




 下载:
下载:


-
香榧Torreya grandis ‘Merrillii’为红豆杉科Taxaceae榧属Torreya多年生常绿乔木,为第三纪孑遗植物,是中国特有的珍贵经济干果[1],主要分布在浙江、安徽、福建、湖南等省的丘陵山区。自2014年浙江省提出“香榧南扩”战略以来,香榧种植区已由原来的会稽山脉向浙西南延伸。截至2019年,浙江省香榧种植面积达5.39万 hm2,坚果产量多达6 000 t,产值达15亿元[2]。
目前,食用香榧仍是香榧产业经济效益的最主要来源[3]。香榧授粉至种子成熟跨越2个年度,历时17个月,因此,通常会出现同一株树上既有去年授粉、今年膨大成熟的果实(即“2代果”),也有今年受精形成的幼果[4]。种实生长发育经历了缓生期(头年5月初至当年4月底)、速生期(当年5月初至6月底)、种子内部充实期(当年6月底至9月上中旬)以及成熟期(白露至秋分之间的9月上中旬) 4个阶段。其中,3月上旬至7月上旬为香榧种实早期生长的关键时期,也是营养枝生长和花芽分化的重要时期,因此该阶段枝叶及种实内部的营养状况直接影响香榧种实膨大、成熟及品质[5]。研究发现:氮、磷、钙能够促进速生期香榧种实生长,钾能够提供种实膨大结束后的树体营养需求,种实采收后补充镁能够提高香榧的产量和质量[6]。另外,在澳洲坚果Macadamia integrifolia生长前期(3—6月),树体内的氮、钾主要用于开花、坐果及果实的生长发育;随着果实的成熟,氮、钾在叶片中大幅度升高,并逐渐用于营养生长[7]。薄壳山核桃Caraya illinoensis果实膨大期对氮、磷、钾的需求量较大[8]。香榧种实快速膨大过程一般从5月上旬开始,到6月下旬结束[5]。目前,对香榧栽培方面的研究较多,如土壤类型以及叶面施肥对香榧种实品质的影响等[9−10],但对香榧种实生长早期枝叶和种实中的矿质元素积累、分配的动态特征研究较少。本研究对香榧种实生长早期矿质元素的积累量和质量分数的动态变化进行研究和分析,旨在从营养供需的角度为提高香榧质量、产量和科学施肥提供理论依据。
-
试验地位于浙江省杭州市临安区浙江农林大学东湖校区香榧林。林地所在地属于中亚热带季风气候区南缘,温暖湿润,光照充足,雨量充沛,四季分明;年均降水量为1 613.9 mm,年均降水日为158.0 d,年均无霜期为237.0 d。
-
试验林中香榧树龄为(2+17) a,即砧木为2年生榧树,嫁接后生长了17 a,树体生长正常,并已进入结果期。林地每年施用复合肥2~3次,单株年施肥量平均为250~750 g·株−1·a−1,随树龄增大,逐年增加;有机肥隔年施1次,在树冠外围开20 cm深环状沟,单株施用腐熟羊粪10 kg·株−1。试验林0~20 cm土壤中碱解氮为71.470 mg·kg−1,全磷为1.216 g·kg−1,全钾为34.708 g·kg−1。
-
于试验林中随机选取10株香榧树,2019年3月上旬至7月上旬每隔15 d在香榧树体中部和东、南、西、北4个方向采集当年生侧枝及2代果实(即去年授粉,预计今年膨大成熟的果实),每株树采集10个种实和10个枝条,共10株树的果实混样和枝条混样,各分3组作为3个分析重复,共采样9次。根据生长发育阶段,3月10日至5月10日为缓生期;5月10日至6月25日为速生期;6月25日至7月10日为种实内部充实期。
-
将采集的样品先后用自来水、纯净水冲洗干净,再用滤纸干燥,待测。枝叶、种实分别用称量法称量,将枝条、种实置于烘箱中,105 ℃ 杀青 30 min,然后 60 ℃ 烘干至恒量,计算枝叶、种实样品的含水率,重复3次,取平均值。
-
将采集的枝叶及种实样品用自来水清洗干净后再用去离子水冲洗3次,将样品表面水分擦干后测量新鲜种实横径、纵径、质量。之后将香榧枝叶及种实置于烘箱中,经105 ℃杀青30 min后于60 ℃烘干至恒量,称干质量,即为香榧干物质积累量,再计算元素积累量[11]。单枝(单个种实)元素积累量=枝叶(种实)元素质量分数×干物质枝叶(种实)积累量[12]。
-
干燥后的香榧种实和枝叶样品经钢制粉碎机(DFT-50,温岭市林大机械有限公司)研磨后过60目尼龙筛。准确称取样品粉末0.1 g (精确至0.000 1 g)于玻璃消解管中,加入5 mL浓硝酸后加热至120 ℃保持45 min,冷却后向消解管加入体积分数为30%双氧水1 mL继续加热至消解液澄清。消解液冷却至室温后转移到50 mL容量瓶,用去离子水将消解管冲洗3次。冲洗液一并转移到容量瓶,去离子水定容后使用电感耦合等离子体-质谱仪(ICP-MS, Agilent 7700x)测定磷、钾、钙、镁、锌、锰、铜质量分数[13]。准确称取样品粉末0.1 g (精确至0.000 1 g),用元素分析仪(varo MACRO cube,elementar 公司)测定碳、氮、硫质量分数[14],计算种实和枝叶中矿质元素的积累量[15−16]。
-
采用SPSS 22.0进行单因素方差分析和相关分析,使用最小显著差异法(LSD,P<0.05)计算变异系数和回归系数,使用软件Excel 2007以及Chiplot (https://www.chiplot.online/)绘图。
-
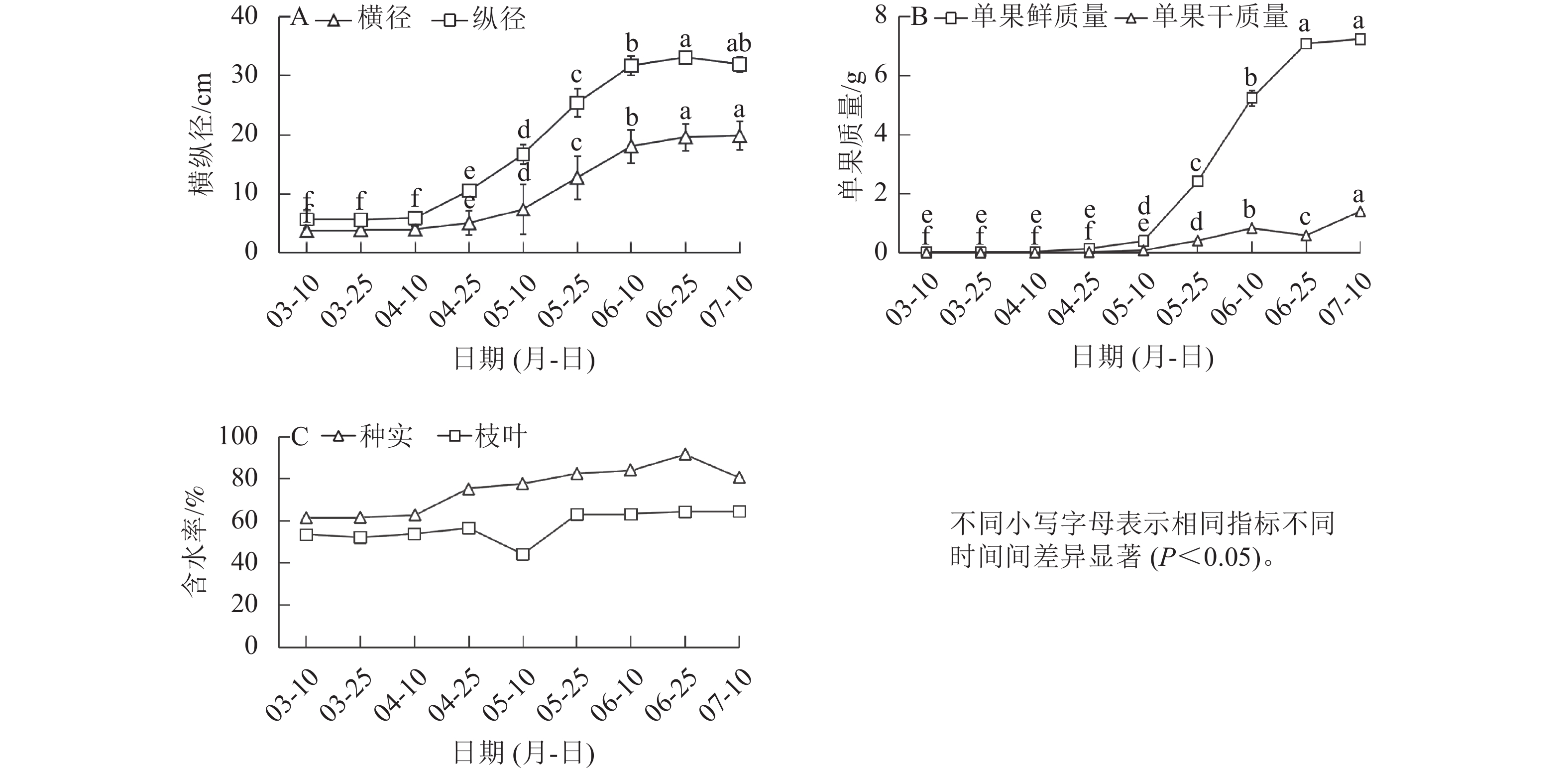
在香榧种实早期生长阶段,种实的横径与纵径在4月中旬前基本不变,4月中旬后显著增长(P<0.05),其中,纵径增幅较横径更大,且增长速度更快;到达6月中旬后,横径与纵径进入稳定期,不再增长(图1A)。与形态变化规律相似,单果的鲜质量和干质量在5月中旬前变化较小,5月中旬至6月底迅速增长,7月初进入平稳期(图1B)。4月上旬前枝叶、种实的含水率基本不变;4月中旬后枝叶含水率出现短暂下降后上升再趋于平稳,此时至6月下旬种实的含水率处于平稳上升状态(图1C)。
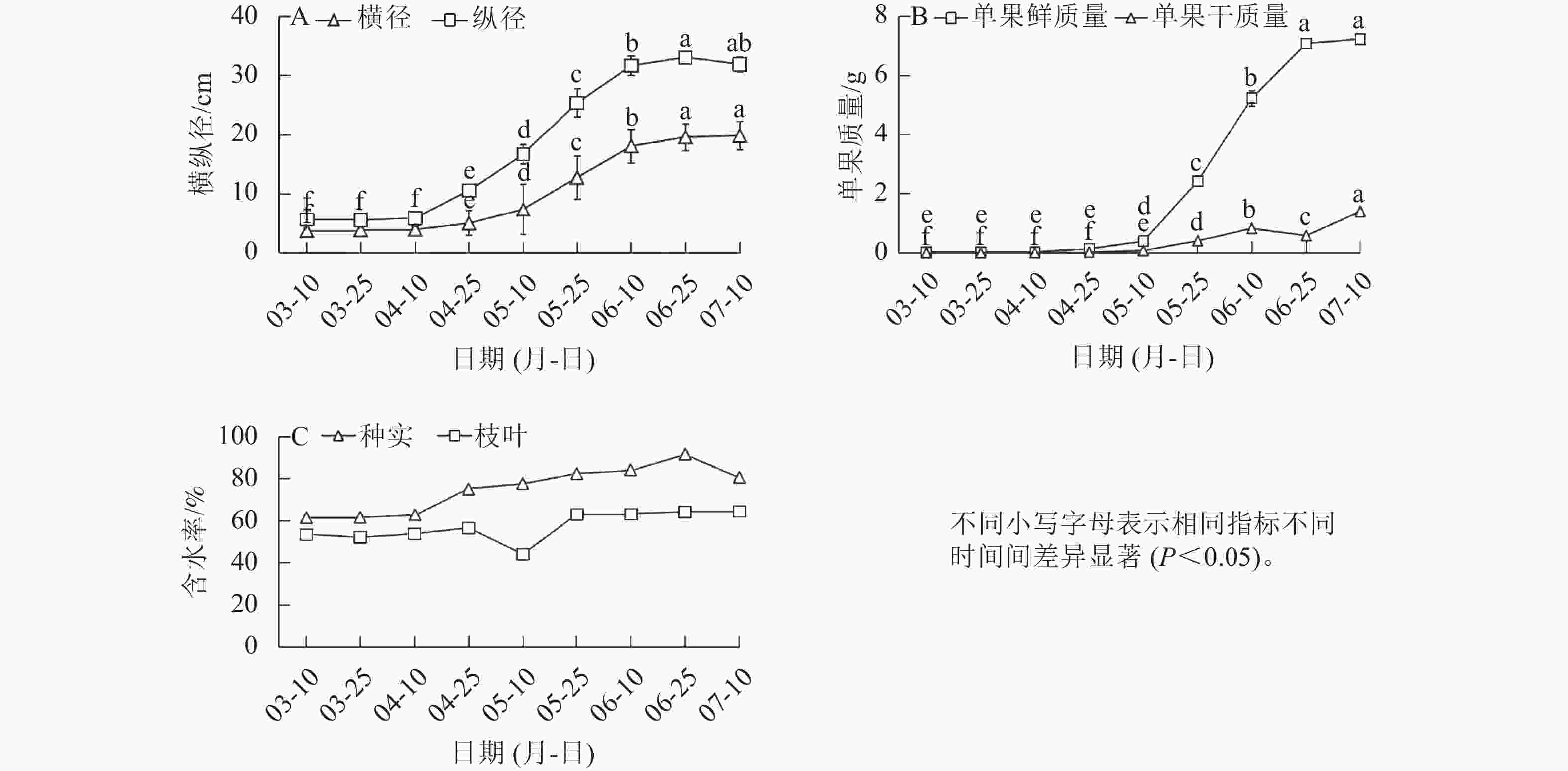
图 1 香榧种实横径与纵径、鲜质量与干质量
Figure 1. Transverse, longitudinal diameter, fresh weight, and dry weight of T. grandis ‘Merrillii’ seeds
-
在香榧种实早期生长阶段,枝叶中大量元素(氮、磷、钾)质量分数无显著差异,而在种实中则呈先上升后下降的趋势,其中,氮与磷质量分数均在4月下旬达到峰值,钾质量分数在6月中旬达到最高值(图2A~C)。枝叶中氮、磷、钾积累量在3月上旬至5月下旬之间较高,随后下降,而种实中氮、磷、钾积累量则在5月下旬后明显升高(图2D~F)。氮、磷、钾平均质量分数在枝叶中从大到小依次为氮、钾、磷,种实中依次为钾、氮、磷,且整个采样期种实中的氮、磷、钾质量分数均高于枝叶(图2A~C);枝叶与种实中氮、磷、钾元素积累量的动态曲线大致相反,氮、磷、钾的平均积累量在枝叶中从大到小依次为氮、钾、磷,而种实中顺序为钾、氮、磷。
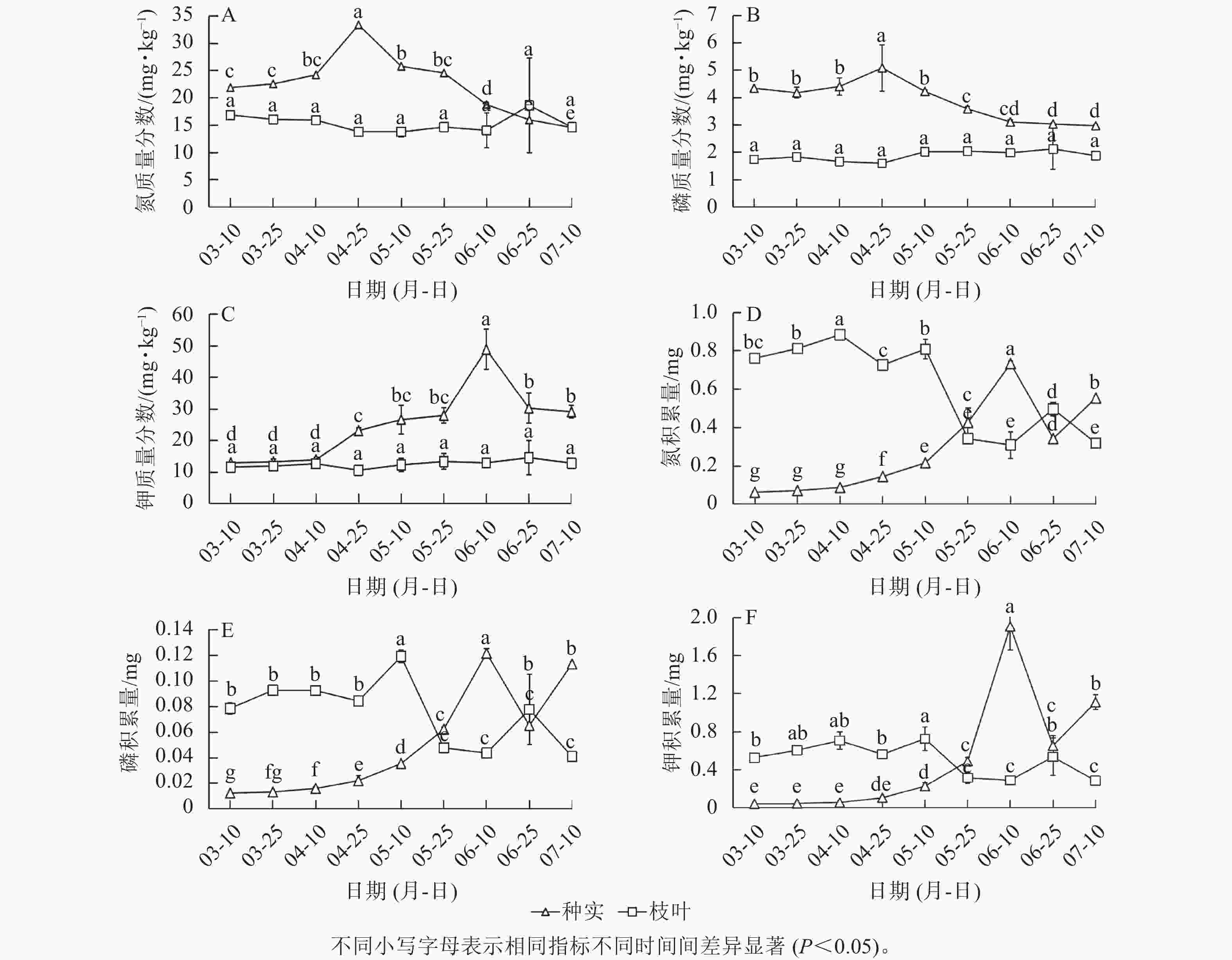
图 2 种实与枝叶氮、磷、钾质量分数及积累量
Figure 2. Contents and accumulation of N, P, K in seeds and leaves of T. grandis ‘Merrillii’
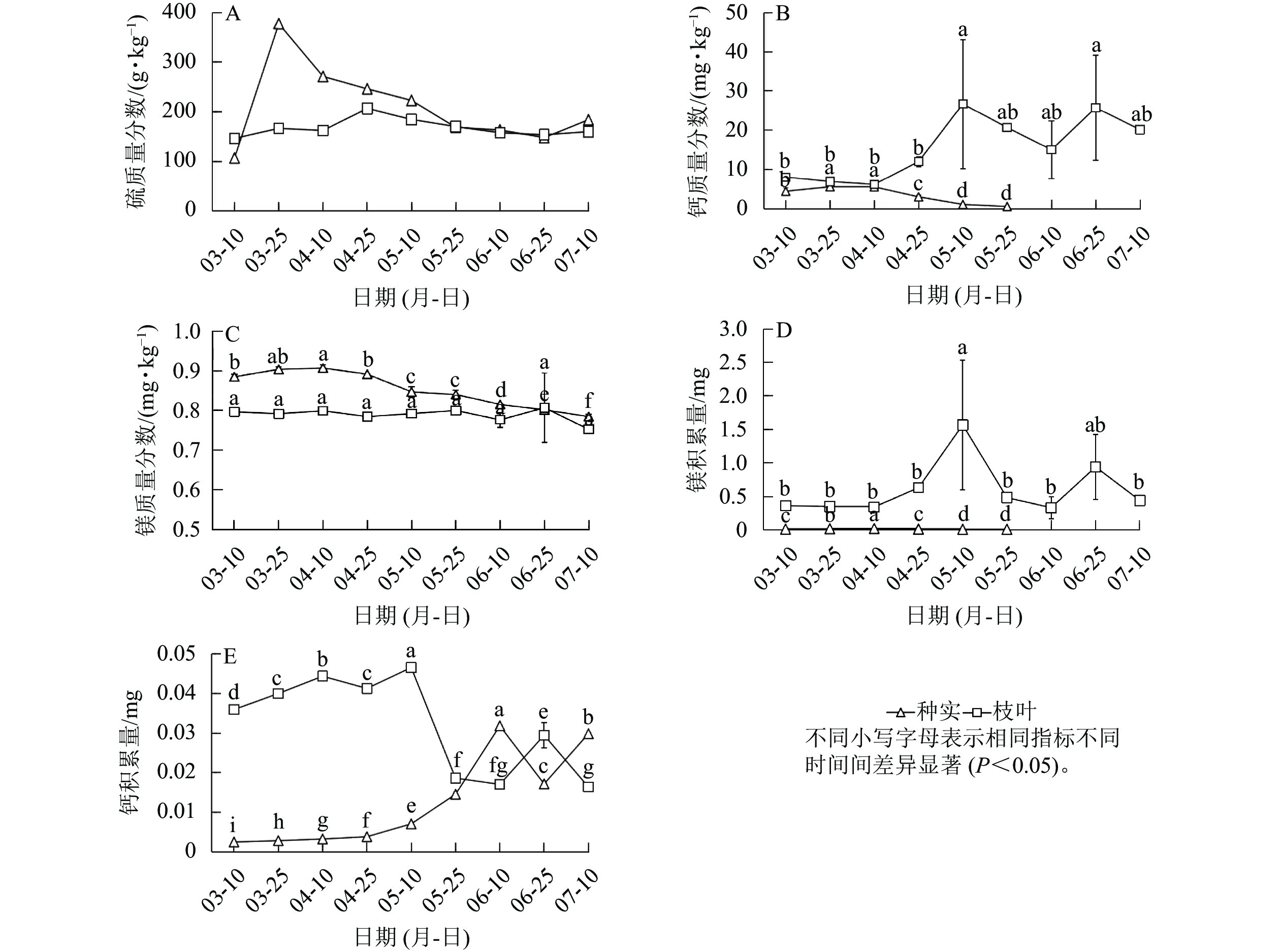
香榧种实早期生长阶段种实中的硫质量分数整体平稳,而枝叶中硫质量分数则呈先上升后下降的趋势(图3A)。枝叶中钙质量分数呈逐渐上升趋势,镁质量分数则较为稳定;种实中钙、镁质量分数均呈逐渐降低趋势(图3B~C)。此外,钙、镁积累量分析发现:随着种实早期生长,枝叶中钙积累量先上升后下降,镁积累量在5月中旬前较高,之后显著下降。种实中钙积累量总体很低,镁积累量则随着种实发育逐渐增加(图3D~E)。总体来看,钙质量分数和积累量枝叶均高于种实,镁质量分数枝叶也高于种实,但枝叶和种实镁积累量表现出相反趋势,5月底之前表现为枝叶高于种实,随后则为种实高于枝叶(图3E)。
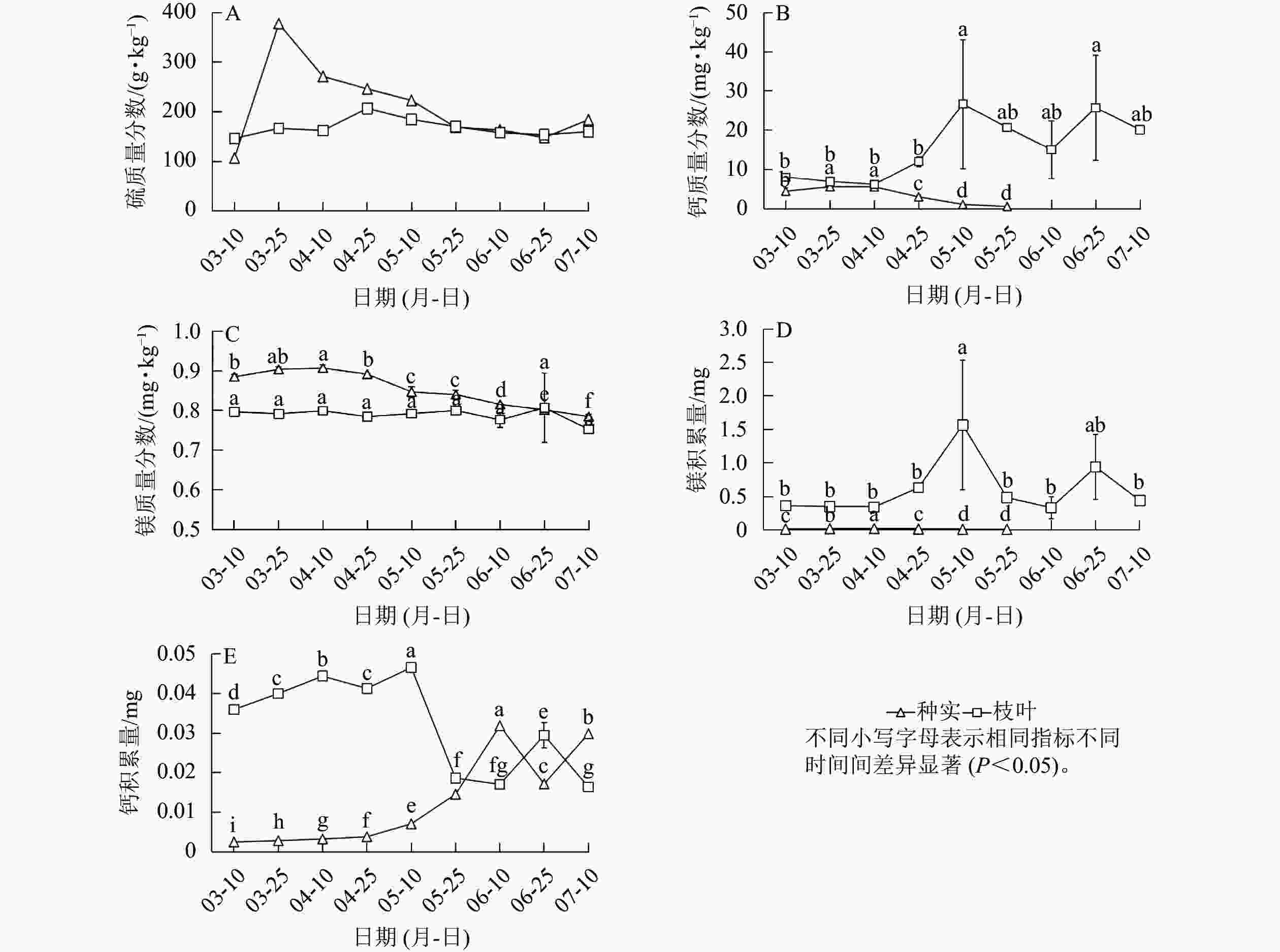
图 3 种实与枝叶硫、钙、镁质量分数和钙、镁积累量
Figure 3. S,Ca,Mg contents and Ca, Mg accumulation in seeds and branches and leaves of T. grandis ‘Merrillii’
-
随着香榧种实早期发育,枝叶中的锌和铜质量分数无显著变化,锰质量分数呈现先上升后下降趋势。种实中锌、铜质量分数均高于枝叶,且锌质量分数高于铜,但种实生长早期阶段锌、铜质量分数波动无明显规律,种实中锰质量分数总体也呈现先上升后下降趋势,但总体低于枝叶(图4A~C)。积累量分析发现:随着种实早期发育,枝叶中锌、铜、锰均在5月中旬前呈现较高积累量,而后逐渐降低,而种实中锌、铜、锰积累量与枝叶相反,在5月底之前积累量无明显变化,且远低于枝叶中的积累量,随后积累量逐渐增加,在6月中旬左右达到峰值(图4D~F)。
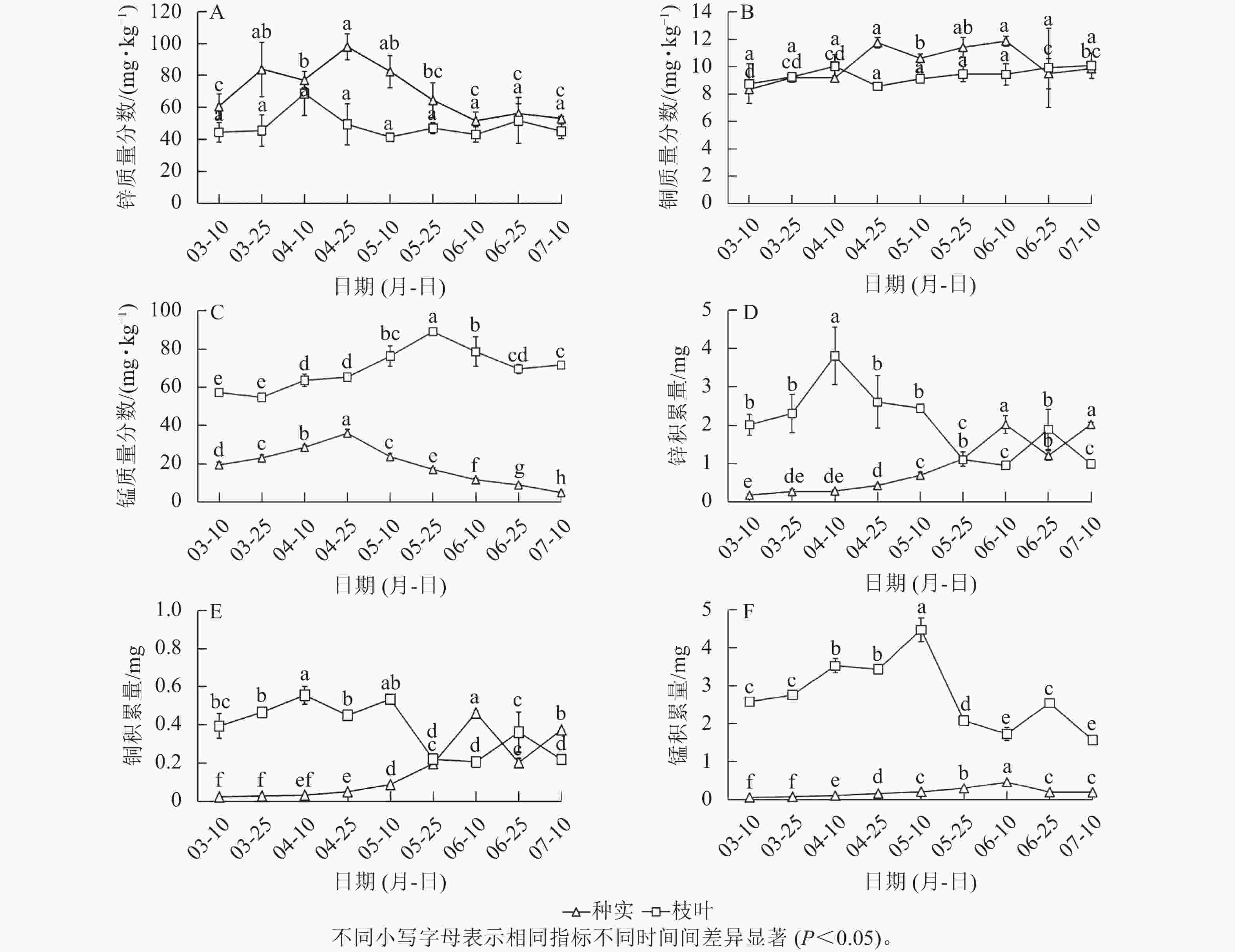
图 4 种实与枝叶锌、锰、铜质量分数和积累量变化
Figure 4. Contents and accumulation of Zn, Mn, Cu in seeds and leaves of T. grandis ‘Merrillii’
-
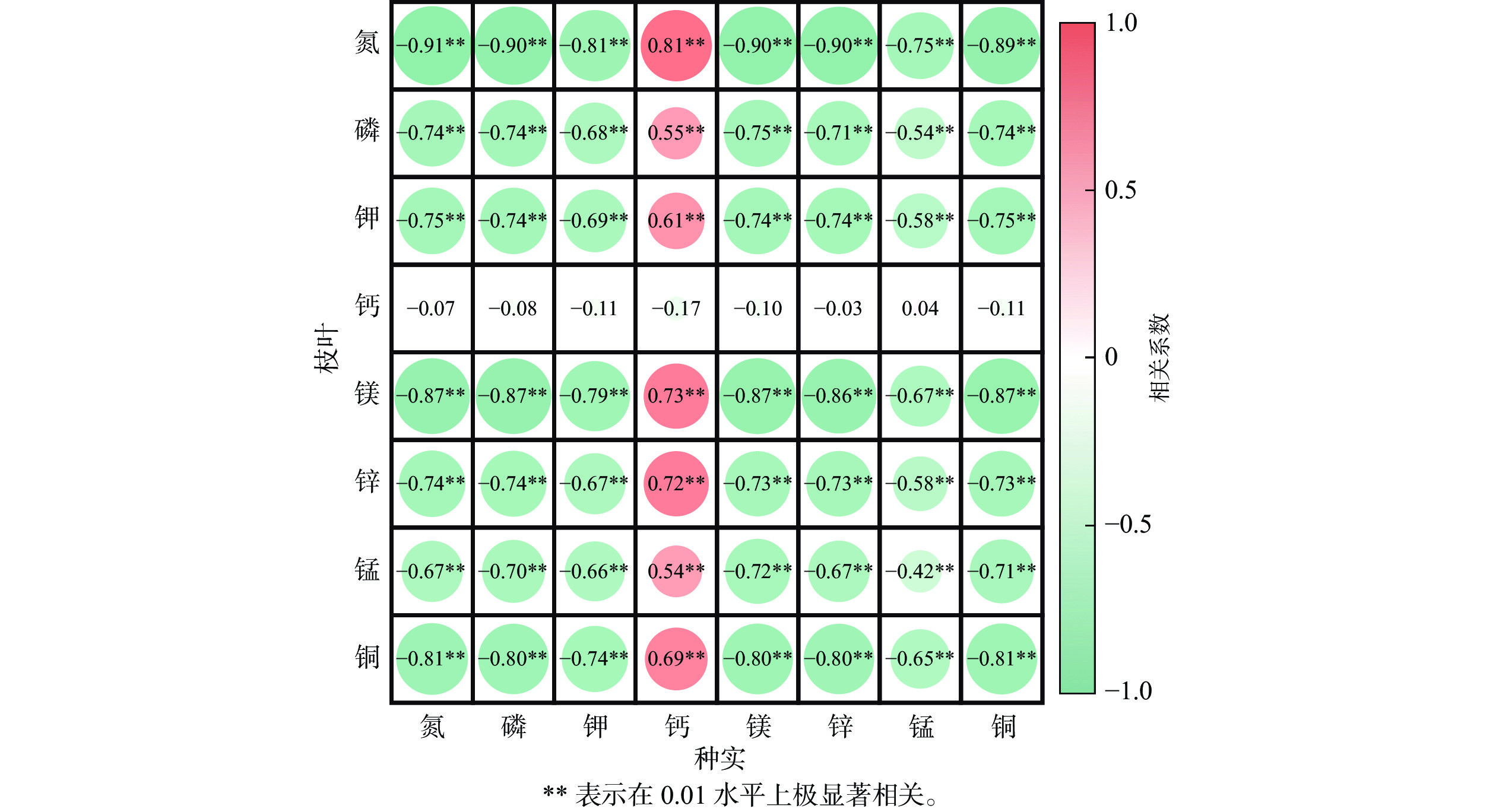
相关性分析(图5)发现:种实与枝叶中的氮、磷、钾、镁、锌、锰、铜积累量间存在显著负相关(P<0.05),而种实中的钙积累量与枝叶中上述元素积累量则呈显著正相关(P<0.05);枝叶中钙积累量则与种实中的各种元素积累量均无显著相关性(图5)。
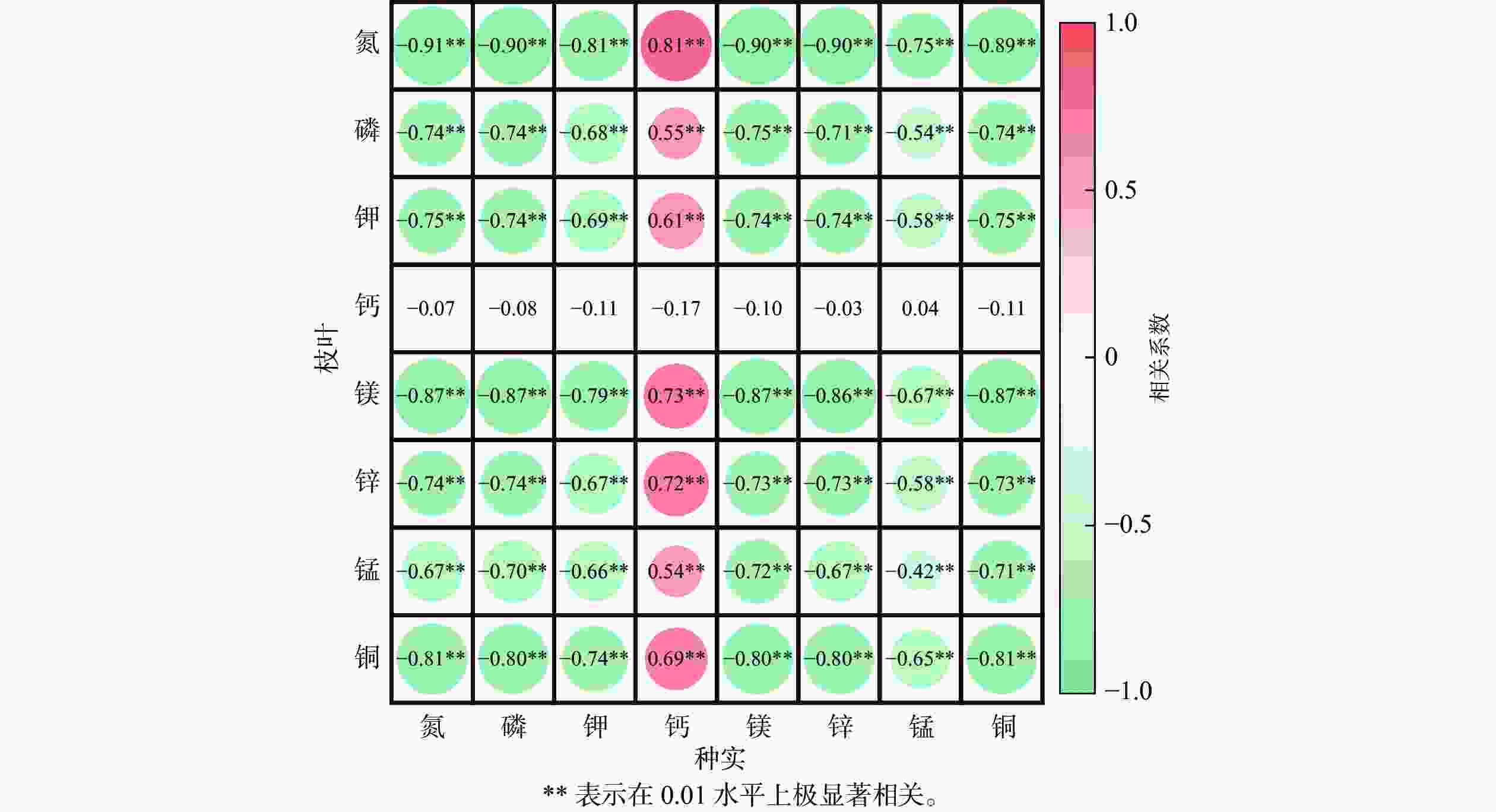
图 5 种实与枝叶元素积累量相关性分析
Figure 5. Correlation analysis of elemental accumulation in seeds and leaves of T. grandis ‘Merrillii’
-
种实是香榧产生经济价值最重要的来源。本研究发现:4月下旬至5月下旬香榧种实的横纵径显著增加,表明其进入快速膨大期。随着香榧种实膨大,体积也迅速增长,含水率显著增加,枝叶中出现短暂的含水率下降现象,可能与种实迅速膨大所需水分急剧上升有关,因此,种实鲜质量增幅大于干质量,这与张晓兰等[17]的研究结果类似。4月中旬为香榧种实早期生长阶段,水分是带动香榧营养元素转移和转化的重要载体。
植物中矿质元素质量分数变化揭示了植物与环境之间的相互关系[18]。植物矿质元素积累量是植物生产力以营养物质形式的表现,其大小由植物的生物量和组织中元素质量分数共同决定[19]。本研究发现:4月上旬前处于缓生期的香榧种实对氮、磷、钾的需求量较小,4月中下旬随着种实进入速生期,对氮、磷、钾的需求逐渐增加。此时,枝叶中的氮、磷、钾优先供应种实生长,导致新梢枝叶中的氮、磷、钾积累量下降,尤其是5—6月种实体积生长旺盛期叶片中氮、磷、钾积累量下降极为显著。5月后种实氮、磷质量分数由上升转为下降,但积累量仍在继续上升,因此氮、磷质量分数的下降可能是由于生长旺盛期种实含水率上升造成的。种实中的钾质量分数在4—5月速生期显著持续积累,可见种实生长发育对钾的需求巨大。从整体水平来看,氮、磷、钾3种元素在种实中的积累量在6月下旬开始下降,因此,为了满足种实生长发育需求,生产上可在速生期开始之前施加氮、磷、钾肥。6月中下旬以后,种实由速生期转入内部充实期,生理活动逐渐变缓,转为种仁发育和内部物质积累[5]。这一阶段伴随种实落果,减少了对养分的需求,留存的种实间竞争减少,营养元素的积累量进一步上升。钙是植物生长发育所必需的矿质元素和重要调节因子[20],钙在韧皮部中较难移动,蒸腾作用带动的水流速率是影响钙离子沿导管运输的关键动力[21−22]。本研究表明:种实早期生长过程中,钙质量分数和积累量均逐渐降低,这主要是因为速生期种实含水率的提高稀释了钙元素。此外,3—7月正值营养枝旺盛生长期,枝叶蒸腾作用加剧了叶片对钙的竞争,因此,为了促进香榧种实的膨大,提升品质,应适当增施钙肥[21, 23]。镁是叶绿素的重要组成成分,在植物生长过程中起着不可或缺的作用[24−25]。本研究中,随着香榧种实早期生长,种实中镁质量分数高于枝叶,但枝叶和种实中镁质量分数都随生长呈下降趋势。进入到速生期后,镁元素优先供应种实的生长需求,因此,果实生长期尤其是成熟期镁肥的施用可能会导致果实品质的下降[26−27]。生产上应在种实成熟采摘后施用足量镁肥来提高次年坐花坐果率和果实品质,在3—7月无需施加镁肥。
此外,本研究发现:香榧种实在缓生期与花芽分化期对锌和铜元素的需求量不高,而进入速生期后对锌和铜的需求增加。因此,可以在速生期适当施加含锌、铜等微量元素肥料,提高果实品质。同时,研究也发现:种实对锰需求量小,主要积累在叶片中,可见树体本身及环境供给已经能够满足种实对锰的需求,因此不需要额外施加。但需要注意,香榧花芽分化受到组织中氮、锌、锰质量分数的影响,过量的氮、锌、锰积累会抑制混合芽数量的增加[23],因此施加微量元素肥料要注意树体本身营养情况,不宜过多。本研究还发现:在7月初种实中营养元素(除钙元素外)的积累量都有一定程度的下降。这可能是由于2代果发育快成熟时,为了让当年授粉成功的果实更好发育,树体本身调节2代果中的一部分营养元素至枝叶中,以便于枝叶供应和转移营养用于当年幼果的发育,因此在前期积累足够的营养物质,不但有利于2代果的成熟和质量提高,也有利于当年幼果的生长。
相关性分析结果也表明:枝叶和种实在氮、磷、钾、镁、锌、锰、铜等营养元素的积累上存在一定的竞争关系,枝叶生长过度旺盛会导致枝叶对营养的竞争力增强,不利于种实的生长;果实中的钙积累量与枝叶中其他元素(除钙以外)正相关关系说明,由于枝叶蒸腾作用对钙竞争力强,当果实中钙的积累量上升时,枝叶已经积累了足够的钙,有利于枝叶对其他元素的吸收,可见施加足量的钙元素才能满足种实生长需求。因此,在种实生长发育期开始之前,不应过度施加氮、磷、钾、镁、锌、锰、铜等营养元素,可结合适当修枝等操作,减少枝叶对养分的竞争;种实生长发育期早期和速生期则需要适当施加钙肥以提高枝叶中的钙元素储量,保障种实的快速生长所需。
综上所述,为有效促进香榧种实生长发育,须在种实缓生期末期和速生期开始时施加氮、磷、钾肥,以供应种实快速生长和充实期对氮、磷、钾元素的大量需求;速生期应适当施加钙肥,促进香榧种实膨大结实,对提高香榧种实品质有重要意义。但是,由于镁肥可能会影响钙肥的吸收,因此建议在种实采收后施用镁肥。同时,适量施用含有锌、铜的微量元素肥料可以有效地协调枝叶与种实中的营养平衡,优化香榧树体营养吸收,从而改善因单一施肥造成产量减少、品质下降的现象,促进香榧产业可持续发展。
-
本研究主要得出以下结论:①香榧种实的体积和含水率在速生期快速增加,进入充实期后趋于稳定。②元素质量分数、积累量与种实早期生长期密切相关,从缓生期到充实期8种元素质量分数与积累量都呈现先升后降的趋势,其中速生期对钾元素需求量最大。为了提高种实品质,应在种实缓生期末期和速生期开始时施加氮、磷、钾肥,并配合适量施用锌、铜等微量元素肥料,在种实采收后施用镁肥,香榧种实早期生长阶段适当增施钙肥。
Dynamic analysis of content and accumulation of mineral elements in branch-leaf mixture and seeds of Torreya grandis ‘Merrillii’ during the early growth stage
-
摘要:
目的 探究香榧Torreya grandis ‘Merrillii’种实早期生长阶段枝叶与种实矿质元素质量分数、积累量的关系,为科学施肥,提升香榧种仁品质及产量提供依据。 方法 选择浙江农林大学东湖校区(2+17) a香榧试验林(砧木为2年生,嫁接后生长了17 a),采集3月上旬至7月上旬当年生侧枝(含叶片)及2代种实,对枝叶、种实中氮、磷、钾、钙、镁、锌、锰、铜质量分数及积累量动态变化关系进行分析。 结果 香榧种实早期生长阶段,种实横纵径、单果干鲜质量及含水率在4月上旬前无明显变化,4月中旬至6月中旬均增长迅速,之后趋于平稳。4月中下旬至6月中旬,枝叶中氮、磷、钾的积累量下降,而种实中氮、磷积累量及钾质量分数持续上升,表明枝叶中的氮、磷、钾优先供应种实的生长,缓生期与花芽分化期种实对氮、磷、钾的需求量较小,速生期种实对氮、磷、钾需求增加,该现象随时间推移而增强;在时间尺度上,钙在枝叶中逐渐吸收、积累,种实中钙质量分数及积累量则呈显著、极显著降低;速生期后,种实中的镁积累量增高,枝叶中镁积累量及质量分数均随着种实的膨大而下降;香榧种实早期生长阶段枝叶中锌、铜积累量呈下降趋势,种实对锌、铜、锰需求量较小。 结论 ①速生期香榧种实的体积和含水率快速增加,进入充实期后趋于稳定。②元素质量分数、积累量与香榧种实生长期密切相关。为提高香榧种实品质,应在种实缓生期末期和速生期开始时施加氮、磷、钾肥,并配合施用适量锌、铜等微量元素肥料,在种实采收后施用镁肥。图5参27 Abstract:Objective The objective is to explore the relationship between the content and accumulation of mineral elements in the branch-leaf mixture and seeds of Torreya grandis ‘Merrillii’ during the early growth stage of seeds, in order to provide a basis for scientific fertilization and improve the quality and yield of T. grandis ‘Merrillii’ seeds. Method T. grandis ‘Merrillii’ trees (2 a rootstocks and 17 a after grafting) were selected for experiment on Donghu campus of Zhejiang A&F University. Lateral branches (with leaves) and seeds of the second generation were collected from early March to early July, and the dynamic changes in nitrogen (N), phosphorus (P), potassium (K), calcium (Ca), magnesium (Mg), zinc (Zn), manganese (Mn) and copper (Cu) content and accumulation in branch-leaf mixture and seeds were analyzed. Result In the early growth stage of T. grandis ‘Merrillii’ seeds, there was no significant change in the transverse and longitudinal diameter of the seeds, dry and fresh weight, and moisture content of each fruit before early April. However, from mid April to mid June, the growth rate increased rapidly and then stabilized, and the accumulation of N, P, and K in branch-leaf mixture decreased while the accumulation and content of N, P, and K in seeds continued to increase, which indicated that N, P, and K in branch-leaf mixture were preferentially supplied to the growth of seeds. During the slow growth and flower bud differentiation stage, the demand for N, P, and K by seeds was relatively small, while during the rapid growth stage, the demand for N, P, and K by seeds increased. This phenomenon intensified with time. On a time scale, Ca gradually absorbed and accumulated in the branch-leaf mixture, while Ca content and accumulation in seeds decreased and ertremely decreased significantly. After the rapid growth period, the accumulation of Mg in seeds increased, while the accumulation and content of Mg in branch-leaf mixture decreased with the expansion of seeds. During the early growth stage of T. grandis ‘Merrillii’ seeds, the accumulation of Zn and Cu in branch-leaf mixture showed a decreasing trend, and the seed had a relatively small demand for Zn, Cu and Mn. Conclusion (1) The volume and moisture content of the fast-growing T. grandis ‘Merrillii’ seeds increase rapidly, and tend to stabilize after the filling period. (2) The content and accumulation of elements are closely related to the long-term growth of T. grandis ‘Merrillii’ seeds. To improve the quality of seeds, N, P and K fertilizers should be applied at the end of the slow growth period and the beginning of the fast growth period, along with an appropriate amount of trace element fertilizers such as Zn and Cu. Mg fertilizer should be applied after seed harvest. [Ch, 5 fig. 27 ref.] -
图 1 香榧种实横径与纵径、鲜质量与干质量
Figure 1 Transverse, longitudinal diameter, fresh weight, and dry weight of T. grandis ‘Merrillii’ seeds
图 2 种实与枝叶氮、磷、钾质量分数及积累量
Figure 2 Contents and accumulation of N, P, K in seeds and leaves of T. grandis ‘Merrillii’
图 3 种实与枝叶硫、钙、镁质量分数和钙、镁积累量
Figure 3 S,Ca,Mg contents and Ca, Mg accumulation in seeds and branches and leaves of T. grandis ‘Merrillii’
图 4 种实与枝叶锌、锰、铜质量分数和积累量变化
Figure 4 Contents and accumulation of Zn, Mn, Cu in seeds and leaves of T. grandis ‘Merrillii’
-
[1] 刘琏, 陆吕佳, 胡渊渊, 等. 不同月份不同树龄香榧叶片光合特性的变化[J]. 浙江农林大学学报, 2022, 39(1): 32 − 40. LIU Lian, LU Lüjia, HU Yuanyuan, et al. Changes of photosynthesis in leaves of Torreya grandis ‘Merrillii’ in different months and different tree ages [J]. Journal of Zhejiang A&F University, 2022, 39(1): 32 − 40. [2] 喻卫武. 香榧生态高效栽培技术[J]. 浙江林业, 2020(10): 22. YU Weiwu. High efficient ecological cultivation techniques of Torreya grandis ‘Merrillii’ [J]. Zhejiang Forestry, 2020(10): 22. [3] 徐翠霞. 浙江省香榧产业发展及其对策研究[D]. 杭州: 浙江农林大学, 2019. XU Cuixia. Study on Torreya grandis ‘Merrillii’ Production Development and Its Suggestions in Zhejiang Province [D]. Hangzhou: Zhejiang A&F University, 2019 [4] 胡绍泉, 张益锋. 香榧的生态生物学特征及其播种培育技术[J]. 绍兴文理学院学报(自然科学), 2015, 35(7): 28 − 31. HU Shaoquan, ZHANG Yifeng. The ecological and biological characteristics and sowing cultivation of Torreya grandis [J]. Journal of Shaoxing University, 2015, 35(7): 28 − 31. [5] 黎章矩, 戴文圣. 中国香榧[M]. 北京: 科学出版社, 2007. LI Zhangju, DAI Wensheng. Torreya grandis ‘Merrillii’ of China [M]. Beijing: Science Press, 2007. [6] 刘萌萌, 曾燕如, 江建斌, 等. 香榧叶片中8种矿质元素年周期季节性变化规律[J]. 经济林研究, 2014, 32(2): 105 − 109. LIU Mengmeng, ZENG Yanru, JIANG Jianbin, et al. Seasonal dynamic changes of eight mineral elements in Torreya grandis leaf [J]. Nonwood Forest Research, 2014, 32(2): 105 − 109. [7] 韩树全, 范建新, 王代谷, 等. 澳洲坚果生育期内叶片矿质营养元素含量及其变化[J]. 安徽农业科学, 2016, 44(23): 8 − 10, 52. HAN Shuquan, FAN Jianxin, WANG Daigu, et al. Study on mineral elements content and change in Macadamia ternifolia leaves during the development period [J]. Journal of Anhui Agricultural Sciences, 2016, 44(23): 8 − 10, 52. [8] 袁紫倩, 杨先裕, 凌骅, 等. 薄壳山核桃‘马汉’叶片主要矿质营养元素生育期动态变化特征[J]. 西北植物学报, 2016, 44(23): 8 − 10. YUAN Ziqian, YANG Xianyu, LING Hua, et al. Dynamic changes of leaves’ main mineral elements in pecan ‘Mahan’s growth period [J]. Acta Botanica Boreali-Occidentalia Sinica, 2016, 44(23): 8 − 10. [9] 曹永庆, 任华东, 王开良, 等. 不同类型土壤栽培香榧种仁品质综合评价和分析[J]. 果树学报, 2022, 39(5): 836 − 845. CAO Yongqing, REN Huadong, WANG Kailiang, et al. Comprehensive evaluation and analysis of kernel quality of Torreya grandis ‘Merrillii’ from different soil types [J]. Journal of Fruit Science, 2022, 39(5): 836 − 845. [10] 郝琪淳, 谢吉全, 戴文圣, 等. 香榧种实充实期叶面施肥对种实品质的影响[J]. 浙江农林大学学报, 2024, 41(3): 457 − 466. HAO Qichun, XIE Jiquan, DAI Wensheng, et al. Effect of foliar fertilization on seed quality of Torreya grandis ‘Merrillii’ during seed filling period [J]. Journal of Zhejiang A&F University, 2024, 41(3): 457 − 466. [11] 何斌, 黄恒川, 曹明, 等. 秃杉人工林微量元素含量、积累与分配特征[J]. 南京林业大学学报(自然科学版), 2009, 33(1): 69 − 73. HE Bin, HUANG Hengchuan, CAO Ming, et al. Concentration, accumulation and distribution of microelementsin Tawania flousiana plantation [J]. Journal of Nanjing Forestry University (Natural Sciences Edition), 2009, 33(1): 69 − 73. [12] 盖甜甜, 邹中华, 任志华, 等. 不同氮养分条件下油茶幼苗生物量及养分利用对磷水平的响应[J]. 江西农业大学学报, 2022, 44(3): 626 − 634. GAI Tiantian, ZOU Zhonghua, REN Zhihua, et al. Effects of biomass and nutrient utilization of Camellia oleifera seedlings to phosphorus level under different nitrogen nutrients [J]. Acta Agriculturae Universitatis Jiangxiensis, 2022, 44(3): 626 − 634. [13] LI Xiaogang, CAO Yini, XIAO Jiang, et al. Bamboo biochar greater enhanced Cd/Zn accumulation in Salix psammophila under non-flooded soil compared with flooded [J/OL]. Biochar, 2022, 4: 7[2023-11-04]. doi: 10.1007/s42773-022-00139-0. [14] 刘凡, 马宗虎, 张廷军, 等. 元素分析仪测定玉米秸秆元素含量不确定度评定[J]. 中国沼气, 2021, 39(4): 25 − 28. LIU Fan, MA Zonghu, ZHANG Tingjun, et al. Evaluation of the uncertainty of element content determination of corn stalks by elemental analyzer [J]. China Biogas, 2021, 39(4): 25 − 28. [15] 葛高波, 高智群, 刘欢, 等. 黄竹地上部分营养元素的吸收、积累和分配特性[J]. 竹子研究汇刊, 2016, 35(1): 41 − 45. GE Gaobo, GAO Zhiqun, LIU Huan, et al. Uptake, accumulation and distribution of main elemental nutrients in the aboveground parts of Dendrocalamus membranceus [J]. Journal of Bamboo Research, 2016, 35(1): 41 − 45. [16] 叶淑媛. 香榧种实早期生长与树体营养的相关性研究[D]. 杭州: 浙江农林大学, 2020. YE Shuyuan. A Study on Early Growth of Seeds and Its Correlation to Tree Nutrition in Torreya grandis ‘Merrillii’ [D]. Hangzhou: Zhejiang A&F University, 2020. [17] 张晓兰, 肖龙洋. 香榧果实生长发育规律观测[J]. 湖南林业科技, 2011, 38(3): 36 − 38. ZHANG Xiaolan, XIAO Longyang. Growth and development rhythm of Torreya grandis fruit [J]. Hunan Forestry Science &Technology, 2011, 38(3): 36 − 38. [18] 王冕之, 张晓蓉, 牟凌, 等. 亚热带常绿阔叶林优势植物4种矿质元素的分配特征[J]. 应用与环境生物学报, 2020, 26(1): 40 − 47. WANG Mianzhi, ZHANG Xiaorong, MOU Ling, et al. Allocation of four mineral elements in dominant plants of subtropical evergreen broad-leavedforest [J]. Chinese Journal of Applied and Environmental, 2020, 26(1): 40 − 47. [19] 王彬, 魏天兴, 刘钊. 黄土丘陵区华北落叶松人工林生态系统生物量与养分循环特征[J]. 水土保持研究, 2017, 24(6): 45 − 51. WANG Bin, WEI Tianxing, LIU Zhao, et al. Characteristics of nutrient cycling and ecosystem structure of Larix principis-rupprechtii in Hilly Loess Plateau [J]. Research of Soil and Water Conservation, 2017, 24(6): 45 − 51. [20] ISLAM M Z, MELE M A, BAEK J P, et al. Cherry tomato qualities affected by foliar spraying with boron and calcium [J]. Horticulture Environment and Biotechnology, 2016, 57(1): 46 − 52. [21] MONTANARO G, DICHIO B, XILOYANNIS C. Significance of fruit transpiration on calcium nutrition in developing apricot fruit [J]. Journal of Plant Nutrition And Soil Science, 2010, 173(4): 618 − 622. [22] WHITE P J, BROADLEY M R. Calcium in plants [J]. Annals of Botany, 2003, 92(4): 487 − 511. [23] 叶淑媛, 曾燕如, 胡渊渊, 等. 香榧初结果母枝性状变化规律与结实能力的关系[J]. 浙江农林大学学报, 2022, 39(1): 41 − 49. YE Shuyuan, ZENG Yanru, HU Yuanyuan, et al. Relationship between character changing and seed-bearing capacity of initial seed-bearing mother shoots in Torreya grandis ‘Merrillii’ [J]. Journal of Zhejiang A&F University, 2022, 39(1): 41 − 49. [24] GUO Wanli, NAZIM H, LIANG Zongsuo, et al. Magnesium deficiency in plants: an urgent problem [J]. The Crop Journal, 2016, 4(2): 83 − 91. [25] 刘萌萌, 曾燕如, 江建斌, 等. 香榧生长期叶片和种子中矿质元素动态变化研究[J]. 浙江农林大学学报, 2014, 31(5): 724 − 729. LIU Mengmeng, ZENG Yanru, JIANG Jianbin, et al. Mineral elements in leaves and seeds of Torreya grandis ‘Merrilli’ during seed development [J]. Journal of Zhejiang A&F University, 2014, 31(5): 724 − 729. [26] 吕玉兰, 黄家雄, 王跃全. 镁肥对台湾青枣叶片叶绿素含量和果实品质的影响[J]. 热带农业科学, 2010, 30(12): 17 − 19. LÜ Yulan, HUANG Jiaxiong, WANG Yuequan, et al. Effects of magnesium fertilizer on chlorophyll content of leaves and fruit quality of ber (Zizvphus mauritiana Lam. ) [J]. Chinese Journal of Tropical Agriculture, 2010, 30(12): 17 − 19. [27] NEDIM O, DAMLA B O. Effect of magnesium fertilization on some plant nutrient interactions and nut quality properties in Turkish hazelnut (Corylus avellana L. ) [J]. Scientific Research and Essays, 2015, 10(14): 465 − 470. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20230583
 点击查看大图
点击查看大图
计量
- 文章访问数: 1832
- HTML全文浏览量: 569
- PDF下载量: 33
- 被引次数: 0
