








-
近年来,随着园林布景、环境绿化要求的日益提高,彩色针叶树的需求量也随之增大。彩色针叶树株型丰富多姿,叶色明亮多彩,观赏期长,是家庭、城市、园林及道路环境绿化的优良植物材料,尤其为北方园林增添了丰富的景观色彩[1-2]。因此,引进彩色针叶树种,研究并提高其引种适应性、生长繁殖能力和观赏价值是科研和园艺工作的热点方向。目前,国内外关于彩色针叶树的研究主要集中在引种适应性[3]、扦插繁殖[4-5]、生长生理[6-7]以及光合特性[8]等方面,对观赏性状如叶色变化的研究甚少。对彩色阔叶树研究发现:许多树种叶色变化与其色素含量显著相关,建立两者的回归方程能够通过叶色参数准确预测色素含量的变化,为实现人工调控彩色植物叶色变化、选育优良观赏植物品种提供了理论基础和技术支撑[9-14]。欧洲刺柏‘金叶疏枝’Juniperus communis ‘Gold Cone’和鹿角桧‘金叶’Juniperus×media ‘Pfitzeriana Aurea’是引种自比利时的彩色针叶树种,作为彩色针叶树中的金叶植物代表,不同季节均有很高的观赏价值,弥补了园林景观及环境绿化色彩单调的缺憾[15]。本研究以欧洲刺柏‘金叶疏枝’和鹿角桧‘金叶’为研究对象,测定不同季节2种树的叶色参数和色素变化,比较两者观赏性状差异;分析叶色参数与色素的相关关系,建立回归模型。本研究将为‘金叶疏枝’和‘金叶’的叶色变化生理机制研究提供基础,也为叶色调控技术研发提供理论依据。
HTML
-
欧洲刺柏‘金叶疏枝’和鹿角桧‘金叶’来自中国林业科学研究院华北林业实验中心,均为2年生扦插苗。
-
选择生长情况相近的的2年生‘金叶疏枝’和‘金叶’各180株,分别分成3个重复;于2019年春季、夏季、秋季和冬季,随机采集各重复内1~2株扦插苗(整株),用塑封袋封好,放入冰盒,立即带回实验室测定其叶片的叶色参数、光合色素(叶绿素a、叶绿素b、类胡萝卜素)质量分数和花色素苷相对含量。
-
参照1970年国际照明委员会推出的CIELab表色系统,以明亮度(L*)、色相(a*)、色相(b*)表示色彩。L*值从0升至100,表示亮度逐渐增加,a*值由小变大,表示由绿色逐渐变为红色,b*值由小到大,表示由蓝色逐渐变成黄色。参照黄可等[11]的方法,采用EPSON扫描仪(日本) 扫描2种刺柏属彩色树扦插苗鲜叶,运用Adobe Photoshop(CS6,Adobe,美国)软件的拾色功能,记录叶尖、中心点、叶柄基部等8个点的L*、a*、b*的值,取平均值作为叶色参数。
-
叶绿素上清液的制备参照王学奎等[16]和陈明等[17]的方法,并略有改动。以体积分数95%的乙醇为对照,采用分光光度计分别在波长665、649和470 nm测定上清液吸光度,分别计算光合色素的质量分数。叶绿素a质量浓度(mg·L−1):Ca=13.95D(665)−6.88D(649),叶绿素b质量浓度(mg·L−1):Cb=24.96D(649)−7.32D(665),叶绿素总质量浓度(mg·L−1):C总=Ca+Cb。类胡萝卜素质量浓度 (mg·L−1):Cc=[1 000D(470) −2.05Ca−114.8Cb]/245。叶绿体色素质量分数(mg·g−1)=(C×V提取液×稀释倍数)/(m样品×1 000)。其中:D(665)、D(649)和D(470)分别表示叶绿素溶液在波长665、649和470 nm处的吸光度,V提取液为提取液体积,m样品为样品质量。
-
花色素苷提取液的制备参照张洁等[18]和唐前瑞等[19]的方法,以体积分数1%的盐酸乙醇为对照,测定提取液在520 nm波长下的吸光度,以1 g叶片鲜质量(mFW)在10 mL提取液中改变0.1个吸光度值作为1个色素单位,则花色素苷相对含量表示为:A=D(520)/(0.1mFW)。
-
采用Excel 2016和SPSS 20.0统计分析软件处理数据,采用单因素方差(one-way ANOVA)分析并采用最小显著性差异法(LSD)检验不同季节2种刺柏属彩色树叶色参数和色素含量的差异显著性。对叶色参数与色素含量进行Pearson相关性检验,建立2种刺柏属彩色树的叶色参数与其色素含量的回归模型。
1.1. 材料
1.2. 试验设计
1.3. 测定指标
1.3.1. 叶色参数测定
1.3.2. 叶片光合色素含量测定
1.3.3. 花色素苷相对含量测定
1.4. 数据分析
-
由图1A可知:‘金叶疏枝’在春、夏、秋3季L*值差异不显著(P>0.05),冬季显著低于其他季节(P<0.05),最高值出现在夏季,最低值在冬季;‘金叶’在夏、秋、冬3季L*值变化不显著(P>0.05),春季显著低于其他季节(P<0.05),最高值出现在秋季,最低值在春季;春、夏、秋3季‘金叶疏枝’L*值均显著高于‘金叶’(P<0.05),冬季‘金叶疏枝’显著低于‘金叶’(P<0.05)。

Figure 1. Changes of L*,a*,b* values of two Juniperus colored trees in different seasons
由图1B可知:‘金叶疏枝’的a*值在春、夏、秋3季均为负值,且数值逐渐变大,冬季变为正值;其绝对值在夏季最高,春、夏季无显著差异(P>0.05),秋季绝对值显著低于春、夏季(P<0.05);冬季绝对值最低,显著低于其他季节(P<0.05)。‘金叶’的a*值在4季均为负值,数值先减小后增大;春、冬季无显著性差异(P>0.05),夏季分别与秋、冬季无显著性差异(P>0.05);绝对值最高出现在秋季,且显著高于春冬季(P<0.05),最低出现在春季。春、夏2季2树种a*值的绝对值无显著性差异(P>0.05);秋季‘金叶疏枝’绝对值显著低于‘金叶’(P<0.05);冬季‘金叶疏枝’a*值为正值,‘金叶’仍为负值,且绝对值显著高于‘金叶疏枝’(P<0.05)。
由图1C可知:‘金叶疏枝’的b*值4季均为正值,春、夏、秋3季之间无显著差异(P>0.05),冬季显著低于其他季节(P<0.05)。‘金叶’b*值4季也均为正值,秋、冬季无显著差异(P>0.05);春、夏、秋3季呈递进式显著增高(P<0.05),秋季最高,且显著高于春、夏季(P<0.05);其次是冬季,也显著高于春、夏季(P<0.05);绝对值最低出现在春季。春、夏、秋3季‘金叶疏枝’b*值均显著高于‘金叶’(P<0.05);冬季2种彩色针叶树b*值无显著性差异(P>0.05)。
-
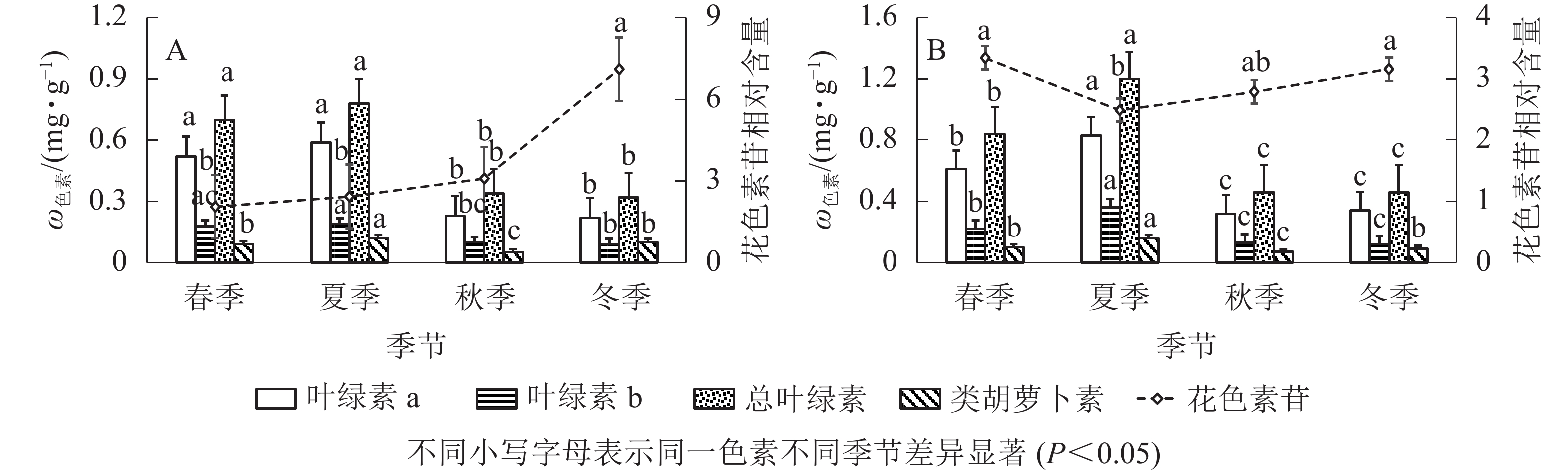
由图2A可知:‘金叶疏枝’叶绿素a、叶绿素b、总叶绿素在不同季节变化趋势相同,秋、冬季显著降低(P<0.05)。其中,叶绿素a质量分数在夏季最高,比秋、冬季显著高156.52%、168.18%(P<0.05);其次是春季,比秋、冬季显著高126.08%、136.36%(P<0.05),冬季最低。总叶绿素质量分数夏季最高,比秋、冬季显著高129.41%、143.75%(P<0.05),冬季最低。叶绿素b质量分数在夏、秋季间显著下降(P<0.05),春夏、秋冬之间无显著性差异;夏季最高,比秋、冬季显著高90.00%、111.11%(P<0.05);其次是春季,比冬季显著高100.00%(P<0.05)。‘金叶疏枝’类胡萝卜素质量分数呈现先增大后减小再增大趋势;夏季最高,比春、秋、冬3季显著高33.33%、140.00%、20.00%(P<0.05);其次是冬季,比秋季显著高100.00%(P<0.05),秋季最低。‘金叶疏枝’花色素苷相对含量在春、夏、秋季均无显著性差异(P>0.05),冬季比前3季显著高246.82%、192.59%、130.84%(P<0.05)。
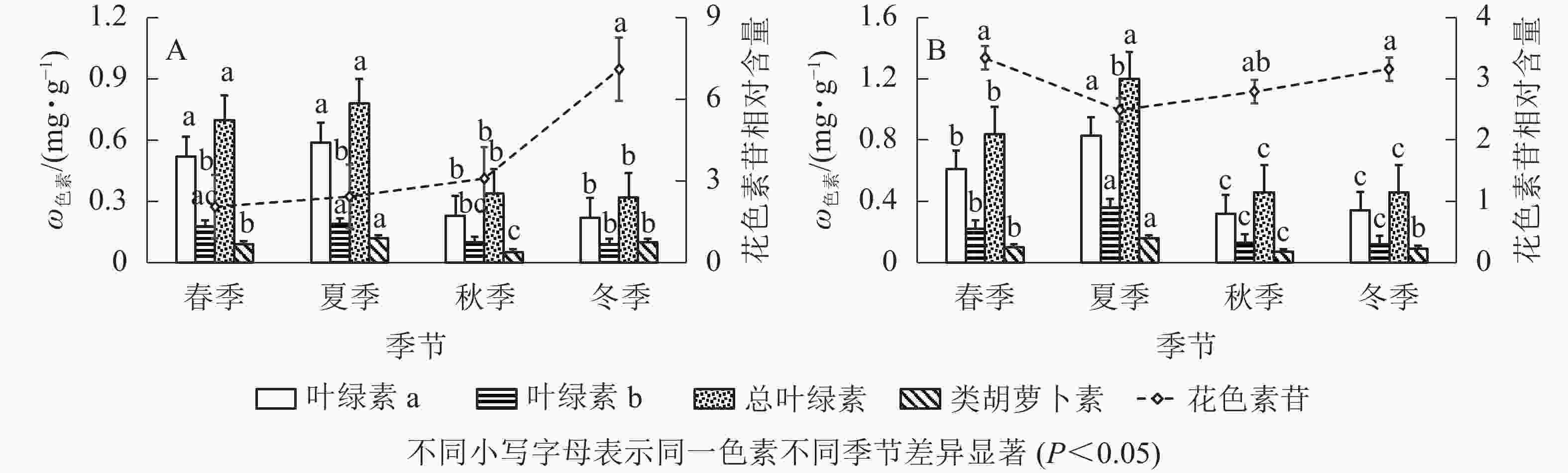
Figure 2. Changes of pigment content of two Juniperus colored trees in different seasons
由图2B可知:‘金叶’的叶绿素a、叶绿素b、总叶绿素质量分数随季节变化趋势相同,均为先增大后减小,且在夏季达到最高值。夏季叶绿素a比春、秋、冬季显著高36.06%、159.37%、144.11% (P<0.05),叶绿素b 比其他3季高63.63%、176.92%、200.00% (P<0.05),总叶绿素比其他3季高42.85%、160.86%、160.86% (P<0.05);春季叶绿素a比秋、冬显著高90.62%、79.41% (P<0.05),叶绿素b比秋、冬季高69.23%、83.33% (P<0.05),总叶绿素比秋、冬季高82.60%、82.60% (P<0.05);秋冬季无显著性差异(P>0.05)。‘金叶’类胡萝卜素质量分数随季节变化呈先增大后减小再增大趋势,夏季分别比春、秋、冬季显著高60.00%、128.57%、77.77% (P<0.05);春、冬季无显著性差异(P>0.05),但分别比秋季高42.85%、28.57%;秋季显著低于其他季节(P<0.05)。花色素苷相对含量呈现“V”字形变化,春到夏季显著降低(P<0.05),秋冬季显著增高(P<0.05),夏季比春、冬季显著低34.13%、26.90%(P<0.05)。
-
对2种刺柏属彩色树叶色参数与色素含量年变化的Pearson相关性分析(表1)可知:‘金叶疏枝’的L*、b*值与花色素苷相对含量呈极显著负相关(P<0.01);a*值与叶绿素a、总叶绿素质量分数呈显著负相关(P<0.05),与花色素苷相对含量呈极显著正相关(P<0.01)。‘金叶’的L*值与叶绿素a质量分数呈显著负相关(P<0.05);b*值与所有光合色素含量均呈极显著负相关(P<0.01)。
植物 叶色参数 叶绿素a 叶绿素b 总叶绿素 类胡萝卜素 花色素苷 ‘金叶疏枝’ L* 0.606 0.542 0.602 0.021 −0.975▲▲ a* −0.682▲ −0.573 −0.668▲ −0.144 0.934▲▲ b* 0.408 0.302 0.388 −0.097 −0.956▲▲ ‘金叶’ L* −0.831▲ −0.311 −0.363 −0.292 −0.519 a* 0.457 0.063 0.069 0.164 0.595 b* −0.983▲▲ −0.980▲▲ −0.984▲▲ −0.949▲▲ −0.212 说明:数据上的▲、▲▲分别表示在0.05、0.01水平上显著相关 Table 1. Correlation analysis between color parameters and pigment content of two Juniperus colored leaves
对2种刺柏属彩色树叶色参数与色素含量的季节性变化进行Pearson相关性分析,由表2可知:春季‘金叶疏枝’和‘金叶’的叶色参数与色素含量相关性均不显著。夏季‘金叶疏枝’a*值与叶绿素b、总叶绿素质量分数呈显著正相关(P<0.05),b*值与花色素苷相对含量呈显著负相关(P<0.05)。夏季‘金叶’L*值与所有光合色素含量均呈极显著负相关(P<0.01),与花色素苷相对含量呈显著负相关(P<0.05);a*值与类胡萝卜素质量分数呈显著正相关(P<0.05),与花色素苷相对含量呈极显著正相关(P<0.01);b*值与叶绿素a、总叶绿素、类胡萝卜素质量分数以及花色素苷相对含量呈极显著负相关(P<0.01),与叶绿素b质量分数呈显著负相关(P<0.05)。秋季‘金叶疏枝’L*值与花色素苷相对含量呈显著负相关(P<0.05);a*值与类胡萝卜素质量分数呈显著正相关(P<0.05),与花色素苷相对含量呈极显著正相关(P<0.01);b*值与花色素苷相对含量呈极显著负相关(P<0.01)。冬季‘金叶疏枝’L*值与光合色素呈显著负相关(P<0.05),‘金叶’叶色参数与色素含量秋冬季相关性均不显著(P>0.05)。
季节 植物 叶色参数 叶绿素a 叶绿素b 总叶绿素 类胡萝卜素 花色素苷 春季 ‘金叶疏枝’ L* 0.668 0.395 0.614 0.747 0.098 a* −0.693 −0.325 −0.614 −0.857 0.495 b* 0.657 0.441 0.618 0.709 −0.593 ‘金叶’ L* 0.073 0.015 0.057 0.071 0.671 a* 0.388 0.411 0.395 0.321 −0.095 b* −0.117 −0.131 −0.121 −0.058 0.311 夏季 ‘金叶疏枝’ L* −0.141 0.258 0.045 −0.449 0.005 a* 0.450 0.562▲ 0.558▲ −0.086 0.303 b* −0.471 −0.408 −0.492 −0.024 −0.585▲ ‘金叶’ L* −0.679▲▲ −0.723▲▲ −0.713▲▲ −0.647▲▲ −0.628▲ a* 0.456 0.473 0.474 0.610▲ 0.648▲▲ b* −0.649▲▲ −0.597▲ −0.644▲▲ −0.755▲▲ −0.771▲▲ 秋季 ‘金叶疏枝’ L* 0.298 0.146 0.246 −0.017 −0.624▲ a* −0.446 −0.424 −0.458 0.580▲ 0.901▲▲ b* 0.246 0.185 0.232 −0.214 −0.715▲▲ ‘金叶’ L* −0.079 0.040 −0.050 0.007 0.039 a* −0.313 −0.374 −0.333 −0.316 0.007 b* −0.359 −0.426 −0.381 −0.127 0.306 冬季 ‘金叶疏枝’ L* −0.645▲ −0.637▲ −0.645▲ −0.642▲ −0.468 a* −0.017 −0.090 −0.035 0.020 −0.170 b* −0.466 −0.544 −0.488 −0.536 −0.525 ‘金叶’ L* −0.173 −0.091 −0.152 −0.076 −0.498 a* −0.469 −0.540 −0.491 −0.010 −0.265 b* −0.506 −0.426 −0.488 −0.368 −0.358 说明:数据上的▲、▲▲分别表示在0.05、0.01水平上显著相关 Table 2. Correlation analysis between color parameters and pigment content of two Juniperus colored leaves in different seasons
-
为进一步探索刺柏属彩色树叶色参数与色素含量的关系,本研究建立了叶色参数与色素含量间的回归模型,为不同叶色期色素含量提供预测。
-
根据表1和表2中‘金叶疏枝’的叶色参数与色素含量相关性分析,以叶色参数L*、a*、b*的数值(l,a,b)为自变量,以与叶色参数有极显著相关关系的花色素苷相对含量为因变量(y),进行逐步线性回归拟合,得到叶色参数与花色素苷的年变化和秋季变化(其他季节回归方程均不显著)回归模型。由表3可知:年变化回归模型拟合系数为0.967,方程拟合度高,因此以a*值为自变量(L*、b*值作为变量被剔除)的回归模型可以准确预测‘金叶疏枝’的花色素苷相对含量年变化。季节变化逐步回归方程拟合系数为0.884,a*值和b*值共同决定秋季‘金叶疏枝’花色素苷相对含量的变化。
色素 逐步回归方程 拟合系数(R2) P (年变化)花色素苷 y=5.782+0.126a 0.967 <0.001 (秋季)花色素苷 y=6.283+0.047a−0.049b 0.884 <0.001 说明:a、b分别为叶色参数a*、b*的数值 Table 3. Stepwise regression model for leaf color parameter and pigment content of J. communis ‘Gold Cone’
-
根据表1和表2中‘金叶’的叶色参数与色素含量相关性分析,以叶色参数L*、a*、b*的数值(l,a,b)为自变量,分别以与其有极显著相关关系的色素含量值为因变量(y),进行逐步线性回归拟合,得到叶色参数与色素含量年变化和夏季变化(其他季节回归方程均不显著)回归模型。由表4可知:b*值与叶绿素a、叶绿素b、总叶绿素及类胡萝卜素质量分数的年变化回归模型的方程拟合系数均大于0.850,拟合程度高,因此认为以b*值为自变量的回归方程能够准确预测‘金叶’的光合色素含量的年变化。此外,夏季b*值能准确预测叶绿素a质量分数和花色素苷相对含量的变化,夏季L*值能准确预测叶绿素b、总叶绿素以及类胡萝卜素质量分数的变化。
色素 逐步回归方程 拟合系数(R2) P (年变化)叶绿素a y=1.757−0.036b 0.958 <0.001 叶绿素b y=0.801−0.017b 0.950 0.001 总叶绿素 y=2.558−0.054b 0.959 <0.001 类胡萝卜素 y=0.324−0.006b 0.875 0.004 (夏季)叶绿素a y=1.148−0.011b 0.847 0.002 叶绿素b y=0.943−0.011l 0.743 0.008 总叶绿素 y=2.384−0.022l 0.806 0.004 类胡萝卜素 y=0.233−0.002l 0.856 0.005 花色素苷 y=5.568−0.093b 0.844 0.018 说明:l、b分别为叶色参数L*、b*的数值 Table 4. Stepwise regression model for leaf color parameters and pigment content of J. ×media ‘Pfitzeriana Aurea’
2.1. 不同季节2种刺柏属彩色树叶色参数变化
2.2. 不同季节2种刺柏属彩色树色素含量变化
2.3. 2种刺柏属彩色树叶色参数与色素含量的相关性分析
2.4. 2种刺柏属彩色树叶色参数与色素含量回归模型
2.4.1. ‘金叶疏枝’叶色参数与色素含量逐步回归模型
2.4.2. ‘金叶’叶色参数与色素含量逐步回归模型
-
彩色植物的叶片色泽是评价其观赏性和综合美景度的重要指标之一[20],色泽鲜亮、呈色周期长的彩色植物通常是园林造景和环境绿化的优势树种[20-21]。通过测定彩色树的叶色参数,量化叶片色泽数值,可减小主观评价对植物观赏价值评估产生的误差[22]。本研究中,欧洲刺柏‘金叶疏枝’的L*、b*值变化趋势相同,均在春、夏、秋3季保持较高水平,b*值在4季中均为正值,即整年中色相始终偏黄,认为最佳观赏季为春、夏、秋3季,与前人研究[6]基本一致;夏季明度和偏黄程度最高,叶色最为鲜亮,认为夏季观赏价值最高。本研究中,鹿角桧‘金叶’的L*值春季最低,之后显著增高,且在夏、秋、冬3季无明显差异,b*值在4季中均为正值,即色相始终偏黄,且秋季黄色相程度最深;认为最佳观赏季为夏、秋2季,与前人研究一致[8];a*值始终为负值,即整年中色相始终偏绿,冬季并未出现褐色变化,与前人研究有差异,可能是供试植株为2年生扦插苗,与成年株的越冬生长有差异。
对‘金叶疏枝’和‘金叶’叶色参数的差异比较发现:前者L*、b*值在春、夏、秋3季明显高于后者。结合前人研究[23],认为春、夏、秋3季‘金叶疏枝’的叶色观赏性高于‘金叶’;两者的a*值在春、夏季无明显差异,秋、冬季前者叶色转红而后者仍旧为绿色叶,认为冬季两者因叶色不同而具有不同观赏价值。
-
植物叶色变化的直接原因是叶绿素、类胡萝卜素和花色素苷含量的时空分布变化,而色素种类和含量的变化受内在遗传因子和外部环境因子共同影响[23-25]。本研究中,2种彩色树的叶绿素a、叶绿素b和总叶绿素变化趋势相同,与季鹏章等[26]的研究结果一致。受秋冬季温度降低和光照减少影响,秋、冬2季‘金叶疏枝’叶绿素分解加快,叶绿素显著减少;而低温诱导了花色素苷的合成加快[19, 27],花色素苷在冬季显著增加。春季‘金叶’花色素苷相对含量较高,可能是因为春季叶片始萌,受春季较大昼夜温差影响,花色素苷分解速度减缓;至夏季,光照增加和温度上升使得叶绿素显著增加,占主导优势[28-29],叶色发生变化。
-
研究表明,许多彩色植物叶色参数与色素种类及含量存在显著相关关系,建立线性回归模型能较准确地通过叶色参数变化预估色素含量变化[9, 13]。但不同时期彩色植物色素变化不同,叶色参数波动也存在差异[30]。本研究中,秋季‘金叶疏枝’光合色素质量分数显著下降,花色素苷相对含量小幅度上升,此时叶色参数L*值、b*值无明显波动,a*值显著上升,说明花色素苷的变化影响a*值,但此时光合色素仍占主导,故色相仍旧偏绿。而秋冬之际花色素苷显著增加,L*值和b*值显著降低,a*值从负变正,叶色转红;分析发现花色素苷相对含量与a*值极显著正相关,与L*值和b*值呈极显著负相关,与郭欢欢等[12]对秋季黄连木Pistacia chinensis色素含量与叶色参数的研究结果和齐睿等[13]对红叶石楠Photinia×fraseri上位叶色参数与花色素苷含量的研究结果相似。
本研究发现:夏季‘金叶’光合色素质量分数显著增加,花色素苷相对含量显著下降;L*值和b*值显著上升,a*值显著下降;植物表现为叶色鲜亮,黄绿色加深,观赏价值提高。推测在春夏之际色素含量基本相同的变化幅度下,花色素苷相对含量变化对L*值和b*值的影响程度更高,且为负相关。夏秋之际光合色素质量分数显著下降,花色素苷相对含量无明显变化;同时L*值无明显变化,b*值显著上升,a*值无明显波动。说明夏秋之际光合色素的变化显著影响b*值,且呈负相关。冬季类胡萝卜素质量分数上升,花色素苷相对含量显著上升,但叶色参数几乎没有变化。推测此时叶绿素质量分数较高,仍占主导,而类胡萝卜素质量分数和花色素苷相对含量的上升也对植物老叶细胞越冬起到一定保护作用[31-32]。
朱书香等[9]研究了4种李属Prunus彩叶植物的叶色参数与色素含量的关系,发现尽管色素变化趋势基本相似,但叶色参数和色素含量的关系却因树种各异不尽相同。本研究发现:欧洲刺柏‘金叶疏枝’和鹿角桧‘金叶’属刺柏属,光合色素变化趋势基本相同;但‘金叶疏枝’叶色参数主要受花色素苷相对含量影响,以a*值为自变量建立的回归模型可以较准确预估花色素苷相对含量,以a*和b*的值为自变量建立的回归模型可以较准确预估秋季花色素苷相对含量。‘金叶’的光合色素主要影响L*和b*的值,尤其是b*值,以b*的值为自变量建立的回归方程可以较准确预估光合色素的变化。
‘金叶疏枝’和‘金叶’为彩色植物中的彩色针叶树代表性树种。对彩色阔叶树的研究发现,叶片微观结构和生理代谢因子可调控叶色变化。彩色针叶树和普通彩色阔叶树的叶片微观结构不同,光合生理也不尽相同,因此开展彩色针叶树叶色变化周期内内部生理因子调控作用研究很有必要。
3.1. 2种刺柏属彩色树叶色参数变化与观赏性分析
3.2. 2种刺柏属彩色树色素含量变化
3.3. 2种刺柏属彩色树叶色参数与色素含量相关关系
-
不同季节欧洲刺柏‘金叶疏枝’和鹿角桧‘金叶’的叶色参数变化趋势不同。2种刺柏属彩色树的叶色参数与色素含量关系显著,但不同树种不同时期存在差异;‘金叶疏枝’花色素苷相对含量与L*、b*值呈极显著负相关,与a*值呈极显著正相关;‘金叶’光合色素与b*值呈极显著负相关。欧洲刺柏‘金叶疏枝’和鹿角桧‘金叶’的叶色参数与色素含量回归方程的建立,证明彩色针叶树的叶色参数可以精确预估色素含量,为人工调控彩色针叶树叶色变化及优良彩色针叶树的选育提供了理论基础。
 DownLoad:
DownLoad: