





-
低温是影响植物生长发育、限制植物生产力和世界各地物种分布的重要因素之一[1−3]。低温胁迫包括寒冷(0~15 ℃)胁迫和冷冻(<0 ℃)胁迫[4]。冷应激通常会导致植物生理变化,如与离子泄漏相关的细胞膜损伤、脯氨酸(Pro)含量的变化、活性氧自由基(ROS)的积累等[5−8],同时会刺激一些抗应激酶的积累,如超氧化物歧化酶(SOD)以及过氧化物酶(POD)的积累[9−11],作为保护酶系统,以限制自由基的水平,保持抗氧化剂和自由基之间的平衡。在已知的低温胁迫基因调控模型中,冷应激首先作用于信号感知和转导途径,诱导转录调控,从而激活多种冷调节蛋白(COR) [12−13],以提高植物对环境胁迫的耐受性[14]。C-环肽结合转录因子(CBF)途径是植物体内重要的、研究得较清楚的冷响应途径,该途径通过 ICE-CBF-COR信号通路介导[15−16]。低温胁迫下CBF基因的表达受多个转录因子的正负调控[17],ICE(inducer of CBF expression)基因是CBFs[C-repeat-binding factors,又称dehybration responsive element factors (DREBs)]冷响应通道上游调控因子,能够诱导CBF基因的表达,提高植物的低温适应能力[4, 18]。
ICE 是植物体内的一类 bHLH转录因子,含有高度保守的碱性螺旋-环-螺旋(bHLH)结构域,该结构域包含ICE特异性序列KMDRASILGDAID/EYLKELL[19−20]。目前已经在水稻Oryza sativa、龙眼Dimocarpus longan和山定子Malus baccata等物种中鉴定出ICE基因,并有研究表明龙眼DlICE1和山定子MbICE1的过表达分别增加了转基因烟草Nicotiana benthamiana[21]和拟南芥Arabidopsis thaliana[22]的耐寒性。此外,过表达水稻OsICE1同时提高了拟南芥的耐寒性[23]、耐旱性和光合作用效率[24]。
毛竹Phyllostachys edulis是生长最快的植物之一[25],具有很高的经济、生态和社会价值[26−27]。由于其独特的“爆炸性生长”特征,被认为是21世纪最具潜力的植物种类[28]。研究竹子的胁迫响应基因有助于提高其抗逆性。目前对竹子非生物胁迫响应潜在的分子机制知之甚少[29]。毛竹中已鉴定出 153 个具有完整保守结构域的 bHLH 基因家族成员[30],它们在毛竹不同组织和不同生长发育时期有不同程度的表达,参与植物许多非生物胁迫调控,但尚未在毛竹bHLH转录因子家族鉴定出ICE亚家族。本研究将对毛竹ICE基因家族进行全基因组鉴定,分析其在低温胁迫条件下的表达模式,鉴定响应毛竹抗寒的关键家族成员,为提高毛竹抗寒性奠定基础。
-
在GigaDB网站下载毛竹所需的数据文件。在Pfam数据库中以 ICE 隐马尔可夫模型(Profile HMM)为模板下载结构域数据[31],以此为种子模型利用HMMER3检索本地毛竹蛋白数据库,设置 E≤1×10−20[32],获得候选基因家族成员。利用SMART[33]和美国国家生物技术信息中心(NCBI) Blast对已鉴定的ICE结构特性全面分析,最终获得毛竹ICE基因家族成员。
-
通过TBtools软件从毛竹全基因组数据库中,提取ICE家族成员的基因编码区序列(CDS)、蛋白fasta序列以及基因结构和位置信息等[34]。利用在线工具Prot Param和TargetP 2.0 Server获取已鉴定的毛竹ICE蛋白序列的氨基酸数量、分子量、等电点以及信号肽等数据。
-
从文献中获取不同物种已鉴定的ICE基因编号,在NCBI中下载得到玉米Zea mays、小麦Triticum aestivum、高粱Sorghum bicolor的ICE基因[35],水稻(
https://www.ricedata.cn/gene/ )和拟南芥的ICE基因在各自基因组数据库中下载。根据获取的5种植物ICE氨基酸序列,通过MEGA7的MUSCLE进行多序列比对并采用邻接法(NJ)构建系统进化树,自举评估(Bootstrap)重复1 000次。 -
保守基序采用在线工具 MEME进行预测,利用TBtools软件的Biosequence Structure illustrator和show Gene On Chromose插件分析内含子、外显子、染色体位置信息并可视化绘图。
-
通过TBtools 软件BLAST模块进行毛竹基因组所有蛋白的自身序列比对以及物种间基因组蛋白序列两两比对,使用 MCScanX分析ICE家族共线性关系并使用Circos 0.69-9将其结果可视化。
-
利用PlantCARE网站对毛竹ICE基因家族成员转录起始位点上游1 500 bp的启动子区域进行顺式作用元件分析。对其结果筛选排序后利用 Tbtools 中Simple Bio Sequence Viewer模块进行可视化。
-
以在温度25 ℃,光照强度4 000 lx,光照16 h/黑暗8 h培养条件下生长50 d的毛竹为材料,在4、0、−2 ℃下进行低温处理,光热条件不变。分别在0(对照)、0.5、1.0、24.0、48.0 h取从上往下数第2、3叶片,脯氨酸(Pro)采用茚三酮显色法测定,POD采用愈创木酚显色法[36],SOD活性采用氮蓝四唑(NBT)法测定[37]。
-
以正常生长的50 d毛竹幼苗为材料,在4、0、−2 ℃下低温处理,0、0.5、1.0、24.0、48.0 h取毛竹从上往下数第2、3叶片,使用RNA提取试剂盒提取总RNA,利用翌圣生物科技公司Hifair® Ⅲ 1st Strand cDNA Synthesis SuperMix for qPCR(gDNA digester plus)试剂盒合成cDNA,Primer 3在线设计 ICE基因RT-qPCR引物,使用翌圣生物科技公司的Hieff® qPCR SYBR® Green Master Mix (No Rox)试剂盒进行 RT-qPCR 试验,反应体系和程序参照试剂说明书,以毛竹肌动蛋白(ACTIN)作为内参基因[38]。采用2−∆∆Ct方法计算相对基因表达水平,Graphpad可视化绘图。
-
数据统计学分析采用 SPSS 24.0进行单因素方差分析(ANOVA)和独立样本t检验。
-
根据基因的染色体定位信息,把获得的4个毛竹ICE基因命名为PeICE1~4。理化性质分析(表1)发现:最大蛋白分子量为53.57 kDa,最小蛋白分子量为38.21 kDa。氨基酸序列长度为367~521个氨基酸。等电点为5.23~5.56。4个ICE基因都是酸性蛋白(理论等电点小于7),不稳定指数范围为 56.15~60.03,脂肪族氨基酸指数显示:该家族蛋白的热稳定性为72.61~75.69,亲水性平均值显示其均为亲水性蛋白。
表 1 PeICEs 理化性质分析
Table 1. Physicochemical properties of PeICEs
基因名称 等电点 相对分
子量/ kDa氨基酸
数量/个脂溶
指数亲水
指数不稳定
系数PeICE1 5.56 53.19 516 72.97 −0.237 56.15 PeICE2 5.48 53.57 521 72.61 −0.274 56.56 PeICE3 5.23 38.98 375 72.77 −0.162 56.81 PeICE4 5.23 38.21 367 75.69 −0.129 60.03 -
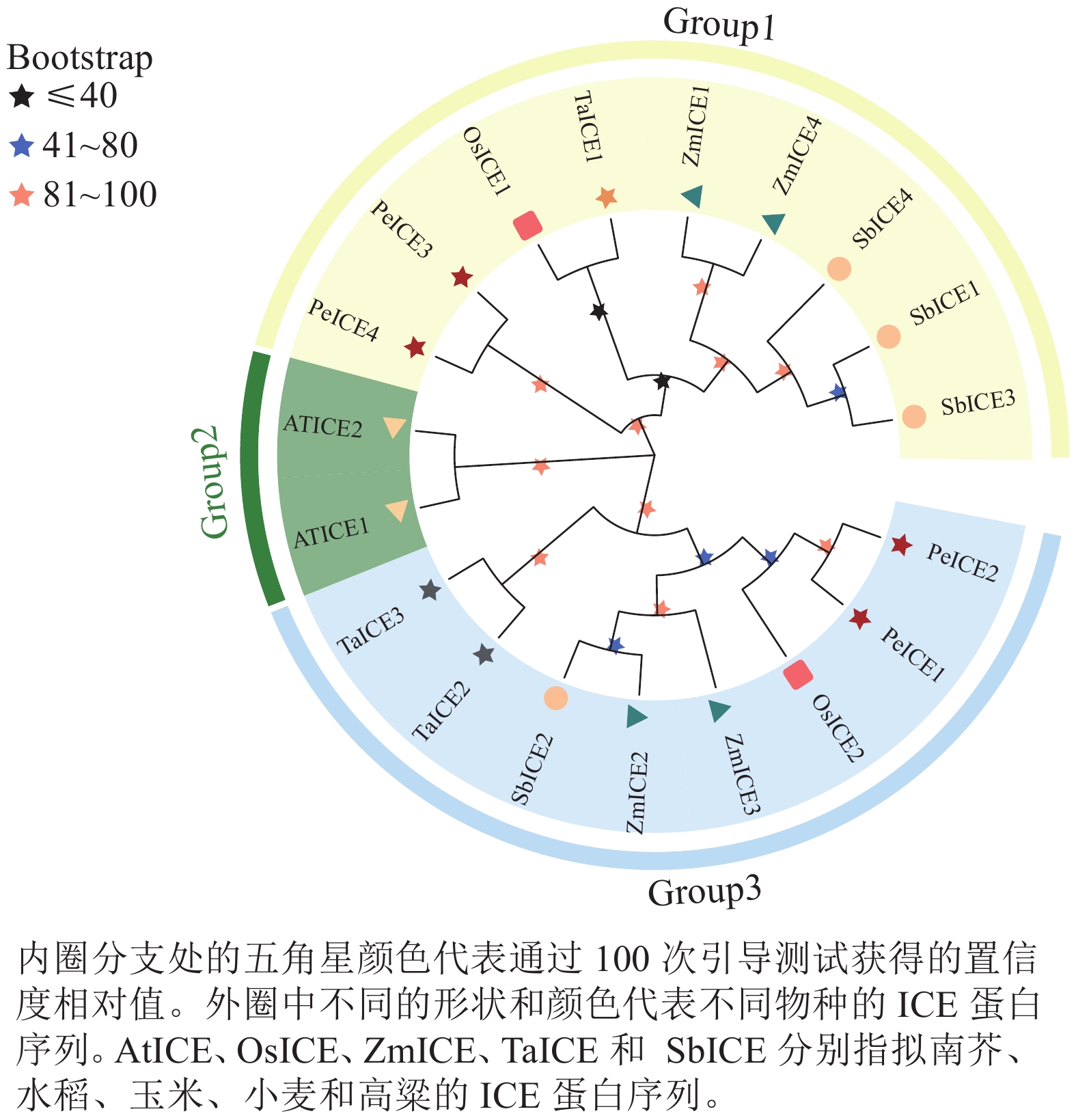
构建毛竹、拟南芥、水稻、小麦、高粱 ICE基因家族的系统进化树(图1)。将其分为3类,其中具有独特motif 5的PeICE1和PeICE2在第3类,具有独特motif 10的PeICE3和PeICE4被分在第1类,拟南芥单独被分在第2类。可以发现毛竹与水稻和玉米的亲缘关系较近,与拟南芥间的亲缘关系较远。
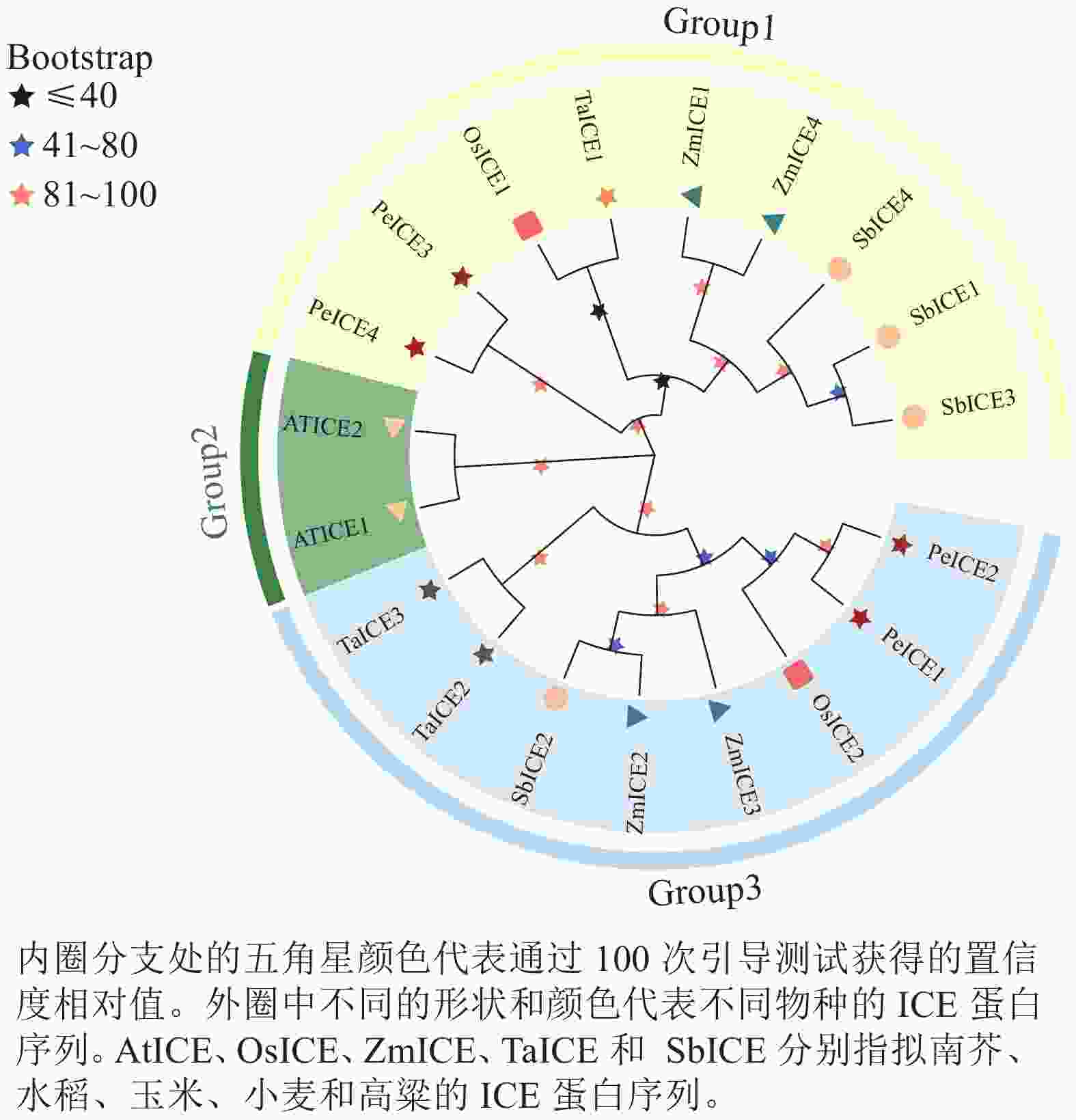
图 1 ICE基因家族进化分析
Figure 1. Evolutionary analysis of ICE gene families
-
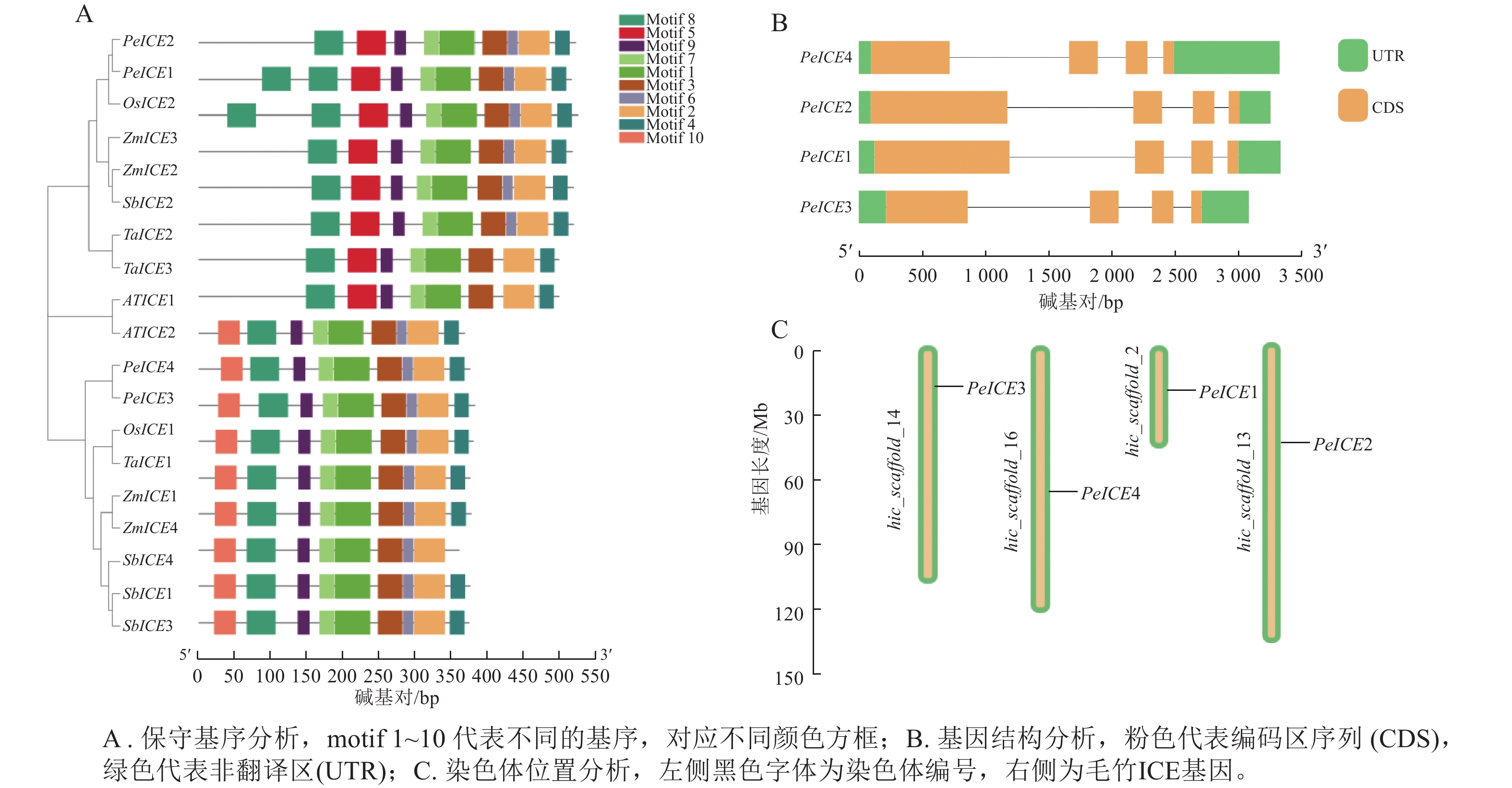
保守基序分析发现:19条 ICE 蛋白序列的保守基序的数量和排列具有一定相似性。19 条ICE均含有motif 1、motif 2、motif 3、motif 4、 motif 6、 motif 7、motif 8 、motif 9等8个保守基序。其中第3类群中的8个ICE具有第1类群没有的motif 5,PeICE1含有2个motif 5,第1类群中9个ICE则具有独特的motif 10,并且这些基序的排列顺序基本一致,说明这些基序在进化过程中高度保守(图2A)。PeICE基因结构分析发现:其内含子和外显子数量和排列基本一致(图2B),染色体分布(图2C)显示:4个PeICE基因均匀分布在4条染色体上。
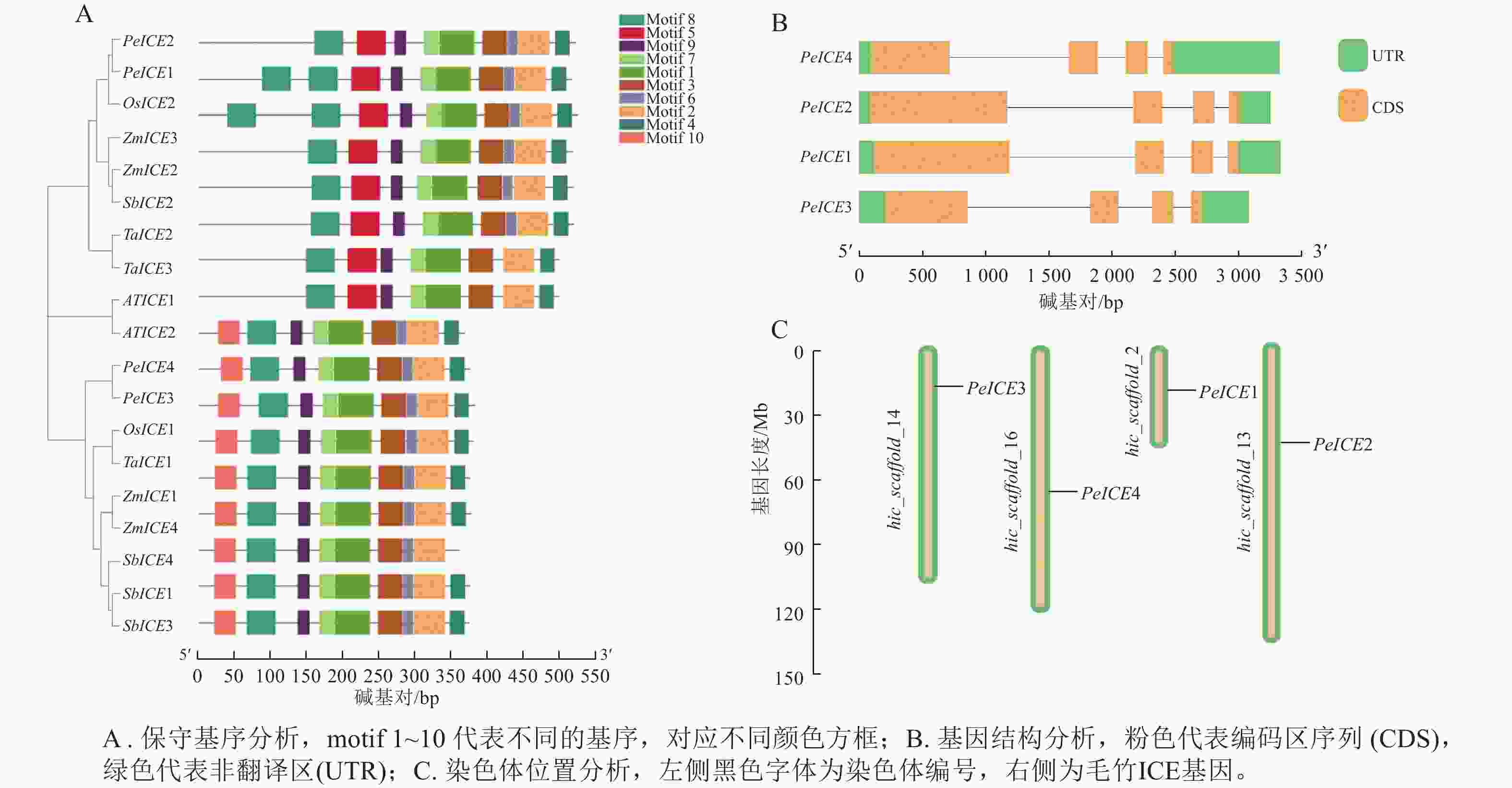
图 2 保守基序、基因结构及染色体位置分析
Figure 2. Conserved motif, gene structure and chromosome position
-
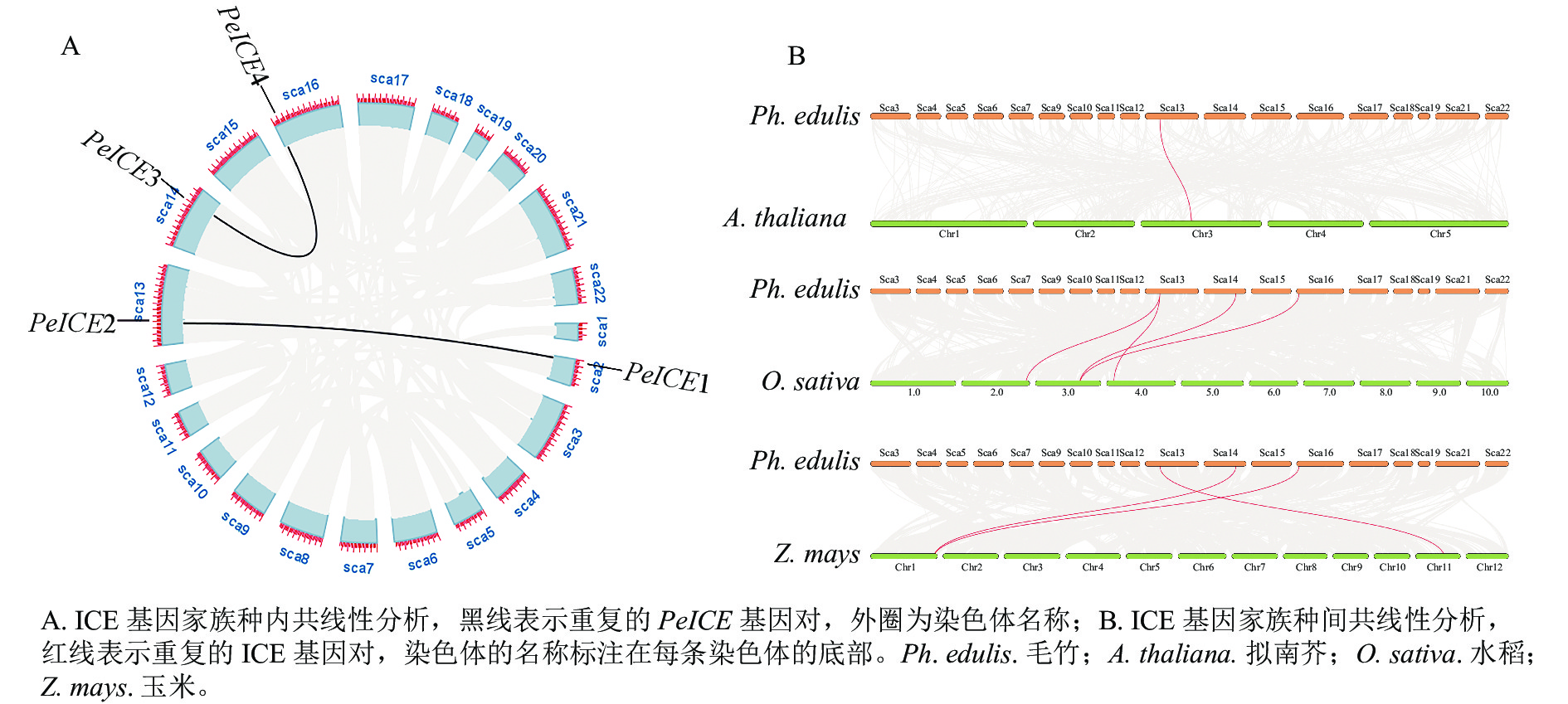
共线性分析发现:在毛竹基因组中,PeICE基因形成2个基因对(图3A),由片段复制产生,不存在串联重复基因。基因组间的共线性分析结果显示:毛竹与单子叶植物水稻、玉米之间的进化关系更近(图3B)。
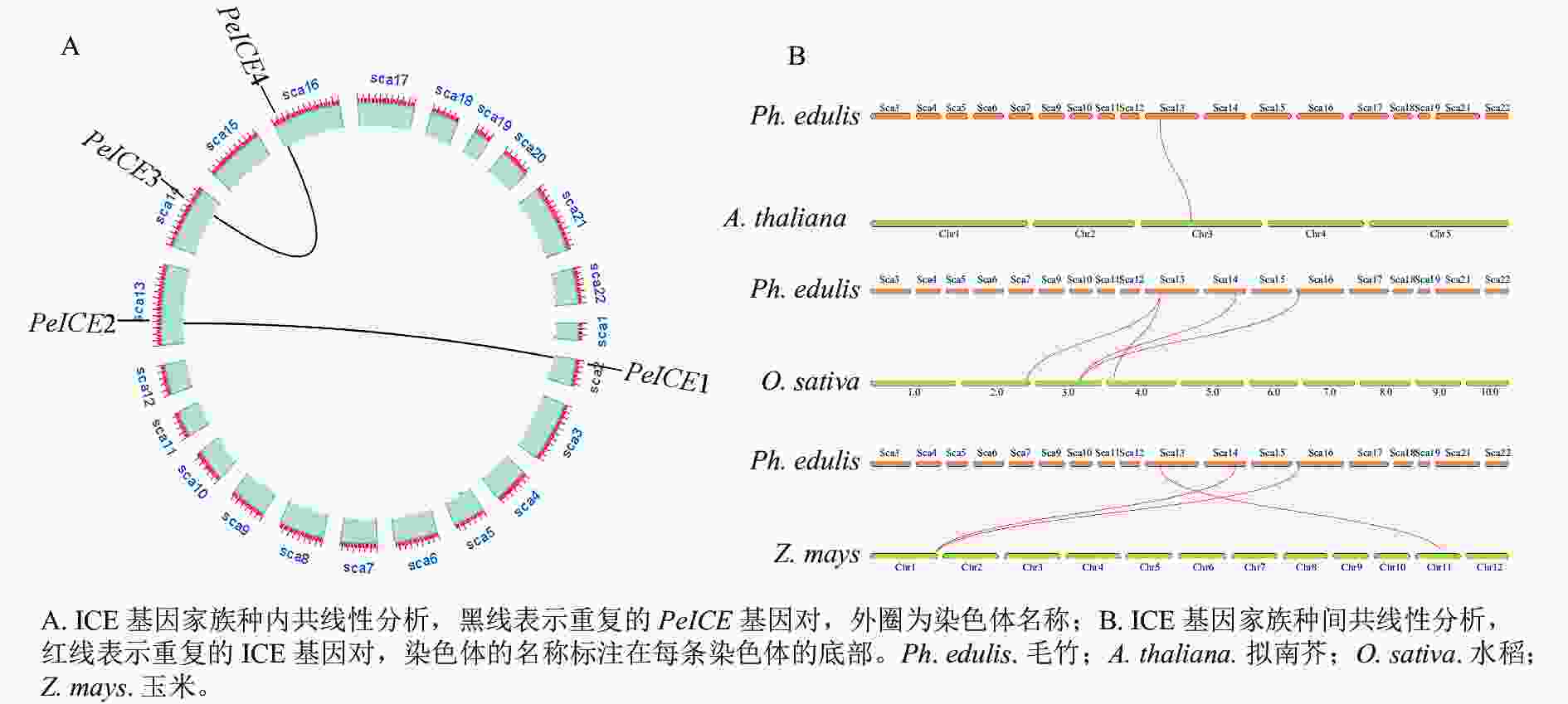
图 3 共线性分析
Figure 3. Collinearity analysis
-
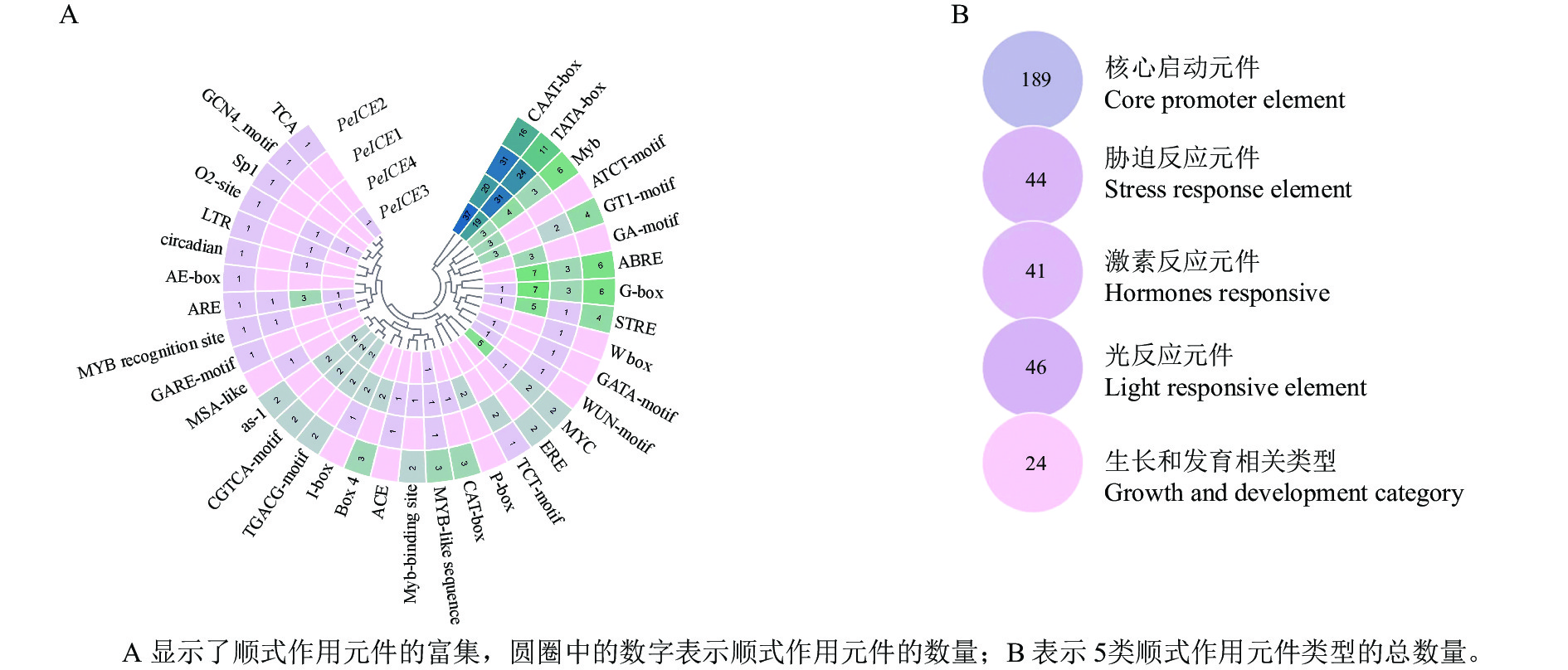
通过提取毛竹ICE基因上游1 500 bp 的启动子区域,利用PlantCARE进行顺式作用元件分析。结果(图4)表明:除了核心启动子元件(TATA-Box和CAAT-Box)外,还存在许多其他启动子元件。根据功能将这些顺式调控元件分为光反应元件、激素反应元件、生长和发育相关类型以及胁迫反应元件。光反应元件包括G-box、GT1-motif、Box 4、AE-box、Sp1、TCT-motif、I-box、ACE、GATA-motif、GA-motif和ATCT-motif ;激素反应元件包括水杨酸反应元件(TCA、as-1),赤霉素反应元件(包括P盒、GARE基序)、脱落酸反应元件(ABRE)、茉莉酸甲酯作用元件(TGACG-motif和CGTCA-motif)和乙烯反应原件(ERE);生长和发育类别包含分生组织表达相关的顺式作用调控元件(CAT-box)、干旱和 ABA 应答的顺式作用元件(MYC)、厌氧诱导反应(ARE),胚乳表达相关(GCN4基序),昼夜节律控制(circadian)、细胞周期调节(MSA-like)和玉米醇溶蛋白代谢相关(O2位点)元件;胁迫反应元件包括富含干旱和 ABA 应答的顺式作用元件(MYC)、干旱诱导元件(MBS)和低温反应元件(LTR)、糖代谢和植物防御信号传导的顺式作用元件(W-box)和伤口相关(WUN基序元件)、胁迫反应元件(STRE)。这些结果表明毛竹ICE基因表达受多种环境因子影响。
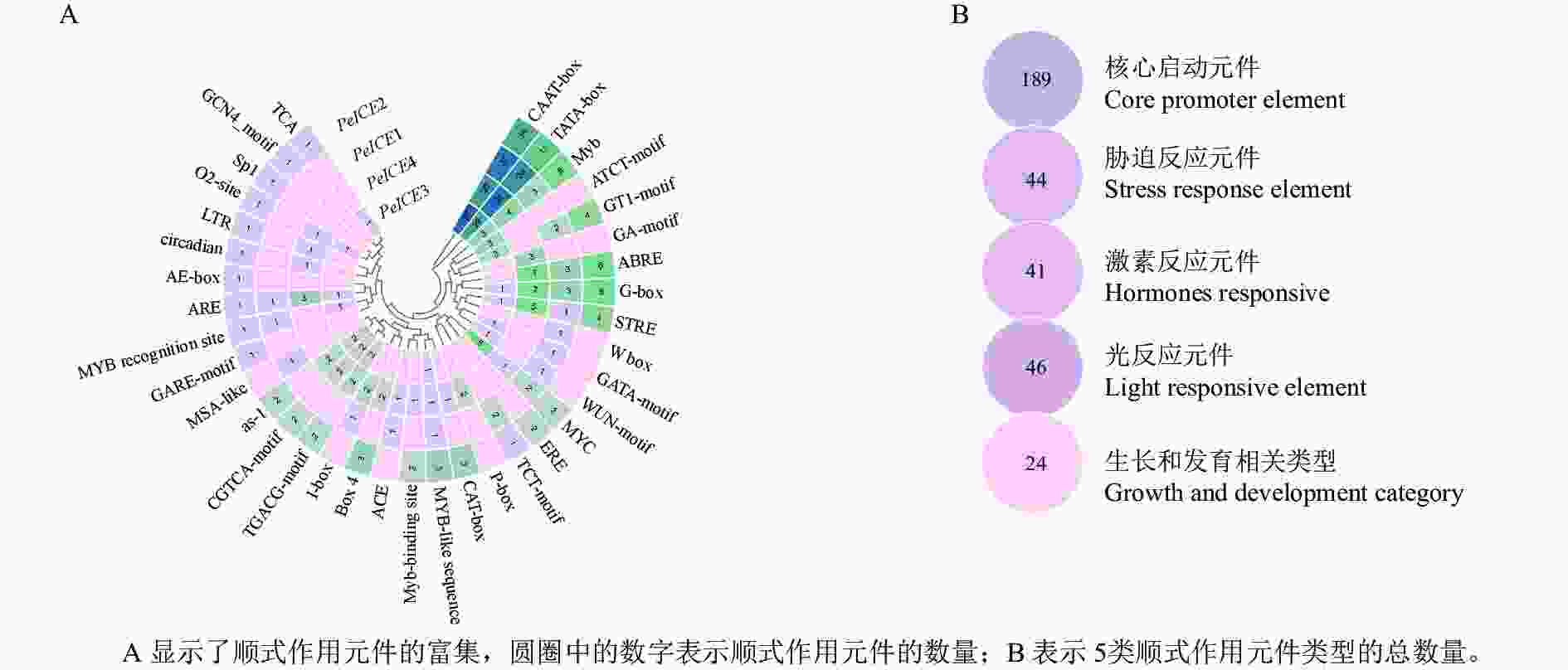
图 4 4个PeICsE启动子区域顺式作用元件分析
Figure 4. Analysis of cis-acting elements in four PeICEs promoter regions
-
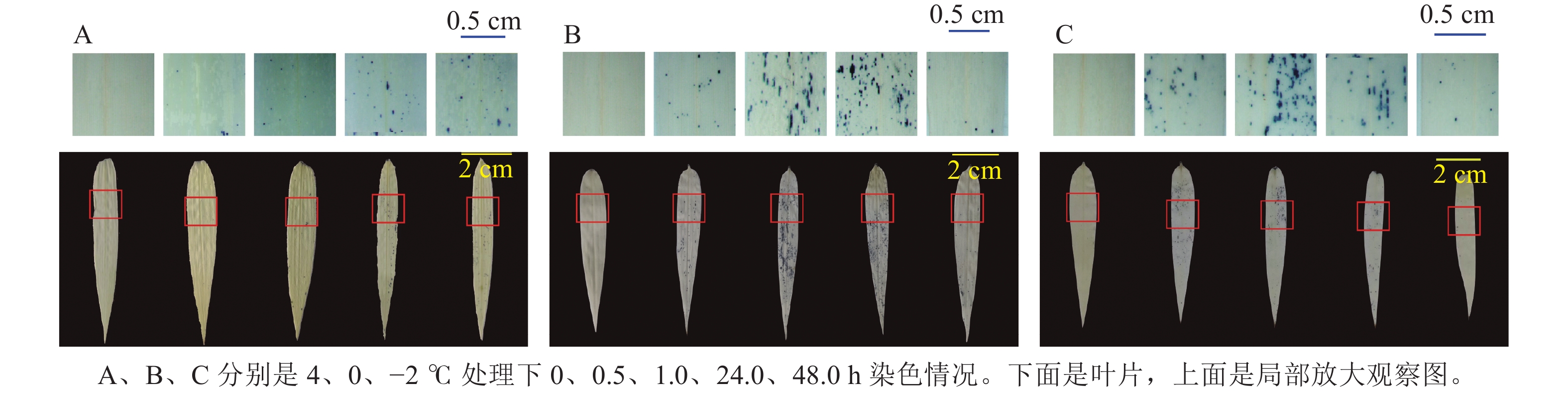
ROS染色(图5)发现:4 ℃时,随着处理时间增长,毛竹叶片上蓝色小圆点不断增多,而0和−2 ℃分别在处理24.0、1.0 h之内蓝色小圆点数量逐渐增多,之后逐渐减少。
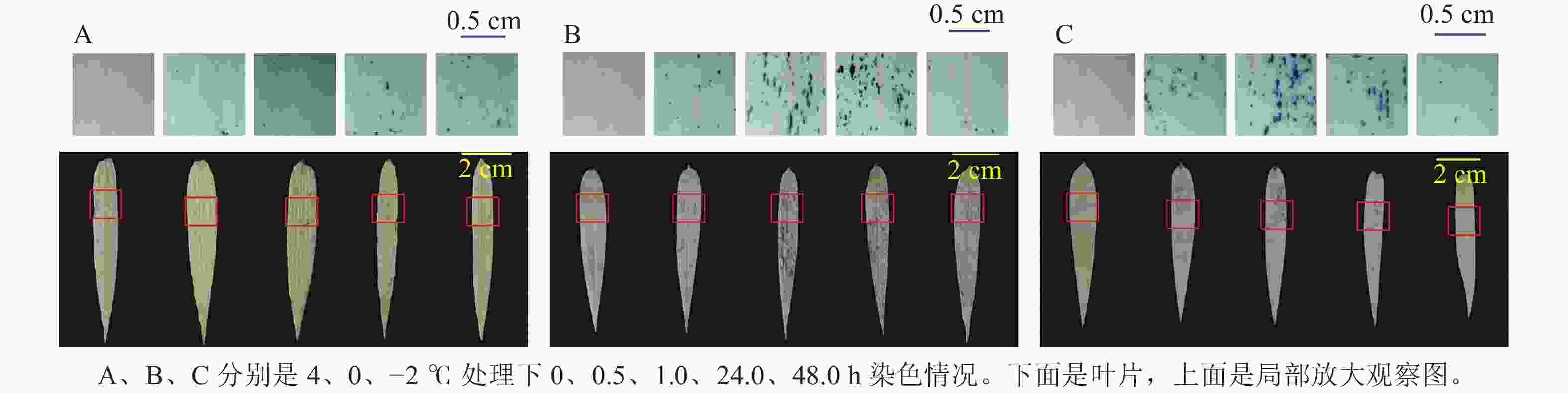
图 5 毛竹水培苗低温胁迫处理后活性氧自由基染色分析
Figure 5. ROS staining analysis of hydroponic seedlings of moso bamboo after low temperature stress treatments
-
由图6可见:4和0 ℃处理下,脯氨酸质量摩尔浓度48.0 h内显著增加(P<0.05),其中在24.0 h增加量最明显,随后减少,SOD活性48.0 h内显著增加。−2 ℃处理下,脯氨酸质量摩尔浓度均低于对照,SOD活性除24.0 h也均低于对照,但POD的活性除0.5 h均显著增加(P<0.05)。
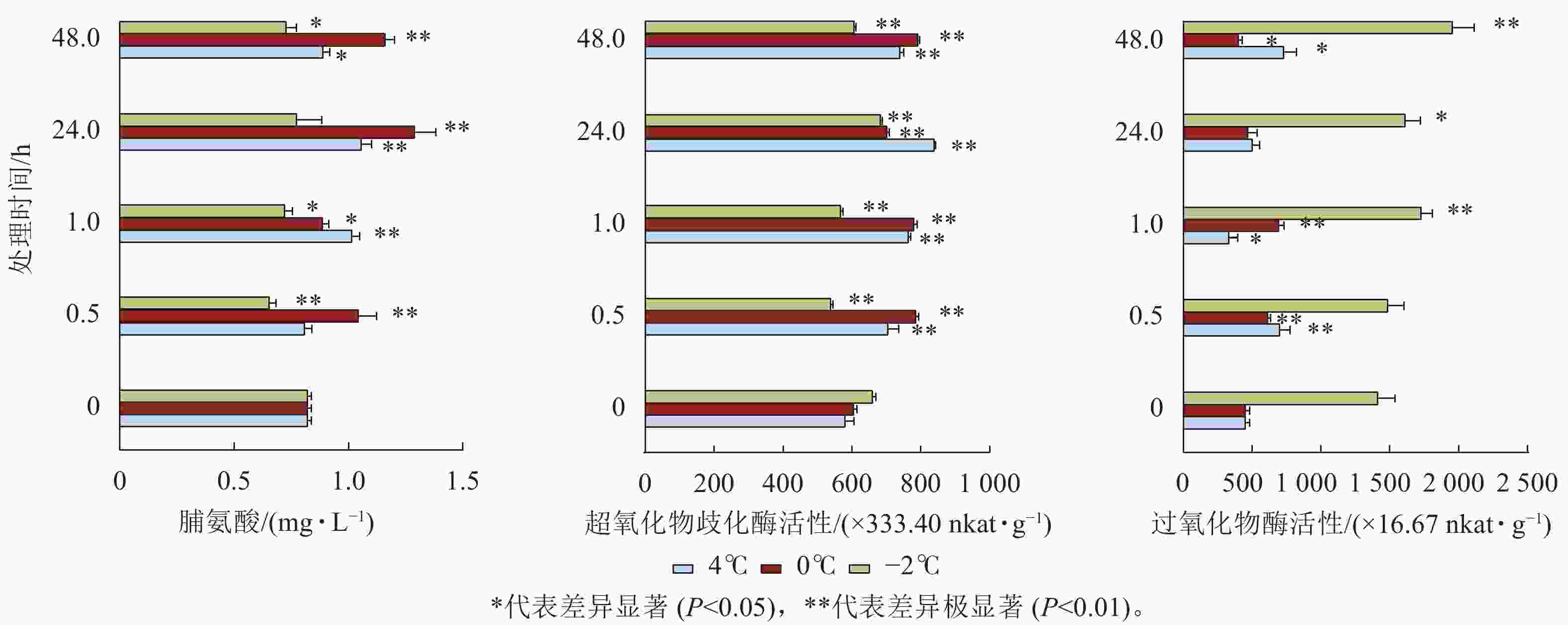
图 6 低温胁迫下毛竹脯氨酸质量摩尔浓度、超氧化物歧化酶活性和过氧化物酶活性
Figure 6. Proline, superoxide dismutase, and peroxidase content
-
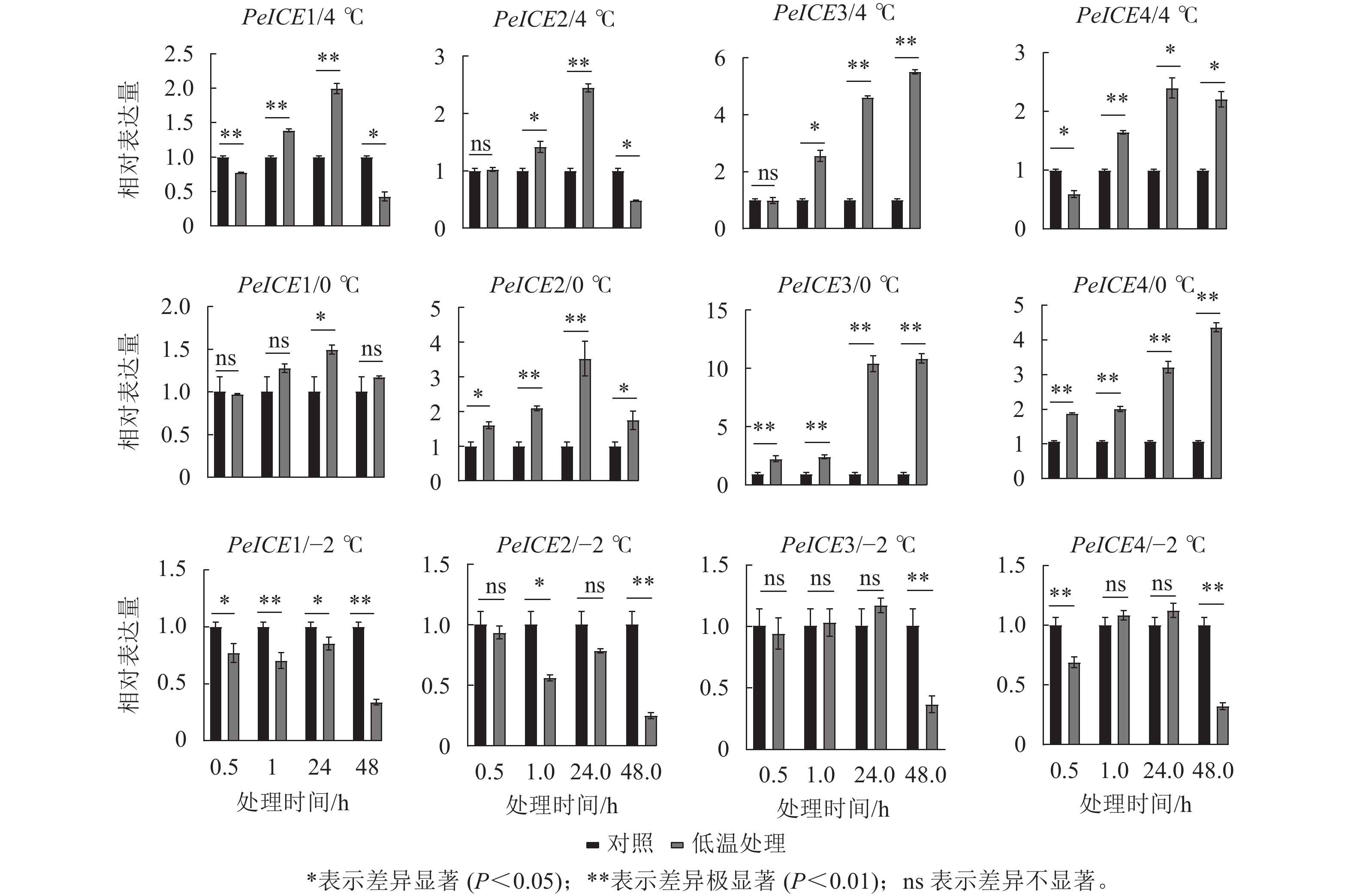
图7显示:在4 ℃处理下,除了PeICE3在48.0 h内表达量逐渐增加,其他PeICE基因都是24.0 h前表达量逐渐增加,之后下降。在0 ℃处理下,PeICE基因呈上升趋势,PeICE1、PeICE2与4 ℃趋势一致,且都以PeICE3增量最明显。在−2 ℃处理下,PeICE基因表达量低于对照或无显著差异。综上所述,PeICE3对低温胁迫最为敏感。
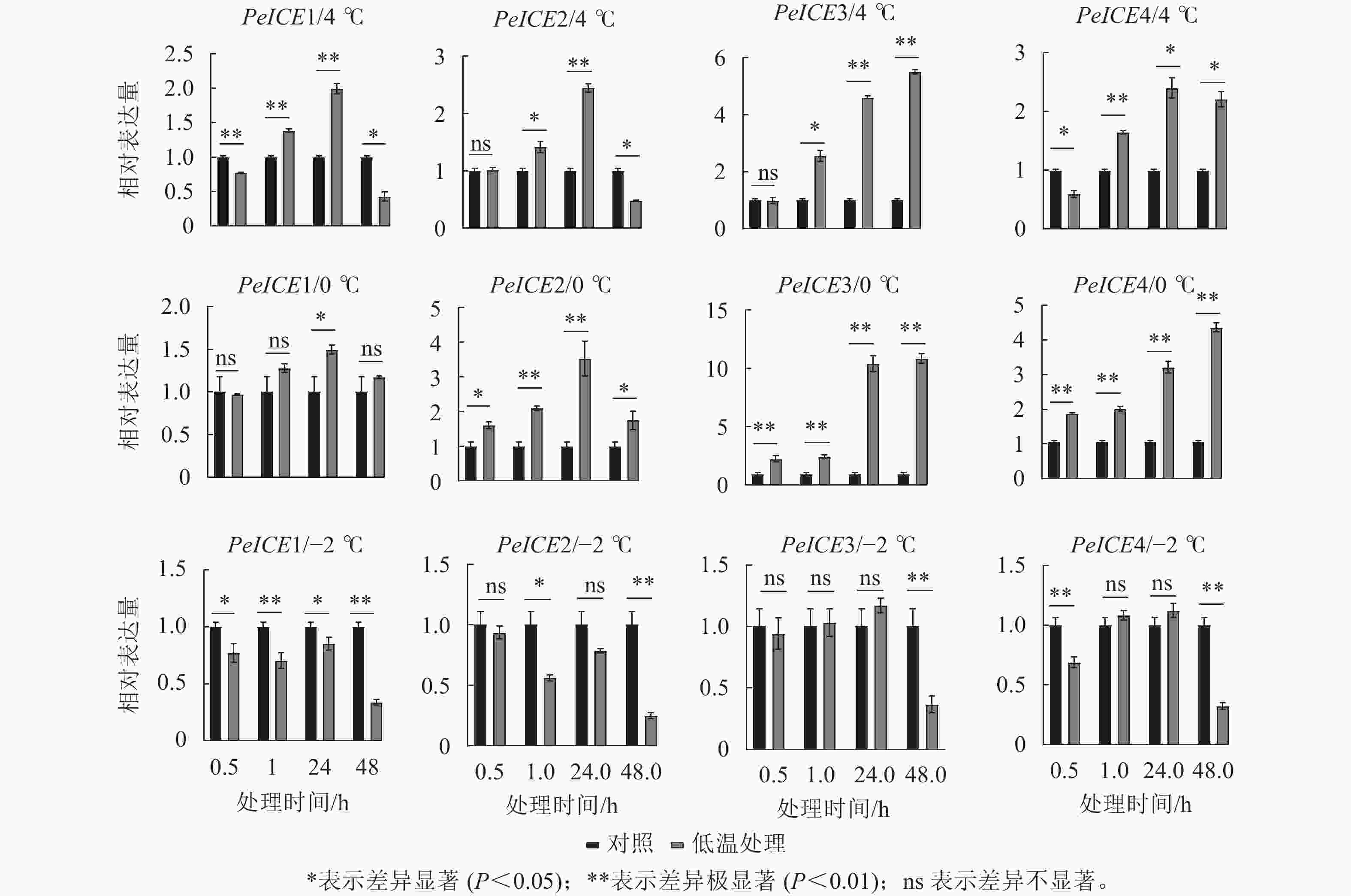
图 7 不同处理条件下PeICE基因相对表达量与对照的比较
Figure 7. Comparison the relative gene expression of PeICE at 0.5, 1.0, 24.0 and 48.0 h with control under 4, 0 and −2 ℃ treatment conditions
-
目前已在多种植物中鉴定出参与低温胁迫响应的ICE基因[39−40]。但尚未进行毛竹ICE基因家族系统鉴定,本研究鉴定了4个毛竹ICE基因家族成员。进一步的系统进化树和种间共线性分析发现:毛竹与单子叶植物水稻、玉米之间的进化关系更近。4个PeICE基因由片段复制产生,不存在串联重复基因,说明在进化过程中 ICE基因通过复制进行家族成员数量的扩张。保守结构域和基因结构分析发现:PeICE基因具有相似的保守基序和基因结构,表明其在进化过程中广泛保守。启动子顺式作用元件预测表明:毛竹ICE基因启动子区域普遍存在光响应、激素响应、生长发育及胁迫响应元件[41],表明ICE家族基因表达可能受到光照、植物激素的调控诱导或参与其信号反应途径,同时还可能参与毛竹生长发育的某些过程以及参与胁迫过程中的抗逆调控。
有研究认为:ROS不仅能损伤细胞,还能作为关键信号分子激活第二信使、感应基因转录和改变酶活性来调控植物的许多生理过程[42]。植物受到轻度胁迫时,体内酶促系统可有效提高ROS 的清除能力,保护植物免受环境胁迫的危害。本研究结果表明:毛竹受到寒冷胁迫时,随着温度降低和处理时间延长,其受到的伤害不断增加,体内SOD、POD活性显著增加以减轻ROS累积受到的损伤,与此同时PeICE基因表达量显著上升,其中PeICE3表达量上升最为显著,推测该基因可能参与毛竹ICE-CBF-COR调控途径抗寒。但当毛竹在冷冻胁迫下,体内ROS逐渐积累,SOD将不发挥作用,PeICE基因表达量低于对照或无显著差异,此时PeICE基因参与抗冻的可能性不大。
-
随着温度降低和处理时间加长,毛竹受到的损伤不断增强,体内酶活系统以及ICE基因积极响应低温胁迫。4个PeICE基因表达模式分析发现:转化PeICE3更容易获得耐低温的材料,研制出抗寒新品种。
Genome-wide identification of the ICE gene family in moso bamboo and its expression pattern under low temperature stress
-
摘要:
目的 对毛竹Phyllostachys edulis ICE基因家族进行鉴定及分析,找出响应毛竹抗寒关键家族成员,研究毛竹ICE基因的生物学功能、响应低温胁迫的分子机制及遗传转化,为提高毛竹抗寒性奠定理论基础。 方法 利用生物信息学方法分析毛竹ICE基因家族成员,并对4、0、−2 ℃低温处理0(对照)、0.5、1.0、24.0、48.0 h的毛竹生理指标和ICE基因的表达模式进行分析。 结果 共鉴定了4个毛竹ICE基因。保守结构域和多重序列比对分析表明:PeICE基因结构高度相似。系统发育关系及启动子顺式作用元件分析显示:PeICE基因与水稻Oryza sativa亲缘关系更近,同时存在大量与非生物胁迫相关的顺式作用元件。活性氧自由基(ROS)染色发现随着处理时间增长,ROS染色逐渐加深,但是其0 ℃处理24.0 h、−2 ℃处理1.0 h后染色逐渐减弱。脯氨酸(Pro)质量摩尔浓度、超氧化物歧化酶(SOD)活性显示:4和0 ℃条件下,Pro质量摩尔浓度和SOD活性整体增加,但−2 ℃时低于对照。过氧化物酶(POD)活性显示:在3个低温处理下均增加。ICE基因表达模式分析发现:4、0 ℃处理时PeICE表达量整体增加,且都以PeICE3 增量最明显;而−2 ℃处理下PeICE 整体表达量水平低于对照。 结论 随着温度降低和处理时间增强,毛竹受到的损伤不断增强,其内酶活系统以及ICE基因积极响应低温胁迫,其中,PeICE3对低温胁迫最为敏感,但在−2 ℃时,ICE基因表达量并未增加,推测该基因家族响应了寒冷胁迫而非冷冻胁迫。图7表1参42 Abstract:Objective The ICE gene family of Phyllostachys edulis (moso bamboo) was identified and analyzed to find out the key family members responding to cold resistance of moso bamboo, and to lay a theoretical foundation for further research on the biological functions of moso bamboo ICE genes, the molecular mechanisms responding to low-temperature stress, and the genetic transformation to improve the cold resistance of moso bamboo. Method Bioinformatics methods were used to identify and analyze the ICE gene family members of moso bamboo, and to analyze the physiological indexes and the expression patterns of ICE genes in moso bamboo at 0, 0.5, 1.0, 24.0, and 48.0 h of low-temperature treatments at 4, 0 and −2 ℃. Result A total of four mosaic bamboo ICE genes were identified. Conserved structural domains and multiple sequence comparison analyses showed that their structures were highly similar. Phylogenetic relationships and promoter cis-acting element analyses showed that the PeICE gene was more closely related to rice, and a large number of cis-acting elements associated with abiotic stresses were also present. Reactive oxygen radical (superoxide anion) staining revealed that the ROS staining gradually deepened with the increase in treatment time, but its staining gradually weakened after 24.0 h of 0 ℃ treatment and 1.0 h of −2 ℃ treatment. Proline content and superoxide dismutase activity assays revealed an overall increase in Pro and SOD content at 4 ℃ and 0 ℃, but their expression was lower than the control at −2 ℃, and peroxidase activity assays revealed an increase at all three temperatures. Analysis of ICE gene expression pattern revealed that the overall expression of PeICE increased in 4 ℃ and 0 ℃ treatment, and the most obvious increase was in PeICE3. The overall expression level of PeICE was lower than that of the control at −2 ℃. Conclusion With the decrease of cold treatment temperature and the enhancement of treatment time, moso bamboo was subjected to increasing damage, and its in vivo enzyme activity system as well as ICE genes responded positively to cold stress, and PeICE3 was the most sensitive to cold stress, whereas the expression of ICE genes did not increase at −2 ℃, and this gene family maybe respond to low-temperature stress instead of freezing stress. [Ch, 7 fig. 1 tab. 42 ref.] -
图 2 保守基序、基因结构及染色体位置分析
Figure 2 Conserved motif, gene structure and chromosome position
图 4 4个PeICsE启动子区域顺式作用元件分析
Figure 4 Analysis of cis-acting elements in four PeICEs promoter regions
图 5 毛竹水培苗低温胁迫处理后活性氧自由基染色分析
Figure 5 ROS staining analysis of hydroponic seedlings of moso bamboo after low temperature stress treatments
图 6 低温胁迫下毛竹脯氨酸质量摩尔浓度、超氧化物歧化酶活性和过氧化物酶活性
Figure 6 Proline, superoxide dismutase, and peroxidase content
图 7 不同处理条件下PeICE基因相对表达量与对照的比较
Figure 7 Comparison the relative gene expression of PeICE at 0.5, 1.0, 24.0 and 48.0 h with control under 4, 0 and −2 ℃ treatment conditions
表 1 PeICEs 理化性质分析
Table 1. Physicochemical properties of PeICEs
基因名称 等电点 相对分
子量/ kDa氨基酸
数量/个脂溶
指数亲水
指数不稳定
系数PeICE1 5.56 53.19 516 72.97 −0.237 56.15 PeICE2 5.48 53.57 521 72.61 −0.274 56.56 PeICE3 5.23 38.98 375 72.77 −0.162 56.81 PeICE4 5.23 38.21 367 75.69 −0.129 60.03  下载: 导出CSV
下载: 导出CSV
-
[1] BHAT K A, MAHAJAN R, PAKHTOON M M, et al. Low temperature stress tolerance: an insight into the omics approaches for legume crops [J/OL]. Frontiers in Plant Science, 2022, 13: 888710[2023-07-20]. doi: 10.3389/fpls.2022.888710. [2] LIU Yuanyuan, WU Chu, HU Xin, et al. Transcriptome profiling reveals the crucial biological pathways involved in cold response in moso bamboo (Phyllostachys edulis) [J]. Tree Physiology, 2019, 40(4): 538 − 556. [3] THEOCHARIS A, CLEMENT C, BARKA E A. Physiological and molecular changes in plants grown at low temperatures [J]. Planta, 2012, 235(6): 1091 − 1105. [4] YANG Chunbao, YANG Haizhen, XU Qijun, et al. Comparative metabolomics analysis of the response to cold stress of resistant and susceptible Tibetan hulless barley (Hordeum distichon) [J/OL]. Phytochemistry, 2020, 174: 112346[2023-07-20]. doi: 10.1016/j.phytochem.2020.112346. [5] EOM S H, AHN M A, KIM E, et al. Plant response to cold stress: cold stress changes antioxidant metabolism in heading type kimchi cabbage (Brassica rapa L. ssp. pekinensis) [J/OL]. Antioxidants, 2022, 11(4): 700[2023-07-20]. doi: 10.3390/antiox11040700. [6] WHITLOW T H, BASSUK N L, RANNEY T G, et al. An improved method for using electrolyte leakage to assess membrane competence in plant tissues [J]. Plant Physiology, 1992, 98(1): 198 − 205. [7] HAYAT S, HAYAT Q, ALYEMENI M N, et al. Role of proline under changing environments: a review [J]. Plant Signaling &Behavior, 2012, 7(11): 1456 − 1466. [8] WANG Haiyue, GUO Lin, ZHA Ruofei, et al. Histological, metabolomic and transcriptomic analyses reveal mechanisms of cold acclimation of the moso bamboo (Phyllostachys edulis) leaf [J]. Tree Physiology, 2022, 42(11): 2336 − 2352. [9] ZHOU Huang, ZHU Peilei, ZHONG Xiaojuan, et al. Transcriptome analysis of moso bamboo (Phyllostachys edulis) reveals candidate genes involved in response to dehydration and cold stresses [J/OL]. Frontiers in Plant Science, 2022, 13[2023-07-20]. doi: 10.3389/fpls.2022.960302. [10] ABID M, TIAN Z, ATA-UL-KARIM S T, et al. Improved tolerance to post-anthesis drought stress by pre-drought priming at vegetative stages in drought-tolerant and -sensitive wheat cultivars [J]. Plant Physiology and Biochemistry, 2016, 106: 218 − 227. [11] MILLER G, SUZUKI N, CIFTCI-YILMAZ S, et al. Reactive oxygen species homeostasis and signalling during drought and salinity stresses [J]. Plant,Cell &Environment, 2010, 33(4): 453 − 467. [12] ZHU Jiankang. Abiotic stress signaling and responses in plants [J]. Cell, 2016, 167(2): 313 − 324. [13] GUO Xiaoyu, LIU Dongfeng, CHONG Kang. Cold signaling in plants: insights into mechanisms and regulation [J]. Journal of Integrative Plant Biology, 2018, 60(9): 745 − 756. [14] WU H L, LI L, CHENG Z C, et al. Cloning and stress response analysis of the PeDREB2A and PeDREB1A genes in moso bamboo (Phyllostachys edulis) [J]. Genetics and Molecular Research, 2015, 14(3): 10206 − 10223. [15] THOMASHOW M F. So what’s new in the field of plant cold acclimation? Lots! [J]. Plant Physiology, 2001, 125(1): 89 − 93. [16] WANG Dazhi, JIN Ya’nan, DING Xihan, et al. Gene regulation and signal transduction in the ICE-CBF-COR signaling pathway during cold stress in plants [J]. Biochemistry, 2017, 82: 1103 − 1117. [17] SHI Yiting, DING Yanglin, YANG Shuhua. Molecular regulation of CBF signaling in cold acclimation [J]. Trends in Plant Science, 2018, 23(7): 623 − 637. [18] HWARARI D, GUAN Yuanlin, AHMAD B, et al. ICE-CBF-COR signaling cascade and its regulation in plants responding to cold stress [J/OL]. International Journal of Molecular Sciences, 2022, 23(3): 1549[2023-07-20]. doi: 10.3390/ijms23031549. [19] JIN Yanan, ZHAI Shanshan, WANG Wenjia, et al. Identification of genes from the ICE-CBF-COR pathway under cold stress in Aegilops-Triticum composite group and the evolution analysis with those from triticeae [J]. Physiology and Molecular Biology of Plants, 2018, 24(2): 211 − 229. [20] 陈露, 杨立明, 罗玉明. 植物ICE蛋白基因家族的系统进化分析 [J]. 江苏农业科学, 2016, 44(2): 42 − 47. CHEN Lu, YANG Liming, LUO Yuming. Phylogenetic analysis of the plant ICE protein gene family [J] Jiangsu Agricultural Sciences, 2016, 44(2): 42 − 47. [21] YANG Xiaoyan, WANG Rui, HU Qinglei, et al. DlICE1, a stress-responsive gene from Dimocarpus longan, enhances cold tolerance in transgenic Arabidopsis [J]. Plant Physiology and Biochemistry, 2019, 142: 490 − 499. [22] DUAN Yadong, HAN Jiaxin, GUO Baitao, et al. MbICE1 Confers drought and cold tolerance through up-regulating antioxidant capacity and stress-resistant genes in Arabidopsis thaliana [J/OL]. International Journal of Molecular Sciences, 2022, 23(24): 16072[2023-07-20]. doi: 10.3390/ijms232416072. [23] DENG Cuiyun, YE Haiyan, FAN Meng, et al. The rice transcription factors OsICE confer enhanced cold tolerance in transgenic Arabidopsis [J/OL]. Plant Signaling & Behavior, 2017, 12(5): e1316442[2023-07-20]. doi: 10.1080/15592324.2017.1316442. [24] CHANDER S, ALMEIDA D M, SERRA T S, et al. OsICE1 transcription factor improves photosynthetic performance and reduces grain losses in rice plants subjected to drought [J]. Environmental and Experimental Botany, 2018, 150: 88 − 98. [25] 兰智鑫, 侯丹, 吴蔼民, 等. 毛竹PeCIGRs基因的克隆及表达分析[J]. 浙江农林大学学报, 2023, 40(5): 982 − 990. LAN Zhixin, HOU Dan, WU Aimin, et al. Cloning and expression analysis of PeCIGRs gene in Phyllostachys edulis [J]. Journal of Zhejiang A&F University, 2023, 40(5): 982 − 990. [26] HUANG Bin, HUANG Zhinuo, MA Ruifang, et al. Genome-wide identification and expression analysis of LBD transcription factor genes in moso bamboo (Phyllostachys edulis) [J/OL]. BMC Plant Biology, 2021, 21(1): 34182934[2023-07-20]. doi: 10.1186/s12870-021-03078-3. [27] GAO Jian, ZHANG Ying, ZHANG Chunling, et al. Characterization of the floral transcriptome of moso bamboo (Phyllostachys edulis) at different flowering developmental stages by transcriptome sequencing and RNA-Seq analysis [J/OL]. PLoS One, 2014, 9(6): 24915141[2023-07-20]. doi: 10.1371/journal. pone. 0098910. [28] PENG Zhenhua, LU Ying, LI Lubin, et al. The draft genome of the fast-growing non-timber forest species moso bamboo (Phyllostachys heterocycla) [J]. Nature Genetics, 2013, 45(4): 456 − 461. [29] HUANG Zhuo, JIN Sihan, GUO Handu, et al. Genome-wide identification and characterization of TIFY family genes in moso bamboo (Phyllostachys edulis) and expression profiling analysis under dehydration and cold stresses [J/OL]. PeerJ, 2016, 4: e2620[2023-07-20]. doi: 10.7717/peerj.2620. [30] 徐秀荣, 杨克彬, 王思宁, 等. 毛竹bHLH转录因子的鉴定及其在干旱和盐胁迫条件下的表达分析[J]. 植物科学学报, 2019, 37(5): 610 − 620. XU Xiurong, YANG Kebin, WANG Sining, et al. Identification of bHLH transcription factors in moso bamboo (Phyllostachys edulis) and their expression analysis under drought and salt stress [J]. Plant Science Journal, 2019, 37(5): 610 − 620. [31] FINN R D, COGGILL P, EBERHARDT R Y, et al. The Pfam protein families database: towards a more sustainable future [J]. Nucleic Acids Research, 2016, 44(D1): 279 − 285. [32] 李新然, 张智俊, 喻珮瑶, 等. 毛竹SWEET基因家族的全基因组鉴定与分析[J]. 生物信息学, 2020, 18(4): 236 − 246. LI Xinran, ZHANG Zhijun, YU Peiyao, et al. Genome-wide identification and analysis of SWEET gene family in Phyllostachys edulis [J]. Bioinformatics, 2020, 18(4): 236 − 246. [33] LETUNIC I, DOERKS T, BORK P. SMART 7: recent updates to the protein domain annotation resource [J/OL]. Nucleic Acids Research, 2012, 40(D1): D302[2023-07-20]. doi: 10.1093/nar/gkr931. [34] CHEN Chengjie, CHEN Hao, ZHANG Yi, et al. TBtools: an integrative toolkit developed for interactive analyses of big biological data [J]. Molecular Plant, 2020, 13(8): 1194 − 1202. [35] LIU Hanmei, WANG Yongbin, LIU Lijun, et al. Pleiotropic ZmICE1 is an important transcriptional regulator of maize endosperm starch biosynthesis [J/OL]. Frontiers in Plant Science, 2022, 13: 895763[2023-07-20]. doi: 10.3389/fpls.2022.895763. [36] 蔡庆生. 植物生理学实验[M]. 北京: 中国农业大学出版社, 2013. CAI Qingsheng. Plant Physiology[M]. Beijing: China Agricultural University Press, 2013. [37] 邹琦. 植物生理学实验指导[M]. 北京: 中国农业出版社, 2000. ZHOU Qi. Guidance for Plant Physiology Experiments [M]. Beijing: China Agricultural University Press, 2000. [38] 吴林军. 非生物胁迫下毛竹qRT-PCR分析中内参基因的选择[D]. 杭州: 浙江农林大学, 2019. WU Linjun. The Selection of Endogenous Reference Genes in Phyllostachys edulis Treated with Abiotic Stresses for qRT-PCR Analysis [D]. Hangzhou: Zhejiang A&F University, 2019. [39] ZHOU L, HE Y J, LI J, et al. An eggplant SmICE1a gene encoding MYC-type ICE1-like transcription factor enhances freezing tolerance in transgenic Arabidopsis thaliana [J]. Plant Biology, 2020, 22(3): 450 − 458. [40] WANG Xipan, SONG Qiping, GUO Haoguo, et al. StICE1 enhances plant cold tolerance by directly upregulating StLTI6A expression [J]. Plant Cell Reports, 2022, 42(1): 197 − 210. [41] WANG Peiwen, ZHU Lin, LI Ziheng, et al. Genome-wide identification of the U-box E3 ubiquitin ligase gene family in cabbage (Brassica oleracea var. capitata) and its expression analysis in response to cold stress and pathogen infection [J/OL]. Plants-Basel, 2023, 12(7): 1437[2023-07-20]. doi: 10.3390/plants12071437. [42] MARINO D, DUNAND C, PUPPO A, et al. A burst of plant NADPH oxidases [J]. Trends in Plant Science, 2012, 17(1): 9 − 15. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20230445
 点击查看大图
点击查看大图
计量
- 文章访问数: 1599
- HTML全文浏览量: 563
- PDF下载量: 42
- 被引次数: 0
