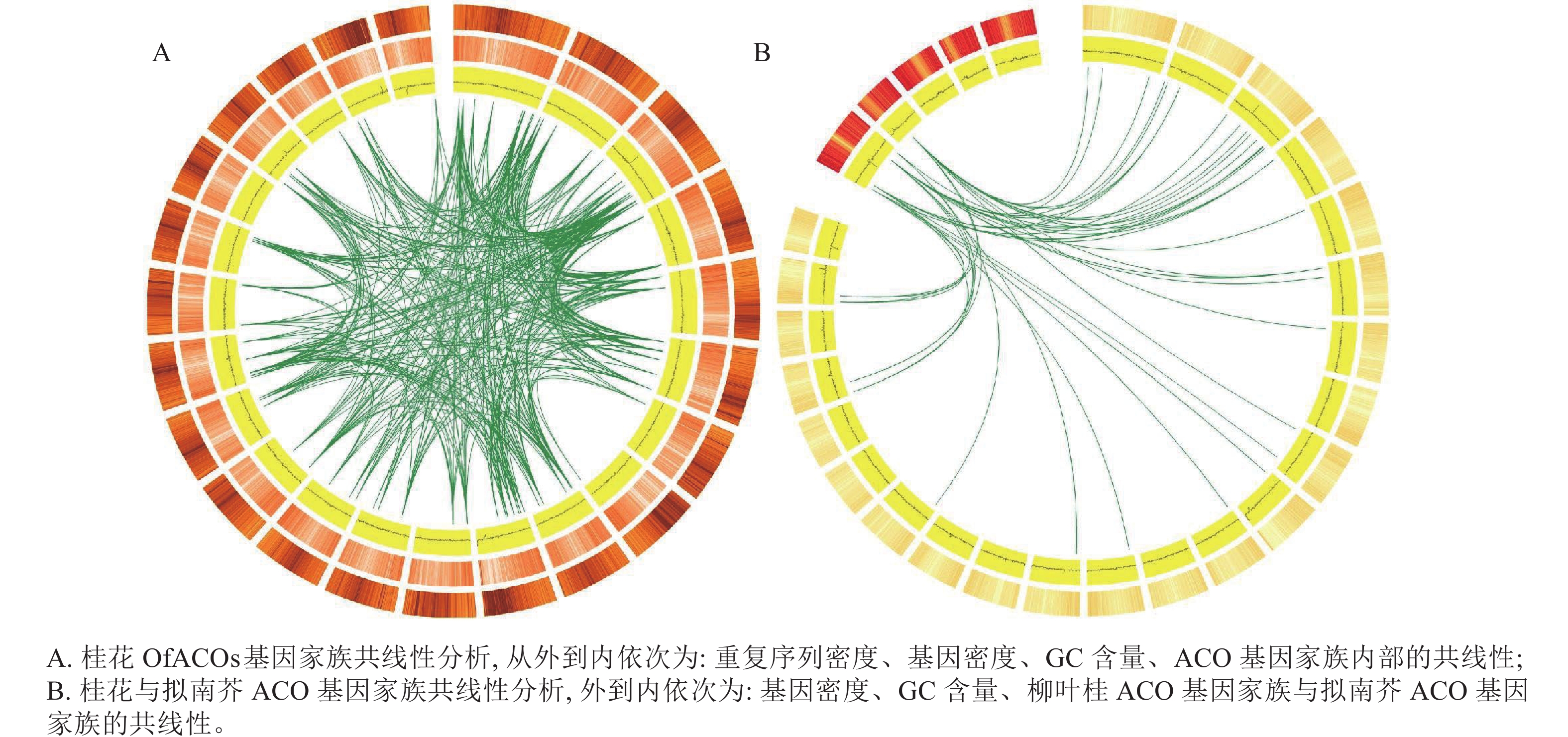
基因组内共线性分析结果(图4A)表明:染色体复制事件发生在桂花的22条染色体上,OfACOs共存在394对染色体共线性对,其中1号染色体上最多,有84对,说明该基因家族可能发生了扩张。
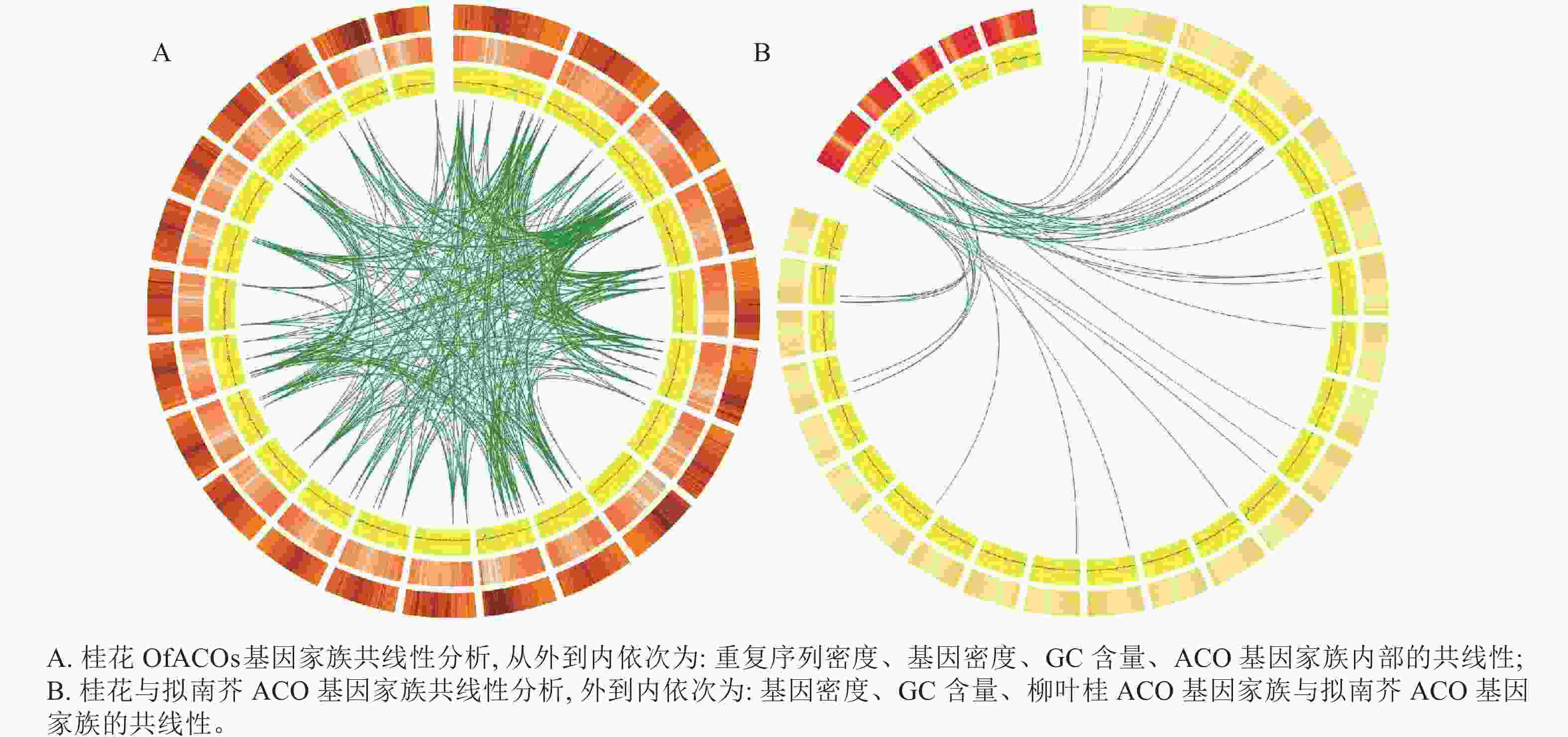
图 4 桂花及拟南芥ACO基因家族共线性分析
Figure 4. Collinearity analysis of ACO gene family in O. fragrans and A. thaliana
桂花与拟南芥ACO基因家族的共线性分析表明:拟南芥的2条染色体与桂花的15条染色体共发生了41对染色体复制对,其中拟南芥1号染色体存在29对染色体复制对(图4B)。综上所述,通过与拟南芥ACO基因共线性分析,有助于利用拟南芥的基因功能探索桂花OfACOs中相应基因的功能。
-
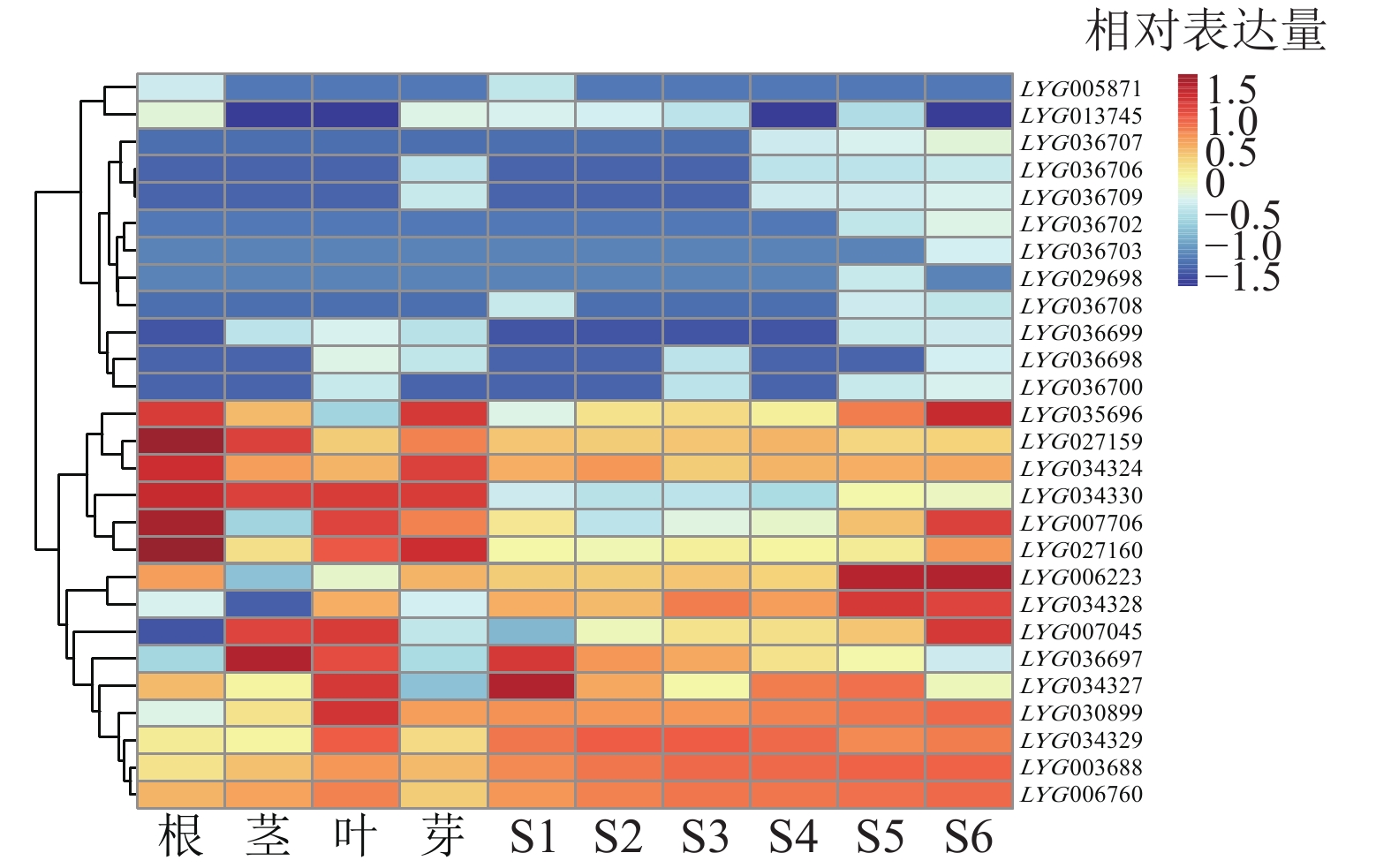
具有相同的DIOX-N (PF14226)和2OG-FeII-Oxy (PF03171)保守结构域并不一定能行使相同的蛋白功能。为更准确地筛选OfACOs 蛋白,通过swiss-prot同源序列对比[27],筛选出27个基因家族成员。对27个OfACOs成员在根、茎、叶、花芽,以及不同开花阶段(S1~S6)进行转录组分析。由图5可知:除去FPKM小于1的成员后,在根茎叶花中表达的OfACOs成员共计17个。其中,LYG013745、LYG034324、LYG027159、LYG027160主要在根中微量表达;LYG007706主要在根、叶和花中表达;LYG007045、LYG030899、LYG036697主要在茎、叶和花中表达;LYG034328在叶和花中表达;LYG003688、LYG006223、LYG006760、LYG034327、LYG034329在根、茎、叶、花中均有表达;LYG034330主要在根、茎、叶中表达;LYG035696主要在根和花中表达;LYG036707在开花后期微量表达。
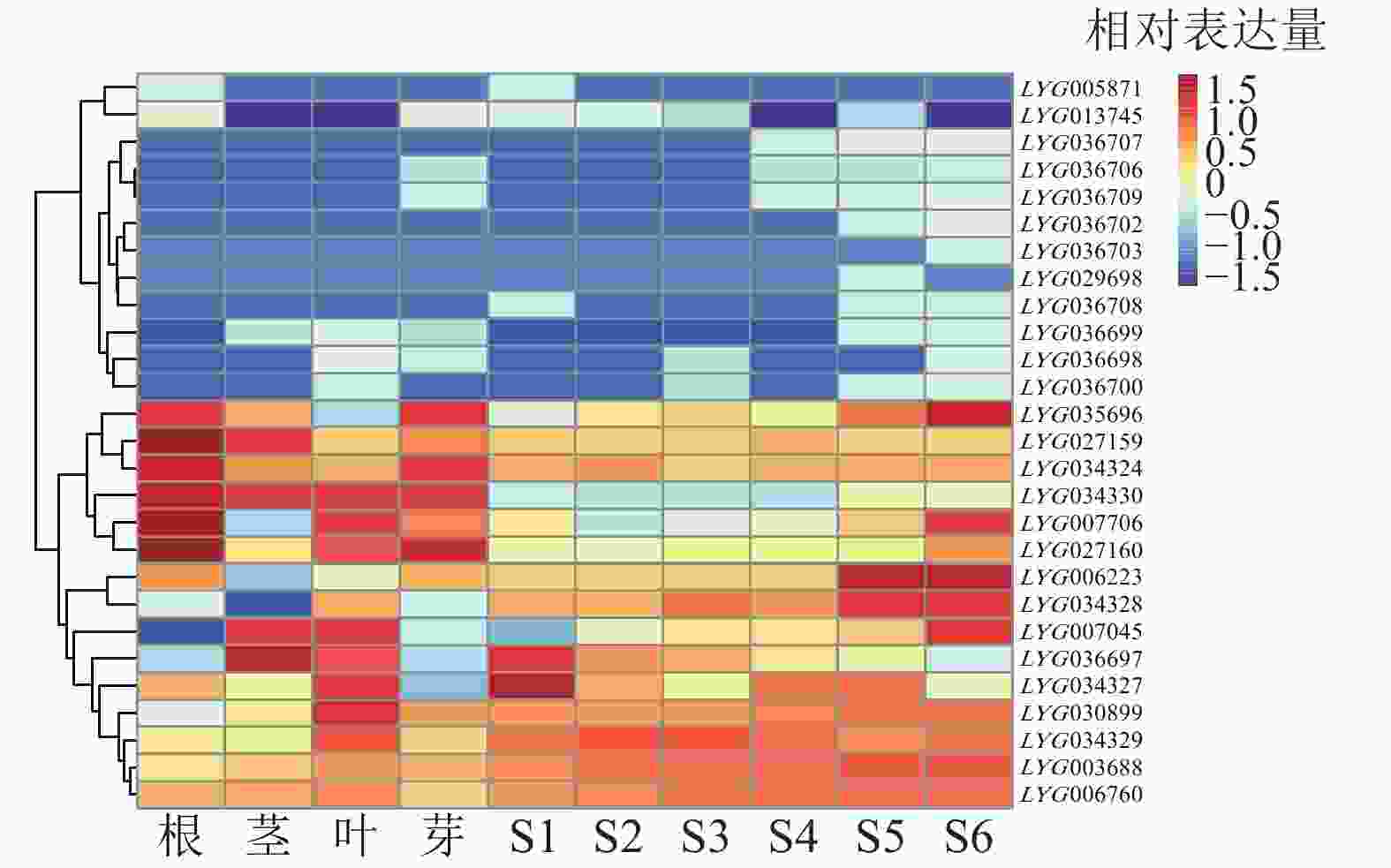
图 5 桂花OfACOs基因家族在桂花不同组织部位以及不同开花阶段中的表达分析
Figure 5. Expression profiles of OfACOs gene family in different tissues and different flowering stages of O. fragrans
-
综合OfACOs基因家族在不同组织部位以及不同开花时期的表达量,以在桂花花瓣中明显表达的成员作为参与乙烯途径花瓣衰老调控的研究对象,筛选出LYG003688、LYG006223、LYG034327、LYG034329、LYG034330、LYG035696、LYG007706、LYG007045、LYG030899、LYG036697、LYG034328、LYG036707等12个成员,其中LYG006223、LYG007706、LYG007045、LYG035696等4个基因在开花后期(S5或S6)极显著差异上调表达。
进一步对这4个OfACOs成员进行RT-qPCR验证分析(图6)。结果表明:这4个成员在开花后期(S5或S6)显著性差异上调表达(P<0.05),与转录组测序结果一致。

图 6 桂花花瓣候选OfACOs实时荧光定量PCR分析
Figure 6. RT-qPCR analysis of candidate OfACOs genes in O. fragrans
-
ACO基因家族是植物的转录因子家族之一,在植物乙烯生物合成中发挥着重要的调控作用[28]。本研究从‘柳叶金桂’的基因组中鉴定出122个ACO基因,基因家族总数量高于普通烟草Nicotiana tabacum (19个ACO基因)[29]、杞柳Salix integra (42个ACO基因)[21]、玉米Zea mays (8个ACO基因)[30]和甜瓜Cucumis melo (9个ACO基因)[17]中的数量,低于陆地棉Gossypium hirsutum (332个ACO基因)[31]。ACO基因在植物基因组中分布不均匀,如玉米[32]中62.0%的基因主要分布在其中2条染色体上,普通烟草[29]40.0%的基因主要分布在其中的2条染色体上。本研究中桂花OfACOs基因的42.6%主要分布在其中的5条染色体上,推测可能与基因复制有关。
通过同源序列比对筛选、不同组织部位及开花阶段的转录组测序分析,最终筛选到4个在花瓣衰老阶段(S5和S6)显著上调表达的成员LYG007706、LYG007045、LYG006223、LYG035696。LYG006223和LYG007045位于3号染色体上,LYG007706位于4号染色体上。家族内共线性分析发现:LYG007045与LYG007706为一对复制对,可见其保守基序、共线性都有紧密联系,可能具有相似的功能。LYG035696家族内进化分析中与LYG006223聚类较近,两者可能具有相似的功能。与拟南芥进化分析中,LYG007706与拟南芥5个基因位于同一个分组内。与拟南芥共线性分析中AT1G12010与LYG007045为共线性对;研究表明AT1G12010参与乙烯生物合成[32],通过负调控ACC2的表达来调控拟南芥内源乙烯水平能[33]。AT1G62380与LYG006223是一对共线性对 ;LYG007706不仅与AT1G62380、AT1G12010共线性,还与AT2G19590共线性。AT2G19590能够通过调控拟南芥乙烯生物合成,调控黑暗中拟南芥幼苗根系发育[34]。
此外,进化分析及基因结构分析表明:桂花中有16个OfACOs成员与拟南芥AtACOs同时聚类于第3组中,其中11个OfACOs成员的Motif完全相同,且与拟南芥的AtACOs含有7个相同的Motif基序。通过转录组测序及RT-qPCR分析,在第3组中筛选到LYG007706在桂花花瓣衰老后期(S6)显著上调表达,与ZOU等[6]研究中乙烯释放峰(脱落期)一致,推断其可能参与乙烯途径的桂花花瓣衰老调控。由此推断,第3组中与拟南芥AtACOs聚类的10个其他OfACOs成员可能参与桂花其他组织部位的调控。
-
本研究通过对桂花OfACOs基因家族的鉴定与表达分析,筛选到4个在开花后期显著差异上调的成员:LYG006223、LYG007706、LYG007045和LYG035696,它们可能参与桂花花瓣衰老的调控。其中,LYG006223和LYG007045都定位在3号染色体上,且有着相同的保守基序;LYG007706定位在4号染色体上;LYG035696定位于21号染色体。后续将进一步验证这4个成员的功能,探索其参与桂花花瓣乙烯合成途径及衰老的调控机制。
Identification and expression of OfACOs gene family in Osmanthus fragrans
-
摘要:
目的 对桂花Osmanthus fragrans乙烯合成路径氨基环丙烷羧酸氧化酶(1-aminocyclopropane-1-carbox-ylate oxidase, ACO)基因家族进行了全基因组鉴定及表达分析,以探索参与桂花花瓣乙烯合成的关键ACO家族成员。 方法 以桂花品种‘柳叶金桂’O. fragrans‘Liuye Jingui’为参考基因组,对桂花OfACOs家族进行鉴定、进化分析、基因结构分析以及根、茎、叶、芽和不同开花阶段的表达模式分析。 结果 通过蛋白保守结构域(protein families, pfam)分析,从桂花基因组中共鉴定出了122个OfACOs成员,分布于22条染色体上;保守基序分析表明:大多数OfACOs基因同时包含 Motif 1、 Motif 2、Motif 3、Motif 4和Motif 7这5个保守基序,它们共同组成OfACOs保守结构域;共线性分析结果表明:与拟南芥Arabidopsis thaliana基因为共线性对的基因可能具有相似的功能。结合swiss-prot同源序列对比分析,筛选出27个成员作为候选研究对象,进行不同组织部位根、茎、叶、芽和不同开花阶段的转录组测序分析,结果发现:12个成员在花瓣中显著表达,其中LYG006223、LYG007706、LYG007045、LYG035696等4个成员在开花后期极显著差异上调表达。进一步对该4个成员进行荧光定量PCR验证分析,结果与转录组测序结果一致。 结论 通过OfACOs基因家族的全基因组鉴定及基因克隆分析,筛选到在开花后期显著差异上调的4个ACO家族成员可能参与桂花花瓣衰老的调控。图6参34 Abstract:Objective The objective of this study is to identify and analyze the gene family of OfACOs in ethylene biosynthesis pathway, so as to explore the key 1-aminocyclopropane-1-carbox-ylate oxidase (ACO) family members involved in the ethylene synthesis of Osmanthus fragrans petals. Method The identification, evolutionary analysis, gene structure analysis, and expression pattern analysis of OfACOs gene family at root, stem, leaf, bud and different flowering stages were carried out taking O. fragrans ‘Liuye Jingui’ as the reference genome. Result Through the analysis of protein conserved domain (protein families, pfam), a total of 122 OfACOs members were identified from O. fragrans genome, which were distributed on 22 chromosomes. The conserved motif analysis showed that most OfACOs genes contained Motif 1, Motif 2, Motif 3, Motif 4, and Motif 7, which together formed the OfACOs conserved domain. Collinearity analysis showed that OfACOs genes that were colinear with Arabidopsis thaliana genes might have similar functions. Combined with comparative analysis of switch-prot homologous sequence, 27 OfACOs members were selected as candidate research objects for transcriptome sequencing analysis of different tissue parts, including root, stem, leaf, bud and different flowering stages, and the results showed that 12 OfACOs members were significantly expressed in petals, among which LYG006223, LYG007706, LYG007045, and LYG035696 were significantly up-regulated in the late flowering stage. The four members were further analyzed by RT-qPCR analysis, and the results were consistent with those of transcriptome sequencing. Conclusion Through the whole genome identification and gene cloning analysis of OfACOs gene family, four members of OfACOs which are significantly up-regulated in the late flowering stage are screened, which may participate in O. fragrans petal senescence. [Ch, 6 fig. 34 ref.] -
Key words:
- Osmanthus fragrans /
- OfACOs /
- ethylene /
- petal senescence /
- gene family
-
图 1 桂花OfACOs基因家族的染色体定位
Figure 1 Chromosomal localization of OfACOs gene family in O. fragrans
图 2 桂花与拟南芥ACOs基因进化分析
Figure 2 Phylogenetic analysis of ACOs in O. fragrans and A. thaliana
图 3 桂花OfACOs基因结构及蛋白质保守结构域
Figure 3 Gene structure and conserved motifs analysis of OfACOs in O. fragrans
图 4 桂花及拟南芥ACO基因家族共线性分析
Figure 4 Collinearity analysis of ACO gene family in O. fragrans and A. thaliana
图 5 桂花OfACOs基因家族在桂花不同组织部位以及不同开花阶段中的表达分析
Figure 5 Expression profiles of OfACOs gene family in different tissues and different flowering stages of O. fragrans
-
[1] 武忠康. 桂花在园艺中的应用[J]. 现代园艺, 2022, 45(15): 134 − 135. WU Zhongkang. Application of Osmanthus fragrans in horticulture [J]. Contemporary Horticulture, 2022, 45(15): 134 − 135. [2] WU Lipeng, LIU Junyi, HUANG Weisu, et al. Exploration of Osmanthus fragrans Lour.’s composition, nutraceutical functions and applications [J/OL]. Food Chemistry, 2022, 377: 131853[2022-11-28]. doi: 10.1016/j.foodchem.2021.131853. [3] WANG Limei, LI Maoteng, JIN Wenwen, et al. Variations in the components of Osmanthus fragrans Lour. essential oil at different stages of flowering [J]. Food Chemistry, 2009, 114(1): 233 − 236. [4] WU Lichen, CHANG Lihui, CHEN Sihan, et al. Antioxidant activity and melanogenesis inhibitory effect of the acetonic extract of Osmanthus fragrans: a potential natural and functional food flavor additive [J]. LWT-Food Science and Technology, 2009, 42(9): 1513 − 1519. [5] CAI Xuan, MAI Rongzhang, ZOU Jingjing, et al. Analysis of aroma-active compounds in three sweet osmanthus (Osmanthus fragrans) cultivars by GC-olfactometry and GC-MS [J]. Journal of Zhejiang University Science B, 2014, 15(7): 638 − 648. [6] ZOU Jingjing, ZHOU Yuan, CAI Xuan, et al. Increases in DNA fragmentation and role of ethylene during the petal senescence of Osmanthus fragrans [J]. Postharvest Biology and Technology, 2014, 93: 97 − 105. [7] ZOU Jingjing, CAI Xuan, WANG Caiyun. The spatial and temporal distribution of programmed cell death (PCD) during petal senescence of Osmanthus fragrans [J]. Acta Horticulturae, 2017, 1185(39): 315 − 324. [8] 洪方蕾, 陆瑶, 俞世姣, 等. 桂花OfABFs基因克隆和表达分析[J/OL]. 浙江农林大学学报, 2023[2022-11-28]. doi: 10.11833/j.issn.2095-0756.20220264. HONG Fanglei, LU Yao, YU Shijiao, et al. Cloning and expression analysis of OfABFs gene in Osmanthus fragrans [J/OL]. Journal of Zhejiang A&F University, 2023, 40(3)[2022-11-28]. doi: 10.11833/j.issn.2095-0756.20220264. [9] 蒋琦妮, 付建新, 张超, 等. 桂花OfAP1基因的克隆及表达分析[J]. 浙江农林大学学报, 2019, 36(4): 664 − 669. JIANG Qini, FU Jianxin, ZHANG Chao, et al. CDNA cloning and expression analysis of OfAP1 in Osmanthus fragrans [J]. Journal of Zhejiang A&F University, 2019, 36(4): 664 − 669. [10] 向其柏, 刘玉莲. 中国桂花品种图志[M]. 浙江: 浙江科学技术出版社, 2008: 93 − 260. XIANG Qibo, LIU Yulian. An Illustrated Monograph of the Sweet Osmanthus Variety in China [M]. Hangzhou: Zhejiang Science & Technology Press, 2008: 93 − 260. [11] 朱诚, 曾广文. 桂花花衰老过程中的某些生理生化变化[J]. 园艺学报, 2000, 27(5): 356 − 360. ZHU Cheng, ZENG Guangwen. Physiological and biochemical changes in flower senescence of Osmanthus fragrans Lour. [J]. Acta Horticulturae Sinica, 2000, 27(5): 356 − 360. [12] ZHOU Yuan, CHENG Zhengwei, WANG Caiyun. Effects of exogenous ethylene and ethylene inhibitor on longevity and petal senescence of sweet osmanthus [J]. Acta Horticulturae, 2008, 768: 487 − 493. [13] PENG Hsiaoping, LIN Teryun, WANG Ningning, et al. Differential expression of genes encoding 1-aminocyclopropane-1-carboxylate synthase in Arabidopsis during hypoxia [J]. Plant Molecular Biology, 2005, 58(1): 15 − 25. [14] ADAMS D O, YANG Shangfa. Methionine metabolism in apple tissue: implication of S-adenosylmethionine as an intermediate in the conversion of methionine to ethylene [J]. Plant Physiology, 1977, 60(6): 892 − 896. [15] SORNCHAI P, van DOORN W G, IMSABAI W, et al. Dendrobium orchids carrying antisense ACC oxidase: small changes in flower morphology and a delay of bud abortion, flower senescence, and abscission of flowers [J]. Transgenic Research, 2020, 29(4): 429 − 442. [16] LOVE J, BJORKLUND S, VAHALA J, et al. Ethylene is an endogenous stimulator of cell division in the cambial meristem of Populus [J]. Proceedings of the National Academy of Sciences of the United States of America, 2009, 106(14): 5984 − 5989. [17] 金乌云. 甜瓜ACO基因家族成员的鉴定及CmACO7和CmACO8基因的克隆[D]. 呼和浩特: 内蒙古大学, 2017. JIN Wuyun. Identification of ACO Gene Family and Cloning of CmACO7 and CmACO8 Gene in Melon [D]. Hohhot: Inner Mongolia University, 2017. [18] CALVO A P, NICOLÁS C, NICOLÁS G, et al. Evidence of a cross-talk regulation of a GA 20-oxidase (FsGA20ox1) by gibberellins and ethylene during the breaking of dormancy in Fagus sylvatica seeds [J] Physiologia Plantarum, 2004, 120(4) : 623 − 630. [19] PARK C H, ROH J, YOUN J, et al. Arabidopsis ACC oxidase-1 coordinated by multiple signals mediates ethylene biosynthesis and is involved in root development [J]. Molecules and Cells, 2018, 41(10): 923 − 932. [20] CHEN Hongguo, ZENG Xiangling, YANG Jie, et al. Whole-genome resequencing of Osmanthus fragrans provides insights into flower color evolution[J/OL]. Horticulture Research, 2021, 8: 98[2022-11-28]. doi: 10.1038/s41438-021-00531-0. [21] 冯志聪, 邹航, 果弘毅, 等. 旱柳、杞柳ACO基因家族的全基因组鉴定及表达分析[J]. 扬州大学学报(农业与生命科学版), 2022, 43(2): 10 − 19. FENG Zhicong, ZOU Hang, GUO Hongyi, et al. Genome-wide identification and expression analysis of ACO gene family inSalix matsudana and Salix integra [J]. Journal of Yangzhou University (Agricultural and Life Science Edition), 2022, 43(2): 10 − 19. [22] MARCHEL-BAUER A, DERBYSHIRE M K, GONZALES N R, et al. CDD: NCBI’s conserved domain database [J/OL]. Nucleic Acids Research, 2015, 43: D222-6[2022-11-28]. doi: 10.1093/nar/gku1221. [23] PEI Jimin. Multiple protein sequence alignment [J]. Current Opinion in Structural Biology, 2008, 18(3): 382 − 386. [24] VOORRIPS R E. MapChart: software for the graphical presentation of linkage maps and QTLs [J]. The Journal of Heredity, 2002, 93(1): 77 − 78. [25] HE Zilong, ZHANG Huangkai, GAO Shenghan, et al. Evolview v2: an online visualization and management tool for customized and annotated phylogenetic trees [J/OL]. Nucleic Acids Research, 2016, 44(W1): W236-41[2022-11-28]. doi: 10.1093/nar/gkw370. [26] WANG Yupeng, TANG Haibao, DEBARRY J D, et al. MCScanX: a toolkit for detection and evolutionary analysis of gene synteny and collinearity [J/OL]. Nucleic Acids Research, 2012, 40(7): e49[2022-11-28]. doi: 10.1093/nar/gkr1293. [27] 郭旭, 张慧莹, 王铮, 等. 绿豆VrWOX基因家族鉴定及表达分析[J]. 生物工程学报, 2023, 39(2): 566 − 585. GUO Xu, ZHANG Huiying, WANG Zheng, et al. Molecular characterization and transcriptional analysis of VrWOX genes in mungbean [Vigna radiate (L.) Wilczek] [J]. Chinese Journal of Biotechnology, 2023, 39(2): 566 − 585. [28] HOUBEN M, van de POEL B. 1-aminocyclopropane-1-carboxylic acid oxidase (ACO): the enzyme that makes the plant hormone ethylene [J/OL]. Frontiers in Plant Science, 2019, 10: 695[2022-11-28]. doi: 10.3389/fpls.2009.00695. [29] 朱丽颖, 杨军, 罗朝鹏, 等. 烟草ACO基因家族鉴定和二氯喹啉酸药害条件下的表达分析[J]. 烟草科技, 2022, 55(9): 10 − 18. ZHU Liying, YANG Jun, LUO Chaopeng, et al. Identification of tobacco ACO gene family and its expression analysis under quinclorac phytotoxicity [J]. Tobacco Science &Technology, 2022, 55(9): 10 − 18. [30] 杨利艳, 边璐, 史瑞翔, 等. 玉米ACO家族的全基因组鉴定与表达分析[J]. 湖南师范大学自然科学学报, 2022, 45(3): 109 − 116. YANG Liyan, BIAN Lu, SHI Ruixiang, et al. Genome-wide identification and expression analysis of ACO family in maize [J]. Journal of Natural Science of Hunan Normal University, 2022, 45(3): 109 − 116. [31] WEI Hengling, XUE Yujun, CHEN Pengyun, et al. Genome-wide identification and functional investigation of 1-aminocyclopropane-1-carboxylic acid oxidase (ACO) genes in cotton [J/OL]. Plants, 2021, 10(8): 1699[2022-11-28]. doi: 10.3390/plants10081699. [32] HUDGINS J W, RALPHY S G, FRANCESCHI V R, et al. Ethylene in induced conifer defense: cDNA cloning, protein expression, and cellular and subcellular localization of 1-aminocyclopropane-1-carboxylate oxidase in resin duct and phenolic parenchyma cells [J]. Planta, 2006, 224(4): 865 − 877. [33] MOON J, KIM S Y, PARK C, et al. BES1 negatively regulates the expression of ACC oxidase 2 to control the endogenous level of ethylene in Arabidopsis thaliana [J/OL]. Plant Signaling & Behavior, 2020, 16(2): 1850625[2022-11-28]. doi: 10.1080/15592324.2020.1850625. [34] GU Shinyuan, WANG Longchi, CHEUH Chiaomei, et al. CHITINASE LIKE1 regulates root development of dark-grown seedlings by modulating ethylene biosynthesis in Arabidopsis thaliana [J/OL]. Frontiers in Plant Science, 2019, 10: 600[2022-11-28]. doi: 10.3389/fpls.2019.00600. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20220783
 点击查看大图
点击查看大图

计量
- 文章访问数: 2018
- HTML全文浏览量: 475
- PDF下载量: 106
- 被引次数: 0
