






-
毛竹Phyllostachys edulis属于禾本科Poaceae竹亚科Bambusoideae刚竹属Phyllostachys,在中国分布范围广,被广泛应用于观赏、木材加工、食用等方面,具有很高的经济和生态价值[1]。毛竹喜湿润环境[2],干旱、盐碱、低温等非生物胁迫将会严重限制毛竹的推广与应用[3]。据报道,持续的高温少雨导致了咸宁市毛竹林大批量死亡[4],湖州市严重干旱导致大量竹笋退笋不能成竹[5],非生物胁迫已经严重影响产地竹产业及社会经济发展。目前已经展开了对毛竹逆境胁迫机理的系列研究,有利于实现毛竹可持续经营,推动竹产业经济高质量可持续发展。
bHLH (basic helix-loop-helix)转录因子在真核生物中广泛存在,主要参与生物和非生物胁迫响应[6−7]、激素信号转导途径[8]、植物的胚胎[9]、雌蕊发育[10]及开花调控[11]。在毛竹中存在153个bHLH[3]家族成员,在拟南芥Arabidopsis thaliana和水稻Oryza sativa中分别有162个[12]和167个[13],是植物中仅次于MYB的第二大基因家族[14−16]。HEIM 等[17]根据基因结构,又将拟南芥bHLH家族划分为 12 个亚家族。大多数bHLH转录因子在植物非生物胁迫信号通路中起着重要的调控作用。FAN等[18]分析了高粱Sorghum bicolor在胁迫处理下bHLH家族的表达模式,鉴定出12个可能和非生物胁迫相关的SbbHLH基因。本研究通过对毛竹实生苗进行非生物胁迫处理,利用转录组数据筛选出1条差异表达基因,命名为PhebHLH6,并对其进行生物信息学分析及表达模式探究,初步讨论了其潜在基因功能,以期为毛竹抗逆基因功能研究提供一定的理论基础。
-
毛竹26个组织转录组数据来自美国国家生物技术信息中心 (NCBI)数据库(GSE169067)[19]。毛竹种子采自广西桂林,在浙江农林大学玻璃温室(温度为26 ℃;湿度为80%)培养1个月,采集幼嫩叶片置于−80 ℃保存,用于后续研究。对生长至2月龄的毛竹实生苗进行处理,设置对照(ck)组观察取样,并在处理后的0、3、24 h分别取样保存至−80 ℃液氮。
-
对筛选得到的差异表达基因PhebHLH6进行克隆及表达模式分析。使用Oligo 7软件[20]设计蛋白编码区(CDS)全长引物,引物由杭州有康生物科技有限公司合成,引物序列参见表1。提取毛竹幼叶RNA并反转录为cDNA作为模板,参考表1中引物进行PCR扩增。反应体系为50.0 μL体系:2×Mix为25.0 μL,10.0 μmol·L−1上下游引物各1.0 μL,cDNA为1.0 μL ,ddH2O为22.0 μL。PCR反应程序:95 ℃预变性 5 min,95 ℃变性30 s,60 ℃退火30 s,72 ℃延伸60 s,34个循环,72 ℃延伸5 min。将回收产物连接pMD18-T克隆载体并转化大肠埃希菌Escherichia coli DH5α,菌检选取阳性克隆送杭州有康生物技术有限公司测序。
用途 引物名称 引物序列(5′→3′) 基因克隆 PhebHLH6-F ATGGACGCGGACATGGGCGACA PhebHLH6-R CTAATAGCTCATCGAGCTCGGG
GGGCTTC实时荧光定量
PCR (RT-qPCR)Q-PhebHLH6-F CGAGAAGCTATACGCGATCC Q-PhebHLH6-F CTGCAGCTGCTGGATGTAAT Q-NTB-F TCTTGTTTGACACCGAAGAGGAG Q-NTB-F AATAGCTGTCCCTGGAGGAGTTT Table 1. Primers used in gene clone and quantitative real-time PCR
-
将克隆获取的PhebHLH6蛋白序列,利用ExPASy的在线软件Protparam (
http://web.expasy.org/protparam/ )和ProtScale (https://web.expasy.org/protscale/ )分别分析氨基酸理化性质和蛋白的亲/疏水性。分别利用PRABI的SOPMA (https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.Html )和Swiss-Model (http://swissmodel. expasy.org/interactive )进行蛋白二级、三级结构分析。蛋白保守结构域使用NCBI Conserved Domain Database (https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi )进行分析。根据拟南芥、水稻、玉米Zea mays、二穗短柄草Brachypodium distachyon、芸香竹Bonia amplexicaulis、瓜多竹Guadua angustifolia、Olyra latifolia和Raddia guianensis [21]蛋白数据库,进行BLAST序列比对,进一步比对得到PhebHLH6的同源基因氨基酸序列。利用MEGA11.0软件Neighbor-Joining算法,自检举1 000次,构建系统进化树。 -
使用TBtools[22]软件提取基因5′UTR上游2 000 bp序列,并使用 PlantCARE在线软件对其进行分析,预测启动子核心元件和基因结合位点。
-
参考毛竹26个组织转录组数据,使用转录数据中的FPKM值并利用TBtools软件绘制热图,对PhebHLH6基因进行不同组织表达量分析。
-
毛竹种子采自广西桂林。将发芽的种子置于温度为(25±2) ℃、相对湿度为(70±10)%的玻璃温室中,用1/4霍格兰营养液水培,每周更换1次,培养至2月龄。分别设置对照、氯化钠(NaCl)、聚乙二醇(PEG)、水杨酸(SA)、脱落酸(ABA) 5个处理。分别使用200 mmol·L−1 NaCl和质量分数为25% 的PEG营养液进行盐和干旱胁迫。此外,分别采用1 mmol·L−1和1 μmol·L−1的SA和ABA营养液进行处理。在处理0、3、24 h分别采集毛竹叶片,每个时间点采集3个生物学重复,液氮速冻,保存至−80 ℃冰箱以便后续分析使用。
使用Primer 3 version 0.4.0设计定量引物,将其在NCBI (Primer designing tool)进行引物特异性分析。实时荧光PCR (RT-qPCR)反应体系如下:cDNA(已稀释)为1.0 μL,上、下游引物各0.5 μL (10.0 μmol·L−1),NovoStart®SYBR qPCR SuperMix plus (Novoprotein)为5.0 μL,ddH2O为3.0 μL。反应程序为:95 ℃ 5 min;95 ℃ 5 s, 60 ℃ 30 s, 72 ℃ 60 s,30 循环;72 ℃ 10 min。39个循环,3次重复。数据分析采用$2^{-\Delta\Delta Ct}$方法[23]。仪器为CFX96TM Real-Time PCR Detection System (Bio-Rad)。内参基因为NTB基因[24]。数据结果采用TBtools软件绘制热图分析,以10为底取对数值。
-
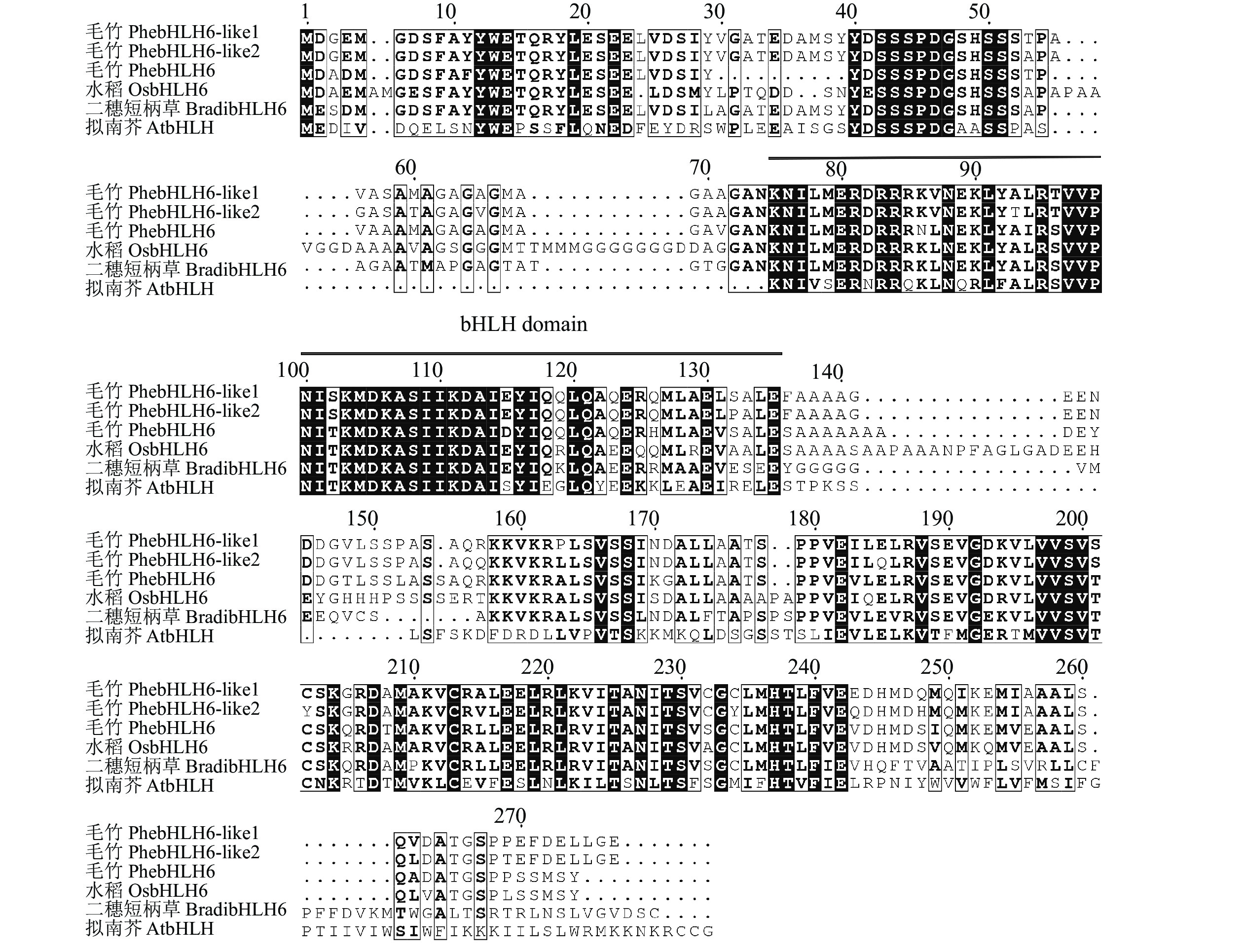
利用特异性引物进行PCR扩增,获得PhebHLH6编码氨基酸序列如图1所示。测序结果显示:编码区序列长801 bp,编码266个氨基酸。保守结构域分析结果表明:PhebHLH6蛋白在64~125氨基酸位置具有完整的bHLH保守结构域,说明该基因是典型的bHLH家族成员。

Figure 1. Alignment of bHLH6 amino acid sequences from different species
-
蛋白序列分析结果表明:PhebHLH6的蛋白质分子量为28.651 kDa,等电点为5.04,平均疏水性为−0.102,蛋白质不稳定指数55.58,脂肪指数为88.83。上述结果表明:PhebHLH6蛋白为亲水性不稳定蛋白。二级蛋白结构预测结果显示:PhebHLH6的蛋白二级结构占比分别为α-螺旋53.76%,β-转角2.63%,延伸链9.02%,无规则卷曲结构34.59%。使用SWISS-MODEL在线比对蛋白序列,预测PhebHLH6蛋白的三级结构并建立空间模型,并与水稻OsbHLH6比较分析,发现其三级结构相似,功能可能较为保守。
-
顺式作用元件分析结果表明(表2):PhebHLH6启动子上游2 000 bp除包含TATA-box和CAAT-box等核心启动元件外,还包含ABRE元件(响应ABA)、TGACG-motif [响应茉莉酸甲酯(MEJA)]、CGTCA-motif (响应MEJA)、TGA-element (响应生长素)、AAGAA-motif (响应ABA)等一系列非生物胁迫及激素响应元件。上述结果表明:PhebHLH6可能参与到了多种非生物胁迫及激素响应途径中。
作用元件 序列 数量 功能 作用元件 序列 数量 功能 ABRE CACGTG 9 脱落酸响应元件 Sp1 GGGCGG 1 光响应元件 ARE AAACCA 1 厌氧诱导顺式作用元件 chs-CMA2a TCACTTGA 1 光响应元件 CAAT-box CCAAT 15 启动子和增强子区域调控元件 TGA-element AACGAC 2 生长素响应元件 CGTCA-motif CGTCA 5 茉莉酸甲酯响应元件 O2-site GATGATGTGG 1 玉米醇溶蛋白代谢调节元件 MRE AACCTAA 1 光响应元件 TATA-box TATA 35 核心启动子元件 G-Box TACGTG 3 光响应元件 TGACG-motif TGACG 5 茉莉酸甲酯响应元件 Table 2. Cis-element analysis of PhebHLH6 gene promoter
-
蛋白比对结果显示:PhebHLH6与毛竹PhebHLH6-like基因及水稻、拟南芥等物种同源基因的氨基酸序列相似,均具有完整的bHLH结构域,推测其可能拥有相似的基因功能。为进一步探究PhebHLH6功能,本研究利用毛竹、水稻、拟南芥等9个物种的bHLH6同源蛋白序列构建生物进化树。如图2所示:毛竹PhebHLH6与其他2种木本竹同源基因的蛋白聚在一起,2种草本竹聚在一起,推测木本竹和草本竹在进化过程中功能出现了一定分化。此外,竹子中的bHLH蛋白与水稻、玉米的蛋白以较高的置信度聚在一起,说明bHLH基因序列在进化方面较为保守,其功能可能具有一定相似性。
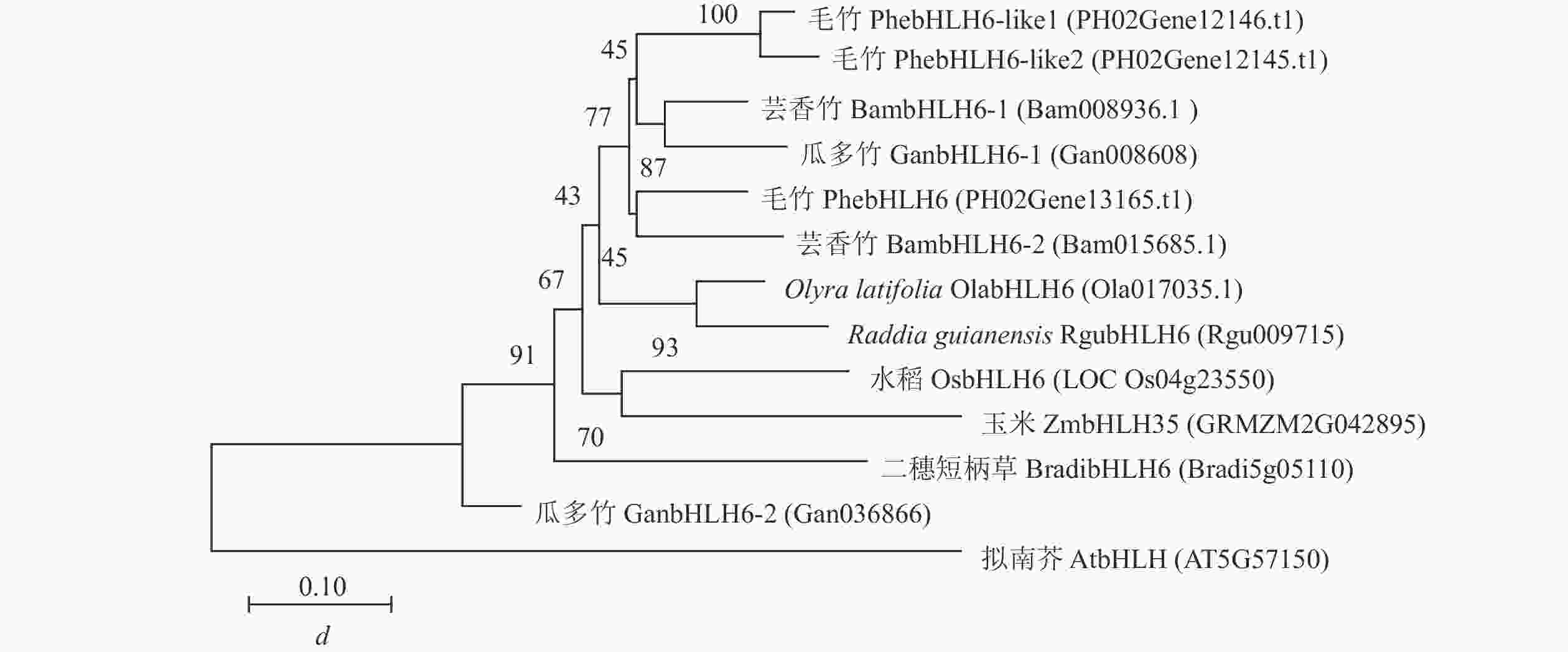
Figure 2. Phylogenetic tree construction of BHLH6-like amino acid sequences
-
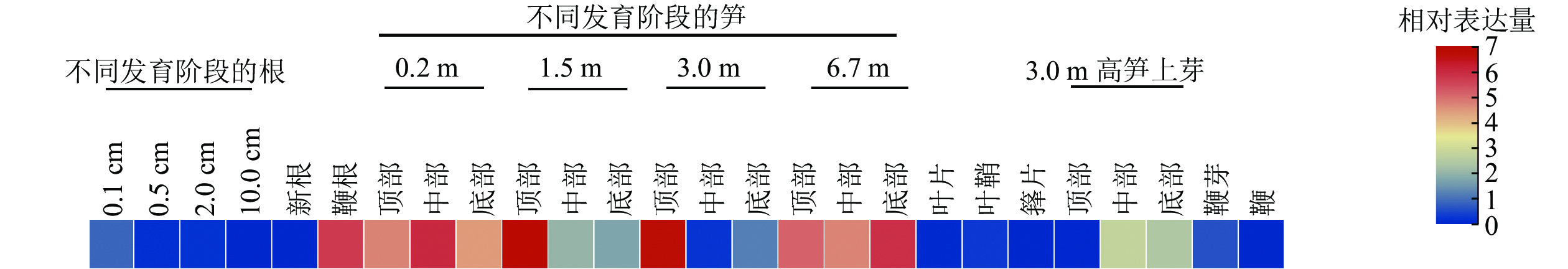
分析了PhebHLH6在毛竹26个组织中的表达水平(图3)。该基因在毛竹不同发育阶段的根、叶片、芽和鞭中相对表达量都很低,在笋中相对表达量相对较高,其中1.5和3.0 m的笋顶部相对表达量最高,6.7 m的笋各部位次之。此外,在鞭根和3.0 m高笋的侧芽中下部,PhebHLH6也有一定的表达。
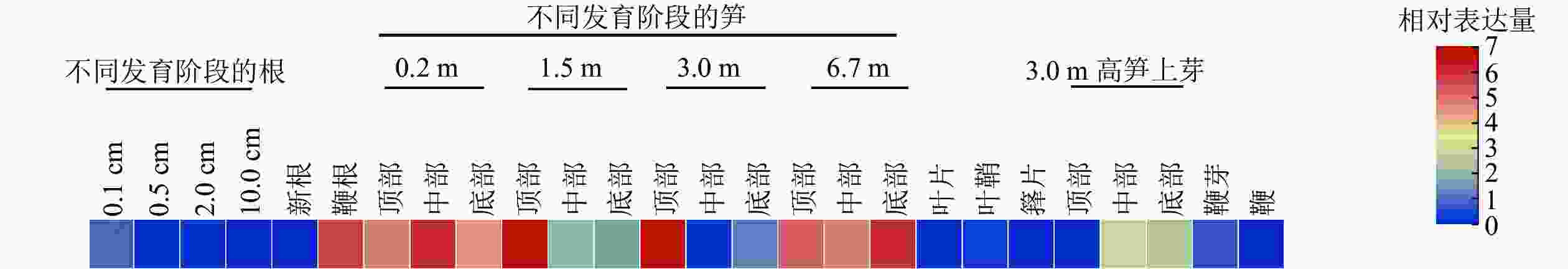
Figure 3. Expression level of PhebHLH6 gene in different tissues
-
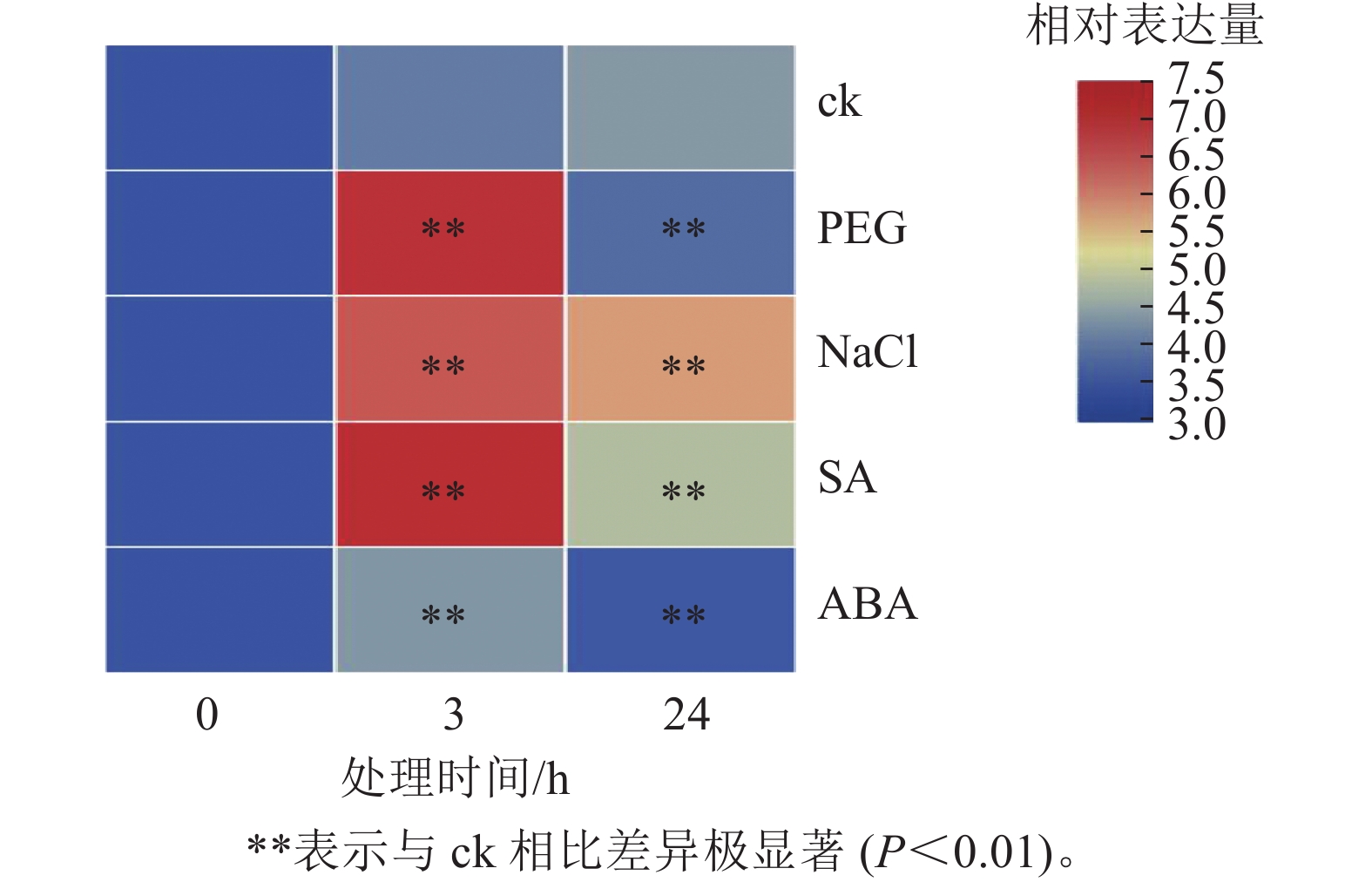
为探究PhebHLH6在非生物胁迫及激素处理下的表达模式,对2月龄毛竹实生苗进行了干旱胁迫、盐胁迫、SA和ABA处理,并通过荧光定量的方式对该基因相对表达量进行了检测(图4)。PhebHLH6基因对干旱和盐胁迫均具有强烈的响应,对SA和ABA也有一定的应答作用。
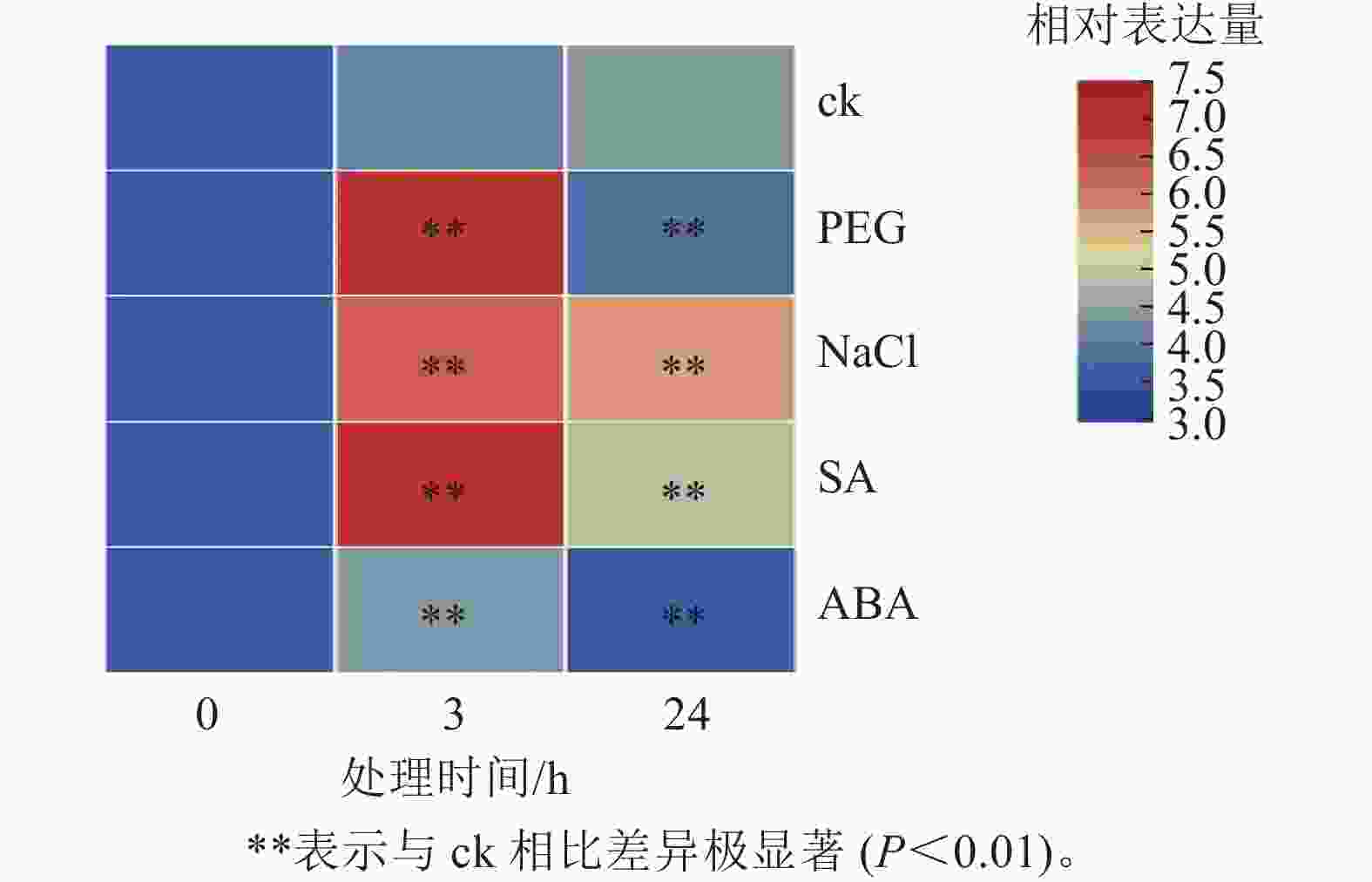
Figure 4. Expression patterns of PhebHLH6 under abiotic stress and hormone treatment
-
如图4所示:干旱胁迫下,与ck相比,PhebHLH6的相对表达量呈先上升后下降趋势,处理3 h后相对表达量为未处理的20倍左右,24 h后相对表达量低于ck,说明PhebHLH6在处理早期受到强烈诱导,处理后期诱导信号减弱。高盐处理下,PhebHLH6在处理3 h后相对表达量明显上升,在处理24 h后相对表达量有一定下降,但与ck相比,相对表达量仍有极显著上调(P<0.01),说明PhebHLH6持续被高盐诱导,但在处理早期诱导程度高于后期。
-
与ck相比,PhebHLH6 在SA处理3 h后相对表达量升高20倍,呈现极显著上调趋势(图4,P<0.01)。在处理24 h后,相对表达量呈下降趋势,说明PhebHLH6在处理后24 h期间持续被SA诱导,但诱导作用主要发生于早期,在处理24 h后诱导作用有所下降。ABA处理下,PhebHLH6相对表达量前期有微弱上调,24 h相对表达量呈下降趋势,说明ABA在处理前期对PhebHLH6诱导作用不强。
-
有研究指出:bHLH家族成员主要作用于植物的信号转导、生长发育及响应生物及非生物胁迫等重要途径[25]。竹子与水稻同为禾本科植物,进化关系较近,毛竹中很多基因功能研究往往参考水稻展开[1]。通过同源序列比对结果可知:PhebHLH6与水稻同源基因蛋白序列相似度为72.35%,序列相似度很高,其功能可能具有相似性。HE等[26]研究发现:水稻OsbHLH6在嫩芽和根中特异性表达,其过表达植株在低磷情况下比野生型拥有更长根系。PhebHLH6在毛竹不同组织中的表达水平分析结果表明:该基因在1.5和3.0 m的笋顶端表达量最高,推测该基因可能与毛竹笋芽顶端生长发育相关。
bHLH家族成员在植物响应逆境胁迫中也发挥重要作用。如张子佳等[27]对水稻bHLH家族响应环境胁迫表达谱分析发现:大部分家族成员响应干旱胁迫和参与ABA调控途径。水稻中OsbHLH148可以通过形成复合体的形式调控水稻抗干旱能力[28]。 OsbHLH6 (RERJ1)通过在细胞核与细胞质间的移动,动态调节水杨酸和茉莉酸(JA)激素信号来控制[29]。OsJAZ9作为bHLH家族的转录因子,通过与OsNINJA和OsbHLH蛋白形成转录调控复合物控制JA合成通路来调控植株的耐盐性[30]。OsbHLH6基因在2004年首次被鉴定为JA响应基因[31],且研究发现该基因受外界伤害和干旱胁迫诱导表达上调[32]。与之相同的是,毛竹PhebHLH6也会受到干旱和盐胁迫的强烈诱导。此外,文献报道OsbHLH6可以通过动态调节SA和JA信号通路来控制抗病能力[28]。本研究表明:毛竹PhebHLH6受SA强烈诱导,在处理后表达量极显著上升,与水稻中同源基因OsbHLH6表达模式一致,推测其可能具有功能相似性,但这些推论还需进一步验证。
-
本研究从毛竹中比对克隆得到1条bHLH同源基因,命名为PhebHLH6。PhebHLH6基因编码区长度为801 bp,编码266个氨基酸,序列包含完整的bHLH结构域,属于bHLH转录因子。组织特异性表达水平表明:该基因主要在毛竹1.5和3.0 m笋顶端表达,推测其可能和毛竹生长发育相关。此外,PhebHLH6对非生物胁迫和激素处理均有响应,其中受干旱、高盐胁迫以及SA激素处理强烈诱导,受ABA轻微诱导,表明PhebHLH6可能在植物逆境响应和激素初期信号转导中发挥着重要的作用。
Cloning and expression analysis of PhebHLH6 gene from Phyllostachys edulis
doi: 10.11833/j.issn.2095-0756.20220553
- Received Date: 2022-08-30
- Accepted Date: 2022-12-18
- Rev Recd Date: 2022-12-14
- Available Online: 2023-07-13
- Publish Date: 2023-08-20
-
Key words:
- Phyllostachys edulis /
- PhebHLH6 /
- gene clone /
- expression analysis
Abstract:
| Citation: | ZHUO Juan, HOU Dan, LIN Xinchun. Cloning and expression analysis of PhebHLH6 gene from Phyllostachys edulis[J]. Journal of Zhejiang A&F University, 2023, 40(4): 731-737. DOI: 10.11833/j.issn.2095-0756.20220553

|

 DownLoad:
DownLoad: