




-
土壤酶是土壤生化过程的积极参与者,是生态系统物质循环和能量流动过程中最活跃的生物活性物质,通常与土壤微生物的代谢速率和养分的生化循环密切相关[1]。前人研究发现,不同养分在土壤中的释放和储存,腐殖质的形成和变化都与土壤酶的种类和活力有着紧密的联系。土壤酶在森林生态系统的养分循环和能量代谢中起到了非常关键的作用,被视为土壤生态系统的核心部分[2−3]。土壤酶主要来源于植物根系、土壤动物、微生物细胞分泌物及残体的分解物,是生态系统中生化过程和养分循环的主要调节者,在推动营养元素转化、生态系统功能调节等方面发挥着非常关键的作用[4]。
不同植被类型对土壤养分的富集和再分配以及养分流失具有重要影响,进而对土壤酶活性产生不同影响。近年来,国内外学者高度重视土壤酶活性的研究,不同空间尺度的土壤酶活性已得到广泛研究[5−10]。刘顺等[11]研究发现,坡向间植被类型通过土壤性质驱动土壤酶活性。贺兰山是干旱区具有完整垂直带谱的山地生物多样性宝库,植被类型具有明显的垂直地带性。已有研究显示:贺兰山东坡海拔显著影响土壤胞外酶活性,随着海拔的升高酶活性整体呈现上升趋势[12],β-葡萄糖苷酶(β-G)酶活性随海拔升高呈先增后减趋势[13]。但贺兰山西坡不同海拔典型植被类型土壤酶活性的分布特性尚不明确。本研究以贺兰山西坡不同植被类型土壤为研究对象,对不同植被类型土壤理化性质和土壤酶活性进行综合研究,以了解贺兰山西坡不同植被类型下土壤酶活性变化情况及影响因素,旨在为干旱区森林生态系统土壤酶活性变化、养分循环模式和调节机制研究提供依据。
-
贺兰山地处宁夏与内蒙古的接壤地带,具以山区为主要特点的典型大陆性季风气候。年均气温为8.6 ℃,年均降水量为209.2 mm,最高为627.5 mm,降水在6—8月最为集中,年均日照时数为3 100.0 h。贺兰山的荒漠草原、森林、亚高山草甸是中国中温带半干旱与干旱地区山地生态系统的典型代表[12]。在不同海拔植被类型中,水热交替具有较强的规律。贺兰山西坡土壤具有明显的垂直分布规律,沿海拔上升不同植被类型土壤自下而上分别为灰漠土、棕钙土、灰褐土、亚高山草甸土。
-
2022年8月,沿贺兰山西坡1 300~2 700 m海拔范围内(38°19′~39°22′ N, 105°49′~106°41′ E),自下而上分别选取具有代表性的荒漠草原(海拔1 349 m)、灰榆Ulmus glaucescens林(1 905 m)、蒙古扁桃Amygdalus mongolica灌丛(2 134 m)、油松Pinus tabuliformis林(2 150 m)、青海云杉Picea crassifoliai-山杨Populus davidiana混交林(2 160 m)、青海云杉林(2 635 m)和亚高山草甸(2 664 m)等7种典型植被类型作为样地。山地土层厚度不均,不同样地取样深度统一确定为0~10和10~20 cm,在同一海拔每种植被类型设3个样地,样地间隔大于100 m。在每个海拔样地内随机设置3个样方,乔木、灌木和草本标准样方大小分别为20 m×20 m、10 m×10 m、1 m×1 m,共21个样方。在每个样方内采用五点混合法取样,除去地表凋落物层后,用直径4 cm的土钻采集0~10、10~20 cm的土壤样品,将样品充分混合,放入自密封袋中,然后用冷藏箱将其运回实验室。从土壤中去除可见的粗根和石块后,用2 mm的筛子对土壤样品进行筛分。将筛选后的土壤样品分为2组,其中一组放置于阴凉环境中自然风干,测定其理化特性;另一组置于4 ℃的冷藏条件下,以检测土壤酶活性和其他相关指标。
-
采用环刀法测定土壤容重(BD),烘干法测定土壤含水率(SWC),pH计测定土壤pH值(土水质量比为1.0∶2.5)[6]。采用重铬酸钾氧化外加热法测定有机碳(SOC),半微量凯氏定氮法测定全氮(TN),钼锑抗比色法测定全磷(TP)[9],氯化钾溶液提取-分光光度法测定铵态氮(NH4 +-N),比色法测定有效磷(AP)[12]。
采用3, 5-二硝基水杨酸比色法测定蔗糖酶(Inv)活性与淀粉酶(Amy)活性[11−12],以1 g土样24 h催化生成还原糖的毫克数表示;采用比色法测定α-葡糖苷酶(α-G)活性进行;通过硝基苯葡糖苷法测定β-葡糖苷酶(β-G)活性;β-木糖苷酶活性(BXYL)、纤维二糖水解酶(CBH)采用微孔板荧光法,利用多功能酶标仪(SpectraMax M5)测定其荧光度[14−15]
-
用 Excel整理数据,用 SPSS对数据进行统计和分析。利用单因素方差分析(one-way ANOVA)比较不同理化性质及酶活性差异(α=0.05);用双因素方差分析检验不同植被类型和不同土层下土壤理化性质和酶活性交互作用;利用Origin 2022进行相关性分析并绘制热图,采用Canoco 5.0蒙特卡洛检验分析理化性质对土壤酶活性的影响。
-
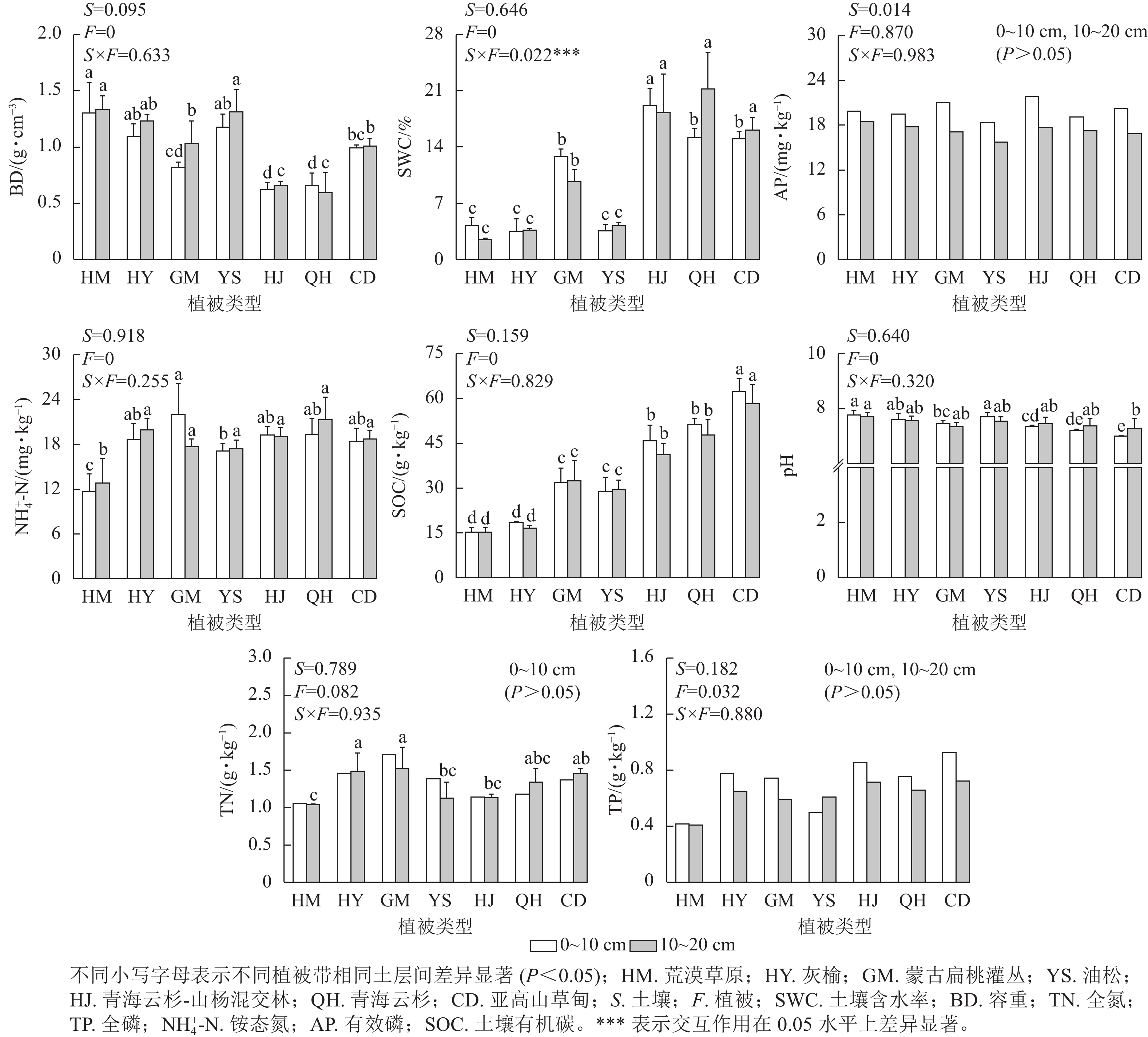
如图1所示:在0~10与10~20 cm土层,有机碳质量分数在亚高山草甸最高,其质量分数为62.30、58.28 g·kg−1;在荒漠草原最低,为15.31、15.32 g·kg−1。在不同植被带中,10~20 cm土层中土壤容重整体比0~10 cm土层高,在荒漠草原植被带不同土层土壤容重质量分数均高于其他植被带;有效磷质量分数在0~10 cm土层高于10~20 cm,各海拔间其质量分数无显著差异;0~10 cm土层,全氮质量分数在不同植被带无显著差异,10~20 cm土层中,蒙古扁桃灌丛全氮质量分数最高,为1.52 g·kg−1,荒漠草原最低,为1.04 g·kg−1。通过对土层、植被带及其交互作用对土壤理化性质的双因素方差分析,结果表明:不同植被带对土壤含水率、容重、pH及全磷、铵态氮、有机碳质量分数产生显著影响,土壤含水率与有机碳质量分数随海拔上升呈增加趋势,土壤容重随海拔上升呈下降趋势;土壤全氮、铵态氮质量分数随海拔升高呈先上升后下降;全磷、有效磷质量分数及pH无显著差异。土层对以上指标均未产生显著影响。土层与植被类型及其交互作用对土壤含水率产生显著影响。

图 1 不同植被类型土壤基本理化性质
Figure 1. Basic physical and chemical properties of soils at different vegetation types
-
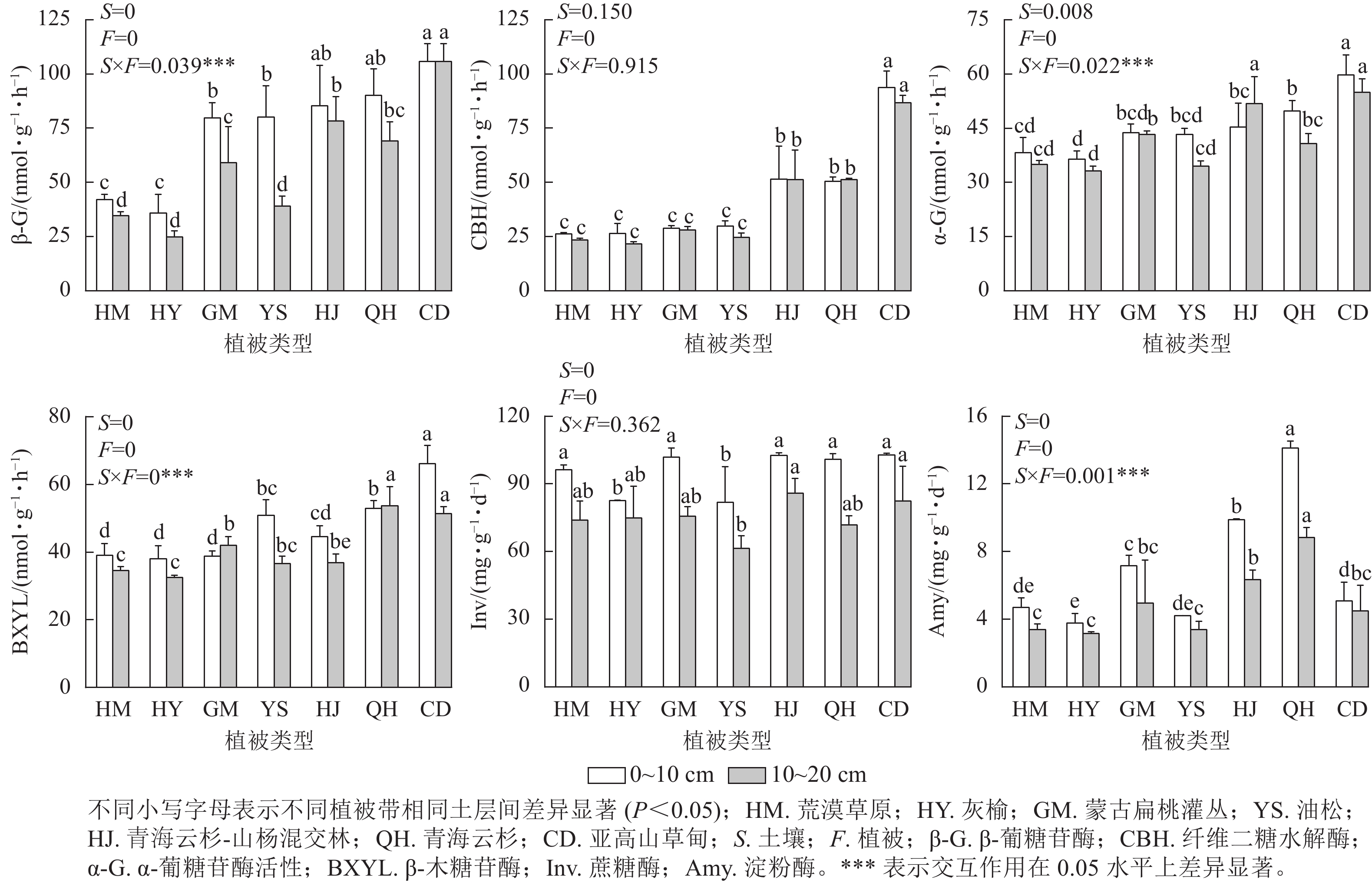
从图2可以看出,在不同海拔植被带中,β-葡糖苷酶活性在0~10 cm土层表现为随海拔升高先下降后上升,亚高山草甸酶活性显著高于其他植被带,为105.81 nmol·g−1·h−1;纤维二糖水解酶在0~10和10~20 cm土层中随海拔上升酶活性升高,在不同土层间酶活性无显著差异性,且在0~10与10~20 cm土层中其酶活性均在草甸处最高,分别为93.77与86.79 nmol·g−1·h−1;在0~10 cm土层中α-葡糖苷酶活性和β-木糖苷酶活性在亚高山草甸最高,分别为59.75、66.08 nmol·g−1·h−1,灰榆林最低,分别为36.41、38.03 nmol·g−1·h−1。在0~10与10~20 cm土层中蔗糖酶活性在油松林最低,分别为81.87、61.33 nmol·g−1·h−1;淀粉酶活性在不同土层以青海云杉林最高,分别为14.13、8.82 nmol·g−1·h−1,灰榆林最低,分别为3.78、3.17 nmol·g−1·h−1。双因素方差分析表明:土层与植被类型的交互作用对土壤β-葡糖苷酶、α-葡糖苷酶、β-木糖苷酶和淀粉酶活性产生显著影响,在不同海拔植被带0~10与10~20 cm土层,土壤β-葡糖苷酶、纤维二糖水解酶、α-葡糖苷酶、β-木糖苷酶随海拔上升整体上升,淀粉酶先下降后上升,蔗糖酶变化无显著规律。
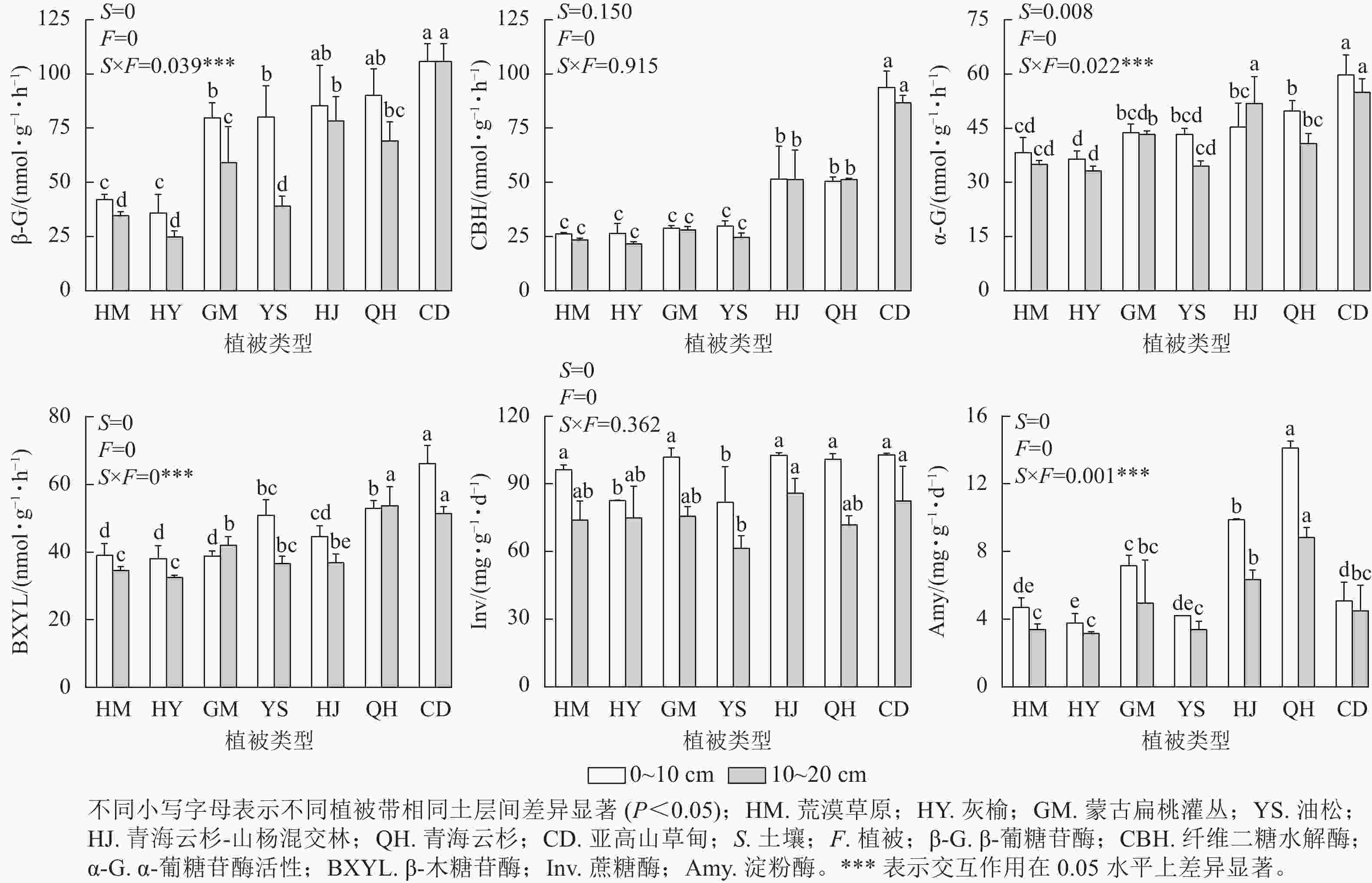
图 2 不同植被类型土壤酶活性
Figure 2. Soil enzyme activities in different vegetation types
-
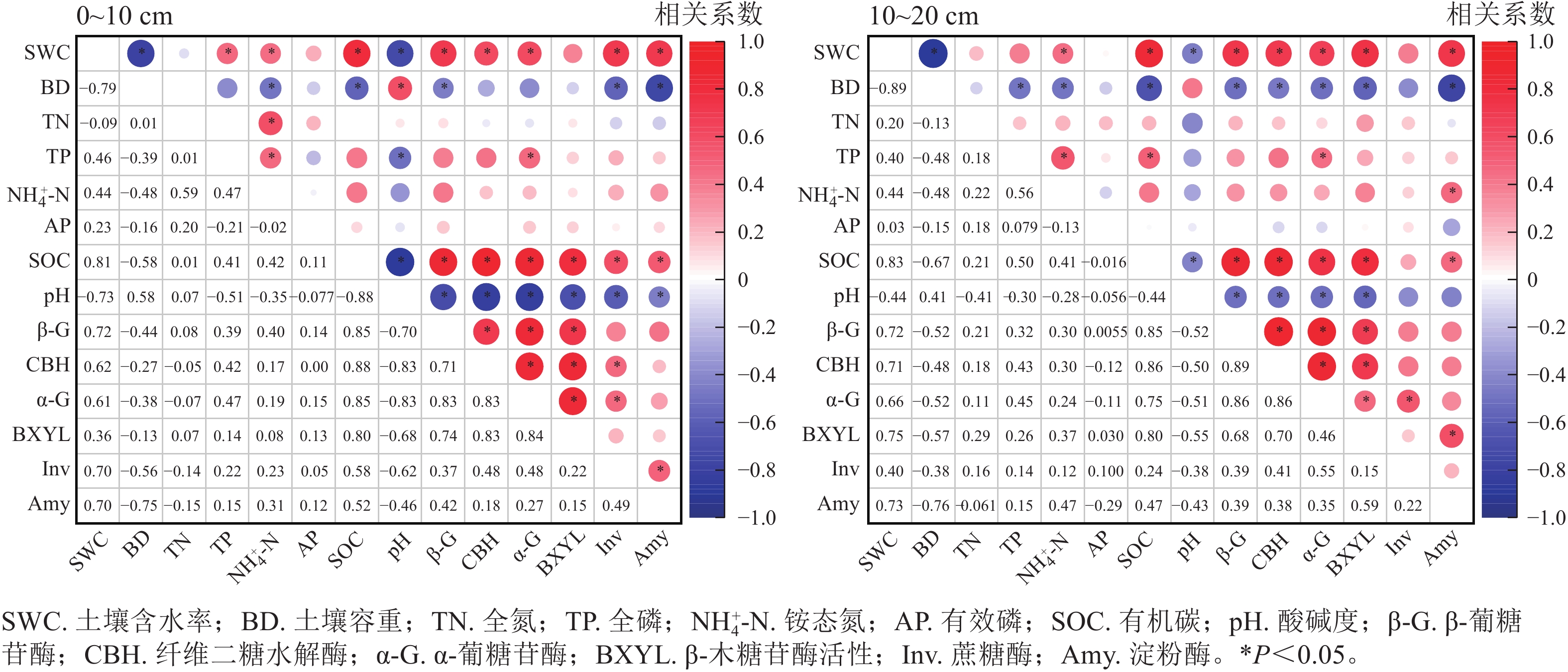
相关性分析如图3所示:在不同植被类型0~10 cm土层中,含水率、有机碳质量分数与各酶活性呈显著正相关(P<0.05),而容重、pH与各酶活性呈负相关,全氮、全磷、有效磷及铵态氮质量分数对各酶活性的影响不显著。在10~20 cm土层中,含水率、有机碳质量分数对土壤各酶活性的影响呈正相关(P<0.05),容重、pH对各酶活性的影响呈负相关,全氮、全磷、有效磷、铵态氮质量分数对各酶活性的影响并不显著。
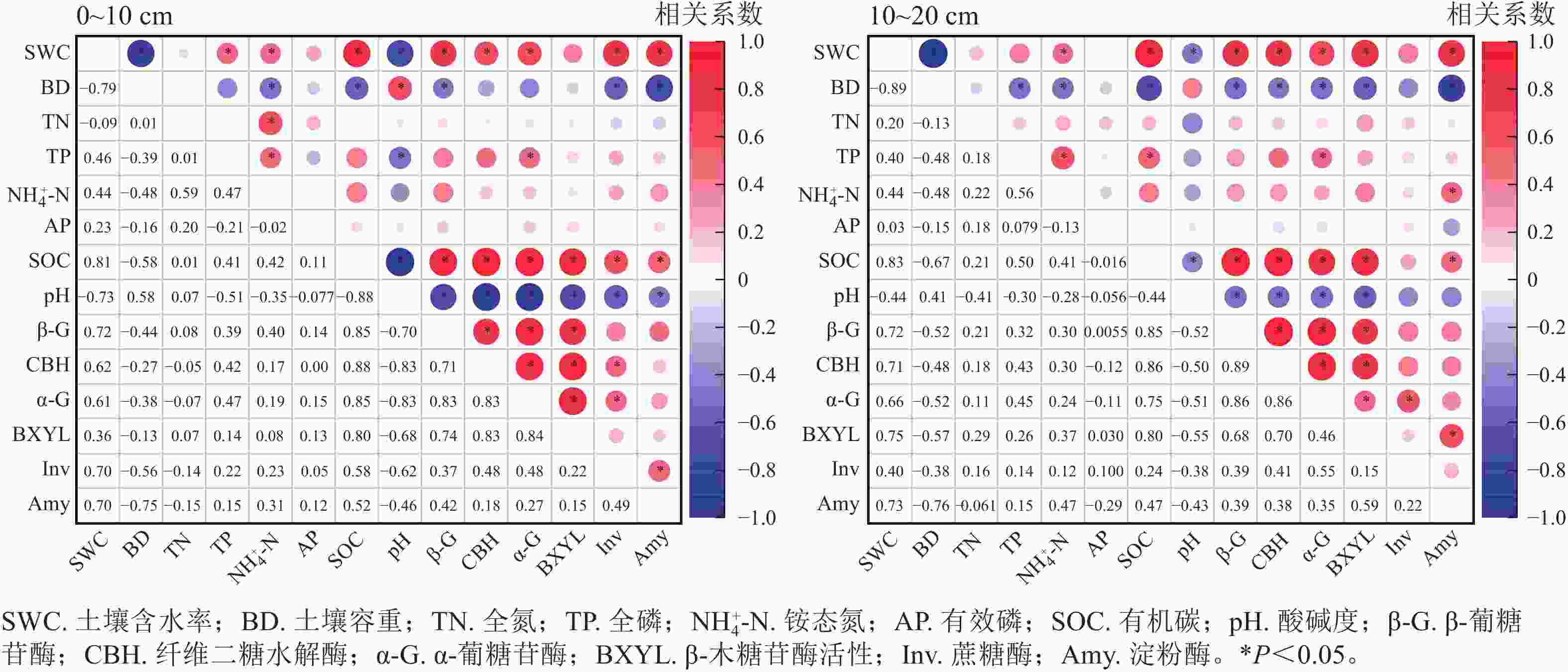
图 3 不同土层土壤理化性质与土壤酶活性的相关性分析
Figure 3. Correlation analysis between soil physicochemical properties and soil enzyme activity in different soil layers
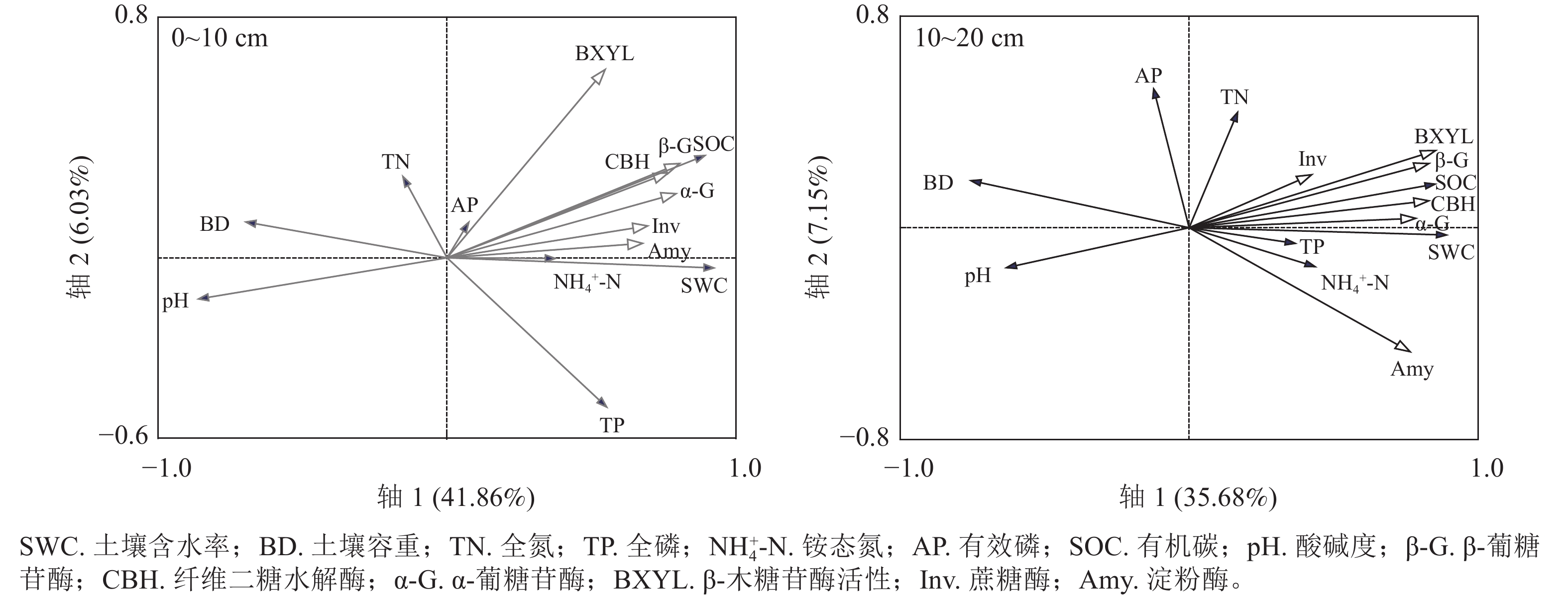
不同海拔土壤酶活性与理化性质的冗余分析(图4)显示:在不同植被类型,0~10 cm土层中,土壤理化性质对土壤酶活性影响重要性由大到小为有机碳质量分数、pH、含水率、容重、全磷质量分数、铵态氮质量分数、有效磷质量分数、全氮质量分数。其中有机碳质量分数、pH、含水率对土壤酶活性的影响达显著水平,而其他理化性质对土壤酶活性的影响并没有达显著水平。10~20 cm土层中,各酶活性与有机碳质量分数、铵态氮质量分数、含水率、全氮质量分数及全磷质量分数均表现为夹角小且方向一致,呈显著正相关,与容重、pH及有效磷质量分数呈负相关。在10~20 cm土层中,土壤有机碳、含水率和pH对土壤酶活性的影响呈显著水平,但其他理化性质对土壤酶活性的影响并没有达显著水平(表1)。
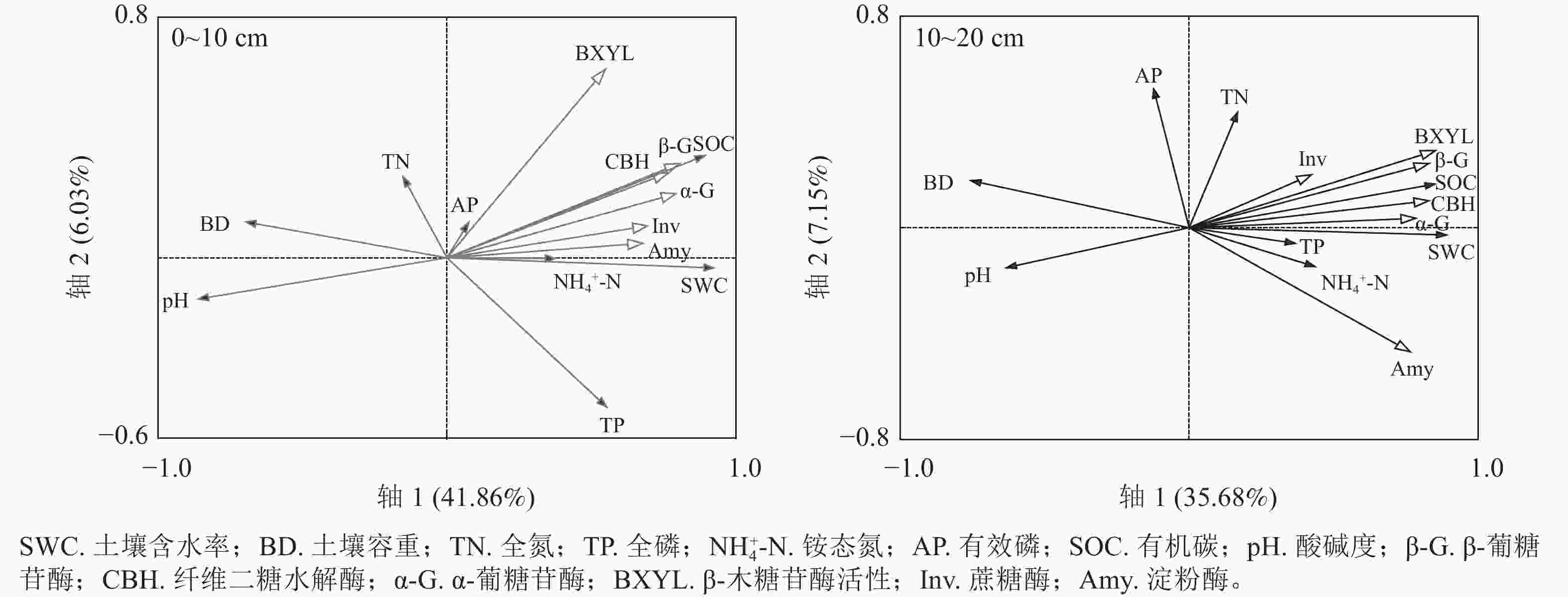
图 4 不同土层土壤酶活性与土壤理化性质的冗余分析(RDA)
Figure 4. Redundancy analysis (RDA) of soil enzyme activity and soil physicochemical properties in different soil layers
表 1 不同土层土壤理化性质对土壤酶活性的贡献率
Table 1. Contribution rate of soil physicochemical properties to soil enzyme activity in different soil layers
理化性质 0~10 cm 理化性质 10~20 cm 贡献率/% F P 贡献率/% F P 有机碳 85.1 44.9 0.002 有机碳 87.9 37.8 0.002 pH 67.8 24.2 0.002 含水率 64.0 17.8 0.002 含水率 52.4 14.5 0.002 pH 33.6 6.5 0.010 容重 18.0 3.3 0.066 容重 32.9 6.3 0.016 全磷 17.8 3.3 0.056 全磷 16.5 2.7 0.138 铵态氮 10.8 1.9 0.168 铵态氮 11.5 1.8 0.206 有效磷 1.4 0.2 0.828 全氮 5.1 0.8 0.412 全氮 0.7 0.1 0.926 有效磷 0.8 0.1 0.906 -
本研究区由于受海拔植被类型影响,0~10与10~20 cm土层的土壤含水率从低海拔到高海拔呈上升趋势。原因在于高海拔区域乔木林有较多植被,其覆盖率较大,树木的根系能较好地保持土壤水分,减少水分的蒸发与流失,这与马剑等[16]的结论一致。本研究结果显示:不同海拔植被类型土壤容重整体均随海拔升高而下降,其原因可能是贺兰山西坡乔木林及亚高山草甸植被在高海拔区域,土壤疏松,土壤腐殖质质量分数也高,且人类活动较少。张珊等[17]则研究认为亚高山不同海拔、不同土层土壤孔隙度和有机质质量分数不同导致了土壤容重分布规律不同。
本研究中,土壤全氮质量分数在不同植被带0~10 cm土层中未表现较大差异,说明贺兰山土壤受其他影响因子亚高山的影响大于植被类型的变化。李彦娇等[18]对内乡宝天曼自然保护区土壤研究发现:土壤全氮随海拔上升其质量分数增加,是由于海拔的上升导致温度下降,微生物活动减少,以及植物残体的分解,因而增加了土壤中全氮的积累,这与本研究不符。在本研究中贺兰山土壤有机碳质量分数在乔木林与亚高山草甸处较高。这可能是草甸植物不断更替,植物死亡所释放的二氧化碳低于其更新速度,同时高海拔地区降水多温度低,限制了凋落物和根系等的分解,有利于有机碳的积累;同时高海拔地区乔木林土壤有较多养分能够为乔木林提供适宜的生长环境,促进有机物的分解和转化,有利于土壤有机碳的积累。宁朋等[19]研究也认为:在高海拔地区,低温不利于土壤微生物生存,土壤的呼吸代谢作用减弱,有利于土壤有机碳的积累。另外,不同海拔的植被类型各不相同,导致土壤中所残留的凋落物特性存在显著差异,从而改变了土壤有机碳质量分数。在本研究中pH在不同植被类型0~10 cm土层中无显著差异,其原因可能为不同海拔植被类型与土壤具有相互作用,同时对pH的差异具有调节作用,这与在藏东南森林研究的结果一致[20]。速效磷质量分数在本研究中无显著变化规律,可能是由于土壤中的磷与土壤颗粒表面的铁、铝等离子结合,形成难溶性的磷酸盐。在干旱区碱性土壤中,这种吸附作用可能更加显著,导致磷的有效性降低。马剑等[16]研究则认为是由于随海拔上升磷素在土壤中迁移速度较慢,降水对磷素在土壤剖面及表面的迁移影响较弱,致使在不同海拔植被带,土壤速效磷无显著变化规律。
-
蔗糖酶活性不仅决定了土壤中的生物活性,也决定了植物对可溶性营养物质的利用能力[21]。本研究中,蔗糖酶活性在不同植被类型土壤表层无显著差异,但随土层加深呈降低趋势,这与秦燕等[22]、陈志芳等[23]的研究结果一致。可能是随土层加深,土壤密度及空隙度和通气性减小,有机质及养分减少,从而限制了微生物的生长与代谢,进而降低了蔗糖酶的活性。本研究表层土壤中β-葡糖苷酶、纤维二糖水解酶、α-葡糖苷酶活性均有上升趋势,这与李丹丹[24]、姚兰等[25]所得结论一致。表明高海拔植被更多的活性碳可能被土壤微生物分解,同时更有利于土壤养分积累,表层土壤通气及水分保持效能较好,有利于土壤微生物的生长和活动,因而高海拔地区酶活性较高。贺兰山西坡不同植被类型土壤酶活性多为0~10 cm高于10~20 cm土层。可能因为土壤表层的水热条件和通风条件好,腐殖质层养分含量高;同时表层土壤容重随植被根系分布增多而变小,从而促进土壤代谢产酶能力[26]。
-
本研究不同植被类型0~10与10~20 cm土层中土壤酶活性与含水率均表现为正相关关系,说明土壤水分有利于各种酶促反应,促进凋落物分解和高分子化合物的形成和积累[27]。在不同植被类型0~10与10~20 cm土层中土壤容重与所有酶活性之间存在着显著负相关,其主要原因为高容重土壤限制了氧气与水分的供应,这与李聪等[28]的研究结果相似。不同植被类型0~10与10~20 cm土层中,β-葡糖苷酶活性与土壤有机碳质量分数呈极显著正相关,与土壤 pH 呈极显著负相关,这与在猫儿山研究结果一致[29]。因为在碱性土壤中,酶的结构和功能可能会发生改变,从而影响其催化活性,同时酶与底物之间的亲和力降低,影响了酶的催化效率。本研究蔗糖酶与pH呈负相关,这与前人得出结论不同[30],这可能是因为土壤蔗糖酶活性与有机质可以相互促进,协同变化。pH可以影响酶的结构从而影响酶活性[31−32],蔗糖酶活性最适pH范围为6.5~7.5 [33−34]。本研究仅分析了不同植被类型的土壤酶活性和土壤理化性质的分布特征及相关关系,但土壤酶是一个复合体,土壤理化性质和水热条件的差异都会导致土壤酶活性的变化,因此,在今后的研究中,应综合考虑环境因素对土壤酶活性的影响,同时加强对土壤酶与土壤有机质[35]、土壤质地关系[36]的研究,结合研究区特点,全面深入调查其对环境因子变化的响应。
-
在贺兰山西坡不同植被类型中,土壤β-葡糖苷酶、纤维二糖水解酶、β-木糖苷酶及α-葡糖苷酶活性在0~10 cm土层随海拔升高整体呈上升趋势,在10~20 cm土层中均呈先下降后上升的趋势,而蔗糖酶与淀粉酶活性无显著变化规律,同时在不同植被类型不同土层中,土壤表层酶活性质量分数高于10~20 cm土层中的酶活性。0~10和10~20 cm土层中土壤有机碳质量分数和含水率均随海拔升高呈上升趋势,对酶活性具有促进作用,而容重与pH随海拔上升呈下降趋势,对酶活性具有抑制作用。
Characteristics of soil physicochemical properties and enzyme activities in different vegetation types on the western slope of Helan Mountain
-
摘要:
目的 探讨贺兰山西坡不同植被类型土壤理化性质对土壤酶活性的影响,为干旱区森林生态系统土壤酶活性变化、养分循环模式和调节机制研究提供参考依据。 方法 以贺兰山西坡土壤为研究对象,按照海拔梯度依次选择荒漠草原(海拔1 349 m)、灰榆Ulmus glaucescens林(1 905 m)、蒙古扁桃Amygdalus mongolica灌丛(2 134 m)、油松Pinus tabuliformis林(2 150 m)、青海云杉Picea crassifolia-山杨Populus davidiana混交林(2 160 m)、青海云杉林(2 635 m)和亚高山草甸(2 664 m)等7种典型植被类型。每个植被类型设置3个重复样地,通过野外调查取样与室内分析相结合的方法,对土壤理化性质(含水率、容重、pH、有机碳、铵态氮、全氮、全磷、有效磷)和酶活性(β-葡糖苷酶、纤维二糖水解酶、α-葡糖苷酶、β-木糖苷酶、蔗糖酶、淀粉酶)进行测定,利用方差分析和冗余分析探明不同植被类型土壤酶活性质量分数差异,并分析其影响要素。 结果 0~10和10~20 cm土层土壤含水率、有机碳质量分数随海拔上升整体呈现上升趋势;而土壤容重随海拔上升在0~10与10~20 cm土层整体呈现下降趋势;土壤全磷、全氮、铵态氮和有效磷质量分数以及pH在0~10和10~20 cm土层中随海拔上升无显著变化规律;在不同海拔植被带0~10 cm土层土壤各酶活性高于10~20 cm土层;土壤纤维二糖水解酶与β-葡糖苷酶随海拔升高在0~10 cm土层呈上升趋势,淀粉酶活性随海拔升高在0~10 cm土层呈先升高后降低再升高趋势,而α-葡糖苷酶、β-木糖苷酶和蔗糖酶活性在不同植被带不同土层之间均变化幅度较小且差异不显著;在0~10与10~20 cm土层,土壤有机碳、全磷、铵态氮、含水率是土壤酶活性的主要影响因素。 结论 海拔梯度变化引起贺兰山西坡土壤理化性质显著变化,进而调控森林土壤酶活性沿海拔及土层分布的变化,其中土壤有机碳、pH及含水率是土壤酶活性的主要影响因素,能较好地解释土壤酶活性的海拔梯度差异。图4表1参36 Abstract:Objective The aim is to investigate the effects of soil physicochemical properties in different vegetation types on soil enzyme activity on the western slope of Helan Mountain, and to provide a reference basis for the changes of soil enzyme activity, nutrient cycling patterns and regulatory mechanisms in forest ecosystems in dry areas. Method The seven vegetation types on the western slope of Helan Mountain were selected as the research object, which were desert grassland, Ulmus glaucescens forest, Amygdalus mongolica shrub, Pinus tabuliformis forest, Picea crassifolia-Populus davidiana forest and Picea crassifolia forest and subalpine meadow respectively from 1 349 to 2 664 m. Three replicate plots were set up for each vegetation type, and soil physicochemical properties (water content, bulk density, pH, organic carbon, ammonium nitrogen, total nitrogen, total phosphorus, and effective phosphorus) and enzyme activities (β-glucosidase, cellobiose hydrolase, α-glucosidase, β-xylosidase, sucrase, and amylase) were measured by field survey sampling and indoor analysis, with ANOVA and redundancy analysis. Result Soil water content and organic carbon mass fraction in the 0 − 10 and 10 − 20 cm soil layers of different vegetation zones at different altitudes showed an overall increasing trend with the rise in altitude; while soil bulk weight showed a decreasing trend with the rise in altitude in the 0 − 10 and 10 − 20 cm soil layers as a whole; soil total phosphorus, total nitrogen, ammonium nitrogen, effective phosphorus, and pH did not show any significant change patterns in the 0 − 10 and 10 − 20 cm soil layers with the rise in altitude; soil enzyme activities in the 0 − 10 cm soil layer were higher than those in the 10 − 20 cm soil layer. The activities of soil enzymes were higher in the 0 − 10 cm soil layer than in the 10 − 20 cm soil layer in different vegetation zones; Soil cellobiose hydrolase and β-glucosidase showed an increasing trend with elevation in the 0 − 10 cm soil layer, amylase activity showed an increasing trend with elevation in the 0 − 10 cm soil layer, while α-glucosidase, β-xylosidase, and sucrase activities showed small and non-significant variations in different soil layers in different vegetation zones; Soil organic carbon, total phosphorus, ammonium nitrogen, and water content were the main factors affecting soil enzyme activities in the 0 − 10 and 10 − 20 cm soil layers. In the 0 − 10 and 10 − 20 cm soil layers, soil organic carbon, total phosphorus, ammonium nitrogen, and water content were the main influencing factors of soil enzyme activities. Conclusion Changes in altitudinal gradient caused significant changes in soil physicochemical properties on the western slope of Helan Mountain, which in turn regulated the changes in forest soil enzyme activity along the distribution of altitude and soil layers. Soil organic carbon, pH and water content were the main factors influencing soil enzyme activity, and could better explain the differences in the altitudinal gradient of soil enzyme activity. [Ch, 4 fig. 1 tab. 36 ref.] -
Key words:
- Helan Mountain /
- vegetation type /
- soil enzyme activity /
- physicochemical properties
-
图 1 不同植被类型土壤基本理化性质
Figure 1 Basic physical and chemical properties of soils at different vegetation types
图 3 不同土层土壤理化性质与土壤酶活性的相关性分析
Figure 3 Correlation analysis between soil physicochemical properties and soil enzyme activity in different soil layers
图 4 不同土层土壤酶活性与土壤理化性质的冗余分析(RDA)
Figure 4 Redundancy analysis (RDA) of soil enzyme activity and soil physicochemical properties in different soil layers
表 1 不同土层土壤理化性质对土壤酶活性的贡献率
Table 1. Contribution rate of soil physicochemical properties to soil enzyme activity in different soil layers
理化性质 0~10 cm 理化性质 10~20 cm 贡献率/% F P 贡献率/% F P 有机碳 85.1 44.9 0.002 有机碳 87.9 37.8 0.002 pH 67.8 24.2 0.002 含水率 64.0 17.8 0.002 含水率 52.4 14.5 0.002 pH 33.6 6.5 0.010 容重 18.0 3.3 0.066 容重 32.9 6.3 0.016 全磷 17.8 3.3 0.056 全磷 16.5 2.7 0.138 铵态氮 10.8 1.9 0.168 铵态氮 11.5 1.8 0.206 有效磷 1.4 0.2 0.828 全氮 5.1 0.8 0.412 全氮 0.7 0.1 0.926 有效磷 0.8 0.1 0.906  下载: 导出CSV
下载: 导出CSV
-
[1] 曹瑞, 吴福忠, 杨万勤, 等. 海拔对高山峡谷区土壤微生物生物量和酶活性的影响[J]. 应用生态学报, 2016, 27(4): 1257 − 1264. CAO Rui, WU Fuzhong, YANG Wanqin, et al. Effects of altitude on soil microbial biomass and enzyme activities in alpine-gorge regions [J]. Chinese Journal of Applied Ecology, 2016, 27(4): 1257 − 1264. [2] 佟小刚. 不同基质和施肥类型对无土栽培莴苣生长及基质中微生物和酶的影响 [D]. 杨凌: 西北农林科技大学, 2005. TONG Xiaogang. The Effects of Different Substrates and Fertilizer on the Growth of Lettuce and the Microorganism and Enzyme in Soilless Culture [D]. Yangling: Northwest A&F University, 2005. [3] MA Heping, YANG Xiaolin, GUO Qiqiang, et al. Soil organic carbon pool along different altitudinal level in the Sygera Mountains, Tibetan Plateau [J]. Journal of Mountain Science, 2016, 13(3): 476 − 483. [4] 谷晓楠, 贺红士, 陶岩, 等. 长白山土壤微生物群落结构及酶活性随海拔的分布特征与影响因子[J]. 生态学报, 2017, 37(24): 8374 − 8384. GU Xiaonan, HE Hongshi, TAO Yan, et al. Soil microbial community structure, enzyme activities, and their influencing factors along different altitudes of Changbai Mountain [J]. Acta Ecologica Sinica, 2017, 37(24): 8374 − 8384. [5] LI Zhijie, TAYLOR J E, LOU Chenglin. An unusual Lophodermium species on needles of Pinus taiwanensis from China [J]. Micrologica Progress, 2016, 15: 1229 − 1237. [6] 金裕华, 汪家社, 李黎光, 等. 武夷山不同海拔典型植被带土壤酶活性特征[J]. 生态学杂志, 2011, 30(9): 1955 − 1961. JIN Yuhua, WANG Jiashe, LI Liguang, et al. Soil enzyme activities in typical vegetation zones along an altitude gradient in Wuyi Mountains [J]. Chinese Journal of Ecology, 2011, 30(9): 1955 − 1961. [7] 彭子洋, 刘卫星, 田瑞, 等. 海拔和坡向对唐古拉山土壤胞外酶活性的影响[J]. 生态学报, 2021, 41(19): 7659 − 7668. PENG Ziyang, LIU Weixing, TIAN Rui, et al. Effects of altitude and aspect on soil extracellular enzyme activities in Tanggula Mountain [J]. Acta Ecologica Sinica, 2021, 41(19): 7659 − 7668. [8] 赵盼盼, 周嘉聪, 林开淼, 等. 不同海拔对福建戴云山黄山松林土壤微生物生物量和土壤酶活性的影响[J]. 生态学报, 2019, 39(8): 2676 − 2686. ZHAO Panpan, ZHOU Jiacong, LIN Kaimiao, et al. Effects of different altitudes on soil microbial biomass and enzyme activities in Pinus taiwanensis forests on Daiyun Mountain, Fujian Province [J]. Acta Ecologica Sinica, 2019, 39(8): 2676 − 2686. [9] 王晶晶. 热带和亚热带不同森林类型土壤胞外酶活性及计量关系的纬度分异规律及驱动因素[D]. 福州: 福建师范大学, 2022. WANG Jingjing. Latitudinal Variation and Driving Factors of Soil Extracellular Enzymes and Stoichiometry in Different Tropical and Subtropical Forest Types [D]. Fuzhou: Fujian Normal University, 2022. [10] 隋夕然, 吴丽芳, 王妍, 等. 滇中岩溶高原不同石漠化程度土壤团聚体养分及酶活性特征[J]. 浙江农林大学学报, 2022, 39(1): 115 − 126. SUI Xiran, WU Lifang, WANG Yan, et al. Characteristics of nutrient and enzyme activity in soil aggregates of different rocky desertification levels in central Yunnan Plateau [J]. Journal of Zhejiang A&F University, 2022, 39(1): 115 − 126. [11] 刘顺, 许格希, 陈淼, 等. 坡向对川西亚高山土壤酶活性和微生物养分限制的影响[J]. 应用生态学报, 2023, 34(11): 2993 − 3002. LIU Shun, XU Gexi, CHEN Miao, et al. Effects of slope aspect on soil enzyme activity and microbial nutrient limitation in subalpine region of western Sichuan, China [J]. Chinese Journal of Applied Ecology, 2023, 34(11): 2993 − 3002. [12] 庞丹波, 吴梦瑶, 吴旭东, 等. 贺兰山东坡不同海拔梯度土壤酶化学计量特征[J]. 生态学报, 2023, 43(19): 7950 − 7962. PANG Danbo, WU Mengyao, WU Xudong, et al. Responses of soil enzyme activities and their stoichiometric characteristics to different altitude on the eastern slope of Helan Mountain [J]. Acta Ecologica Sinica, 2023, 43(19): 7950 − 7962. [13] 万红云, 陈林, 庞丹波, 等. 贺兰山不同海拔土壤酶活性及其化学计量特征[J]. 应用生态学报, 2021, 32(9): 3045 − 3052. WAN Hongyun, CHEN Lin, PANG Danbo, et al. Soil enzyme activities and their stoichiometry at different altitudes in Helan Mountains, Northwest China [J]. Chinese Journal of Applied Ecology, 2021, 32(9): 3045 − 3052. [14] 张振, 梁海彬, 陈有超, 等. 模拟家畜不同采食强度下高寒草甸土壤水解酶活性的变化特征[J]. 浙江农林大学学报, 2023, 40(2): 265 − 273. ZHANG Zhen, LIANG Haibin, CHEN Youchao, et al. Responses of soil enzyme activities to different defoliation intensities in alpine meadow [J]. Journal of Zhejiang A&F University, 2023, 40(2): 265 − 273. [15] 张珊. 不同林龄序列云杉人工林土壤化学计量特征及其与土壤因子的相关性研究[D]. 兰州: 甘肃农业大学, 2017. ZHANG Shan. Study on the Relationship between Soil Ecological Stoichiometry of Spruce Plantation in Different Forest Age and Soil Factor [D]. Lanzhou: Gansu Agricultural University, 2017. [16] 马剑, 刘贤德, 金铭, 等. 祁连山青海云杉林土壤理化性质和酶活性海拔分布特征[J]. 水土保持学报, 2019, 33(2): 207 − 213. MA Jian, LIU Xiande, JIN Ming, et al. Soil physicochemical properties and enzyme activities along the altitudinal gradients in Picea crassifolia of Qilian Mountains [J]. Journal of Soil and Water Conservation, 2019, 33(2): 207 − 213. [17] 张珊, 田晓娟, 顾振东, 等. 甘肃亚高山不同海拔梯度云杉人工林土壤理化性质研究[J]. 甘肃农业大学学报, 2021, 56(6): 111 − 118. ZHANG Shan, TIAN Xiaojuan, GU Zhendong, et al. Study on soil physical and chemical properties in the artificial forest of spruce plantation along altitudinal gradients in the subalpine region of Gansu Province [J]. Journal of Gansu Agricultural University, 2021, 56(6): 111 − 118. [18] 李彦娇, 赵燕, 王立民, 等. 内乡宝天曼自然保护区土壤理化性质和酶活性的海拔特征研究[J]. 信阳师范学院学报(自然科学版), 2016, 29(4): 560 − 566. LI Yanjiao, ZHAO Yan, WANG Limin, et al. A study of soil physicochemical property and soil enzyme along altitude gradient at Neixiang Baotianman Nature Reserve [J]. Journal of Xinyang Normal University (Natural Science Edition), 2016, 29(4): 560 − 566. [19] 宁朋, 程小毛, 杨晓锋, 等. 贡嘎山不同海拔峨眉冷杉根际土壤化学性质及胞外酶活性[J]. 应用与环境生物学报, 2021, 27(5): 1138 − 1146. NING Peng, CHENG Xiaomao, YANG Xiaofeng, et al. Chemical properties and extracellular enzymatic activity in the rhizosphere soil of Abies fabri at different altitudes on Mount Gongga [J]. Journal of Applied and Environmental Biology, 2021, 27(5): 1138 − 1146. [20] 斯贵才, 袁艳丽, 王建, 等. 藏东南森林土壤微生物群落结构与土壤酶活性随海拔梯度的变化[J]. 微生物学通报, 2014, 41(10): 2001 − 2011. SI Guicai, YUAN Yanli, WANG Jian, et al. Microbial community and soil enzyme activities along an altitudinal gradient in Sejila Mountains [J]. Microbiology China, 2014, 41(10): 2001 − 2011. [21] 张仕艳, 原海红, 陆梅, 等. 滇西北不同利用类型土壤酶活性及其与理化性质与微生物的关系[J]. 亚热带水土保持, 2010, 22(2): 13 − 16. ZHANG Shiyan, YUAN Haihong, LU Mei, et al. The soil enzyme activities of different land use types and the relationships between the soil enzyme activities and physical-chemical properties or microorganism in mountainous area of northwest Yunnan Province [J]. Subtropical Soil and Water Conservation, 2010, 22(2): 13 − 16. [22] 秦燕, 牛得草, 康健, 等. 贺兰山西坡不同类型草地土壤酶活性特征[J]. 干旱区研究, 2012, 29(5): 870 − 877. QIN Yan, NIU Decao, KANG Jian, et al. Characteristics of soil enzyme activities in different grasslands in the western slope of the Helan Mountain, China [J]. Arid Zone Research, 2012, 29(5): 870 − 877. [23] 陈志芳, 刘金福, 吴则焰. 不同海拔土壤理化性质与酶活性研究[J]. 河北北方学院学报(自然科学版), 2014, 30(1): 38 − 42. CHEN Zhifang, LIU Jinfu, WU Zeyan. Soil physicochemical properties and enzyme activities at different elevation gradients [J]. Journal of Hebei North University (Natural Science Edition), 2014, 30(1): 38 − 42. [24] 李丹丹. 北方温带森林不同海拔梯度土壤有机质分解酶活性及动力学特征研究[D]. 沈阳: 沈阳农业大学, 2016. LI Dandan. Characterisation of Enzyme Activities and Kinetics of Soil Organic Matter Decomposition at Different Altitude Gradients in a Boreal Temperate Forest [D]. Shenyang: Shenyang Agricultural University, 2016. [25] 姚兰, 张焕朝, 胡立煌, 等. 黄山不同海拔植被带土壤活性有机碳、氮及其与酶活性的关系[J]. 浙江农林大学学报, 2019, 36(6): 1069 − 1076. YAO Lan, ZHANG Huanzhao, HU Lihuang, et al. Soil labile organic carbon and nitrogen and their relationship with enzyme activities in different vegetation zones along an altitudinal gradient on Mount Huangshan [J]. Journal of Zhejiang A&F University, 2019, 36(6): 1069 − 1076. [26] 曹婷婷, 郭振. 森林土壤酶活性与土壤质量相关性探究[J]. 生物化工, 2019, 5(6): 144 − 146. CAO Tingting, GUO Zhen. Study on correlation between forest soil enzyme activity and soil quality [J]. Biological Chemical Engineering, 2019, 5(6): 144 − 146. [27] 张莎莎, 李爱琴, 王会荣, 等. 不同海拔杉木人工林土壤碳氮磷生态化学计量特征[J]. 生态环境学报, 2020, 29(1): 97 − 104. ZHANG Shasha, LI Aiqin, WANG Huirong, et al. Ecological stoichiometry of soil carbon, nitrogen and phosphorus in Cunninghamia lanceolata plantation across an elevation gradient [J]. Ecology and Environmental Sciences, 2020, 29(1): 97 − 104. [28] 李聪, 吕晶花, 陆梅, 等. 滇东南典型常绿阔叶林土壤酶活性的海拔梯度特征[J]. 林业科学研究, 2020, 33(6): 170 − 179. LI Cong, LÜ Jinghua, LU Mei, et al. Variations of soil enzyme activity in typical evergreen broadleaved forests along altitude gradient in Southeast Yunnan [J]. Forest Research, 2020, 33(6): 170 − 179. [29] 宋贤冲, 陈晓鸣, 郭丽梅, 等. 猫儿山不同海拔典型植被带土壤酶活性变化特征[J]. 基因组学与应用生物学, 2016, 35(12): 3545 − 3551. SONG Xianchong, CHEN Xiaoming, GUO Limei, et al. Soil enzyme activities in typical vegetation zones along an altitude gradient in Mao’er Mountain [J]. Genomics and Applied Biology, 2016, 35(12): 3545 − 3551. [30] 陈双林, 郭子武, 杨清平. 毛竹林土壤酶活性变化的海拔效应[J]. 生态学杂志, 2010, 29(3): 529 − 533. CHEN Shuanglin, GUO Ziwu, YANG Qingping. Soil enzyme activities in moso bamboo forests along an altitude gradient [J]. Chinese Journal of Ecology, 2010, 29(3): 529 − 533. [31] 杨万勤, 王开运. 土壤酶研究动态与展望[J]. 应用与环境生物学报, 2002, 8(5): 564 − 570. YANG Wanqin, WANG Kaiyun. Advances on soil enzymology [J]. Journal of Applied and Environmental Biology, 2002, 8(5): 564 − 570. [32] 刘秉儒, 张秀珍, 胡天华, 等. 贺兰山不同海拔典型植被带土壤微生物多样性[J]. 生态学报, 2013, 33(22): 7211 − 7220. LIU Bingru, ZHANG Xiuzhen, HU Tianhua, et al. Soil microbial diversity under typical vegetation zones along an elevation gradient in Helan Mountains [J]. Acta Ecologica Sinica, 2013, 33(22): 7211 − 7220. [33] 马进鹏, 庞丹波, 陈林, 等. 贺兰山不同海拔植被下土壤微生物群落结构特征[J]. 生态学报, 2022, 42(2): 667 − 676. MA Jinpeng, PANG Danbo, CHEN Lin, et al. Characteristics of soil microbial community structure under vegetation at different altitudes in Helan Mountains [J]. Acta Ecologica Sinica, 2022, 42(2): 667 − 676. [34] 张猛, 张健. 林地土壤微生物、酶活性研究进展[J]. 四川农业大学学报, 2003, 21(4): 347 − 351. ZHANG Meng, ZHANG Jian. Advance in research on microbe and enzyme activity in forest soil [J]. Journal of Sichuan Agricultural University, 2003, 21(4): 347 − 351. [35] 孙海燕, 万书波, 李林, 等. 贺兰山西坡不同海拔梯度土壤活性有机碳分布特征及影响因子[J]. 水土保持学报, 2014, 28(4): 194 − 199, 205. SUN Haiyan, WAN Shubo, LI Lin, et al. Distribution characteristics and influencing factors of soil active organic carbon at different elevations on west slope of Helan Mountain [J]. Journal of Soil and Water Conservation, 2014, 28(4): 194 − 199, 205. [36] 肖好燕, 刘宝, 余再鹏, 等. 亚热带典型林分对表层和深层土壤可溶性有机碳、氮的影响[J]. 应用生态学报, 2016, 27(4): 1031 − 1038. XIAO Haoyan, LIU Bao, YU Zai peng, et al. Effects of forest types on soil dissolved organic carbon and nitrogen in surface and deep layers in subtropical region, China [J]. Chinese Journal of Applied Ecology, 2016, 27(4): 1031 − 1038. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20230485
 点击查看大图
点击查看大图
计量
- 文章访问数: 1357
- HTML全文浏览量: 266
- PDF下载量: 49
- 被引次数: 0
