




-
氨基酸、维生素C、粗纤维、粗蛋白质和可溶性糖是人体不可或缺的营养成分,参与体内重要的生命活动和代谢过程,而矿质元素是组成激素、维生素、蛋白质和多种酶的重要组成部分[1],在人体保健方面起着重要作用。竹笋作为中国的传统佳肴,已有2500多年的食用和栽培历史,其味道鲜美、口感清脆、营养丰富而深受人们喜爱,被誉为最受欢迎的五大保健食品之一。中国是世界上最大的竹笋生产国和消费国,每年有大量的竹笋及其相关产品远销海外。其中,毛竹Phyllostachys edulis笋产量居竹笋产量之首,含蛋白质类、纤维类、糖类、多种矿质元素和氨基酸等人体所需的营养物质,毛竹笋具有高纤维、高氮、高矿质、低脂肪、低糖等营养特点[2-4],符合现代社会人们对饮食健康和食品质量的追求,具有很好的市场前景。目前,竹笋品质的研究主要集中在营养动态[5-6]、存储包装方式[7-8]以及经营措施[9-10]对竹笋营养成分的影响。其中,大量研究表明:施肥能显著影响竹笋的产量[10-11]和笋体中的氨基酸、蛋白质和脂肪等营养物质的含量[9, 12]。近年来,随着工农业生产和人类活动强度的增加,工业活动的密集化,如化石燃料燃烧、氮肥施用、畜禽养殖等人为活动的剧增,致使氮排放量不断攀升,大气中活性氮持续升高,氮沉降已从发达地区迅速发展到全球范围,极大地干扰了氮循环[13]。中国的氮沉降情况愈加严重,仅次于欧洲和美国,为全球第三大氮沉降区[14]。中国亚热带地区的平均氮沉降量已达30.00 kg·hm−2·a−1[15],最大氮沉降量已达51.56 kg·hm−2·a−1,而且还有进一步增加的趋势[16]。中国亚热带地区也是毛竹林的主要分布区,毛竹林面积达443万hm2,约占全国竹林面积的70%,毛竹是该区域重要的森林资源和南方山区农民经济收入的重要来源[17]。目前研究表明:氮沉降对毛竹林的土壤酶活性[18]、毛竹叶片化学计量、养分重吸收及凋落叶养分归还[19]等方面都有显著影响,这些变化是否会影响竹笋营养成分含量,进而改变其品质,尚未可知。为此,本研究以毛竹笋为研究对象,通过人工施氮模拟大气氮沉降的方法,研究氮沉降对毛竹笋品质的影响,以期为在日益增强的大气氮沉降下毛竹笋可持续经营提供新的参考。
-
研究区位于浙江省杭州市临安区青山湖街道青山笋材两用毛竹林样地(30°14′N,119°42′E),该区处于中亚热带季风气候区的北缘,雨水充足,气候温润,四季分明。年平均降水量1 420 mm,年平均气温15.6 ℃,最高和最低气温分别为41.7 ℃和−13.3 ℃。年均日照约1 847 h,年均无霜期约230 d。土壤类型属于黄土壤,低山丘陵地貌,海拔100~300 m。研究地毛竹林采用集约经营模式,隔年采伐4 a以上的老竹,现存立竹主要是4龄竹(2016年)和2龄竹(2018年)。每年7−8月垦复除去林下的灌木和杂草,9月撒施450 kg·hm−2复合肥,施肥后翻耕30 cm,使之与表层土壤均匀混合。林地的林分和表层土壤(0~20 cm)初始特征见表1[20]。
表 1 毛竹林分和表层土壤(0~20 cm)特征
Table 1. Original characteristics of the Ph. edulis plantation and surface soil(0−20 cm)
处理 密度/
(株·hm−2)胸径/
cm土壤密度/
(g·cm−3)土壤有机碳/
(mg·g−1)土壤全氮/
(mg·g−1)土壤全磷/
(mg·g−1)土壤pH 对照(ck) 3 362±309 10.16±0.13 0.97±0.07 23.73±0.24 1.11±0.04 0.52±0.01 4.46±0.01 低氮(N30) 3 408±382 10.31±0.32 0.98±0.04 22.56±1.17 1.17±0.09 0.57±0.05 4.43±0.04 高氮(N60) 3 208±187 10.76±0.59 1.00±0.05 23.15±1.74 1.21±0.12 0.54±0.03 4.48±0.08 说明:数值为均值±标准差 -
采用典型样方法,在毛竹林内设立9个20 m×20 m典型样地,样地之间设置20 m以上的缓冲间隔区。以中国亚热带地区平均大气氮沉降量(30 kg·hm−2·a−1)和未来增加趋势为依据[21],设置3个氮沉降处理,分别为对照(0 kg·hm−2·a−1,ck)、低氮(30 kg·hm−2·a−1,N30)和高氮(60 kg·hm−2·a−1,N60),各处理重复3次。2013年1月至2019年4月,每月进行模拟氮沉降处理,全年12次。处理方法为在10 L水中加入各样地所需的相应质量的硝酸铵(NH4NO3),采用电动背式喷雾器将配好溶液均匀地喷洒在样地上;对照组采用同样的方法喷洒等质量的水,以减少外加水因素带来的误差影响。
-
于2019年4月在毛竹林样地采挖大小适中,出土高度约3~5 cm的毛竹笋,要求其整体形态特征明显、饱满健壮、笋体完整,每个样地各采集5个春笋,将笋箨上的泥土和杂物清理干净,装入放有冰袋的恒温采集箱,带回实验室用于分析检测。将新鲜春笋去除掉笋壳和竹篼,选取其中可食的鲜嫩部分切成块状,采用四分法取样,在(130±2) ℃的烘箱中杀青10 min后,放入(60±1) ℃干燥机中烘干,粉碎后过60目筛,放入干燥器中保存以供竹笋营养成分测定。
氨基酸采用氨基酸分析仪测定[9];维生素C用稀酸提取,高效液相色谱法测定[22];粗纤维的测定参照国标GB/T 5009.10−2003《植物类食品中粗纤维的测定》,采用酸碱洗涤法;粗蛋白质采用凯氏定氮法测定[23];可溶性糖采用硫酸苯酚法测定[24];矿质元素采用硝酸-高氯酸(HNO3-HClO4)湿法消化法,消化液用火焰原子吸收光谱法测定[25]。
-
采用单因素方差分析(one-way ANOVA)和最小显著差异法(LSD)分析不同氮沉降强度下毛竹笋中氨基酸、维生素C、粗纤维、粗蛋白质、可溶性糖和矿质元素差异;用SPSS 19.0进行统计分析;用Origin 2018作图。所有数据均为平均值±标准差。
-
由表2可知:氮沉降对毛竹鲜笋的总氨基酸、必需氨基酸和甜味氨基酸的质量分数均有显著提高作用(P<0.05),分别增加了45.03%~54.08%、33.66%~35.64%和96.50%~106.50%;而毛竹鲜笋的鲜味氨基酸、苦味氨基酸和芳香类氨基酸的质量分数分别在高氮下显著增加了20.43%、15.83%和21.21%(P<0.05),低氮下显著降低63.44%、11.15%和27.27%(P<0.05)。同时,不同氮沉降水平下,毛竹鲜笋的必需、鲜味、甜味、苦味和芳香类氨基酸比例变化较大,氮沉降对毛竹鲜笋中氨基酸比例具有显著影响(P<0.05)。其中甜味氨基酸比例在氮添加下显著增加34.02%~35.54%(P<0.05),而氮沉降对必需氨基酸、鲜味氨基酸、苦味氨基酸和芳香类氨基酸比例均有显著降低作用(P<0.05),分别降低7.84%~11.96%、21.92%~74.80%、24.79%~38.76%和21.30%~49.80%。
表 2 不同氮沉降水平下毛竹笋不同类型氨基酸质量分数及比例
Table 2. Contents and proportions of different types of amino acids under different nitrogen(N) deposition
氮处理 必需氨基酸 鲜味氨基酸 甜味氨基酸 苦味氨基酸 芳香类氨基酸 总氨基酸 质量分数/
(mg·g−1)比例/% 质量分数/
(mg·g−1)比例/% 质量分数/
(mg·g−1)比例/% 质量分数/
(mg·g−1)比例/% 质量分数/
(mg·g−1)比例/% 质量分数/
(mg·g−1)对照(ck) 4.04±0.16 Bb 22.57 0.93±0.03 Eb 5.20 2.00±0.10 Db 11.17 2.78±0.11 Cb 15.53 1.32±0.02 Eb 7.37 17.90±0.61 Ab 低氮(N30) 5.40±0.21 Ba 20.80 0.34±0.02 Ec 1.31 3.93±0.18 Ca 15.14 2.47±0.14 Dc 9.51 0.96±0.07 Ec 3.70 25.96±1.10 Aa 高氮(N60) 5.48±0.25 Ba 19.87 1.12±0.04 Da 4.06 4.13±0.14 Ca 14.97 3.22±0.35 Ca 11.68 1.60±0.08 Da 5.80 27.58±1.27 Aa 说明:同列不同小写字母表示不同氮沉降水平对各氨基酸影响差异显著(P<0.05),同行不同大写字母表示相同氮沉降水平对各氨基 酸影响差异显著(P<0.05) -
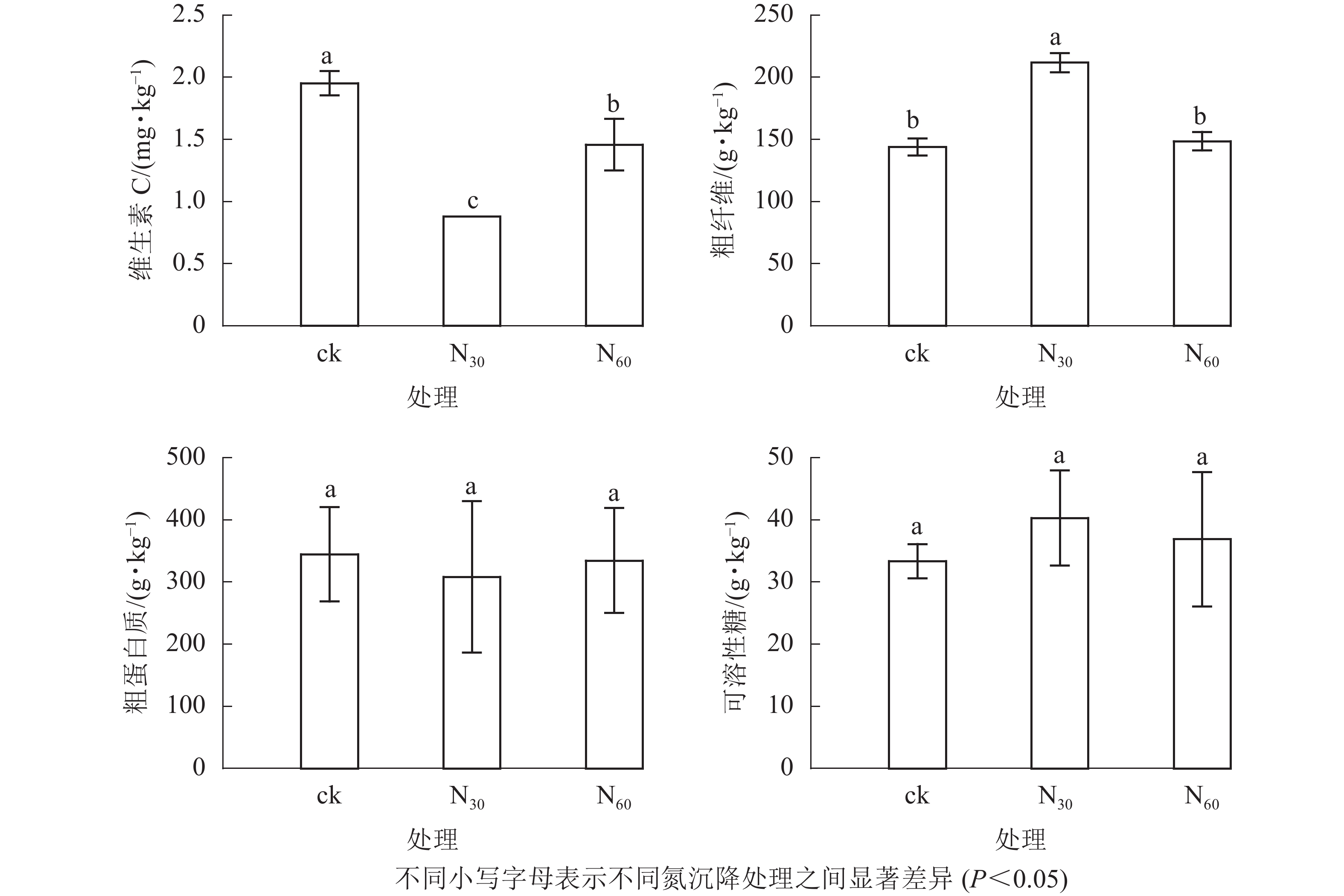
氮沉降对毛竹笋营养成分的影响显著(图1)。随氮沉降增加,维生素C和粗蛋白质质量分数均先下降而后升高,而粗纤维和可溶性糖质量分数则均呈先升高而下降趋势,且各处理毛竹笋的粗蛋白质、可溶性糖并无显著差异(P>0.05)。毛竹笋维生素C质量分数不同处理间则差异显著(P<0.05),而高氮沉降毛竹笋粗纤维质量分数与对照并无显著差异(P>0.05),且均显著高于低氮沉降处理(P<0.05)。

图 1 不同氮沉降水平下毛竹笋维生素C、粗纤维、粗蛋白质和可溶性糖质量分数
Figure 1. Contents of vitamin C, crude fiber, crude protein and soluble sugar under different nitrogen (N) deposition
-
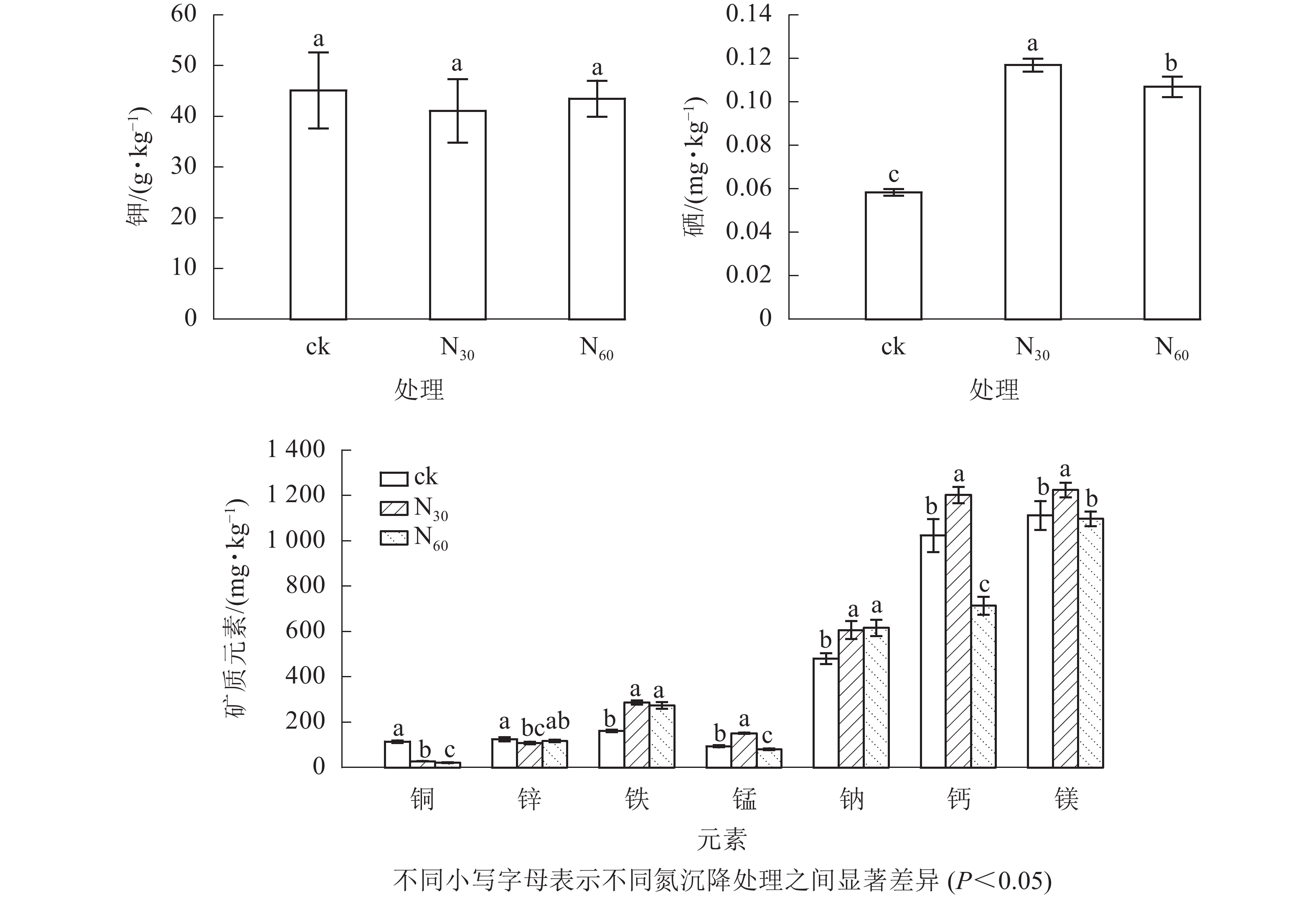
由图2可知:毛竹笋不同矿质元素质量分数差异较大,且对氮沉降的响应不同。其中:钾最高,硒最低。氮沉降对钾和锌无显著影响(P>0.05),但却显著提高了硒(83.15%~100.33%)、铁(69.57%~77.91%)和钠(26.22%~28.16%)(P<0.05)质量分数,显著降低了铜(18.19%~24.72%)(P<0.05)质量分数。同时,锰、钙和镁质量分数在低氮处理下显著增加(P<0.05),在高氮处理下显著降低(P<0.05)。与对照相比,在低氮处理下锰、钙和镁质量分数均显著提高(P<0.05),分别提高61.67%、17.45%和10.08%,而高氮处理下锰和钙质量分数分别降低14.22%、30.32%。
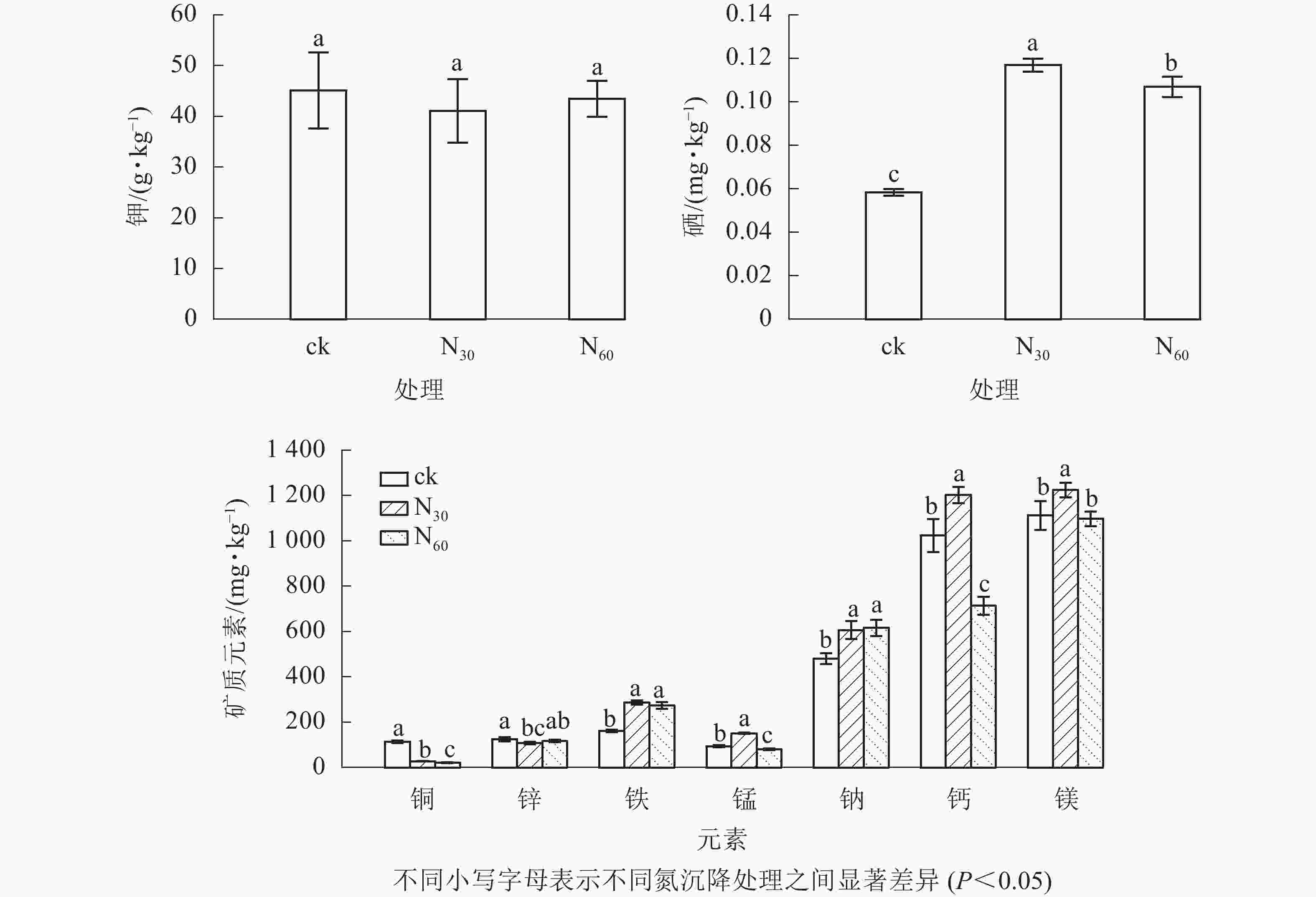
图 2 不同氮沉降水平下毛竹笋矿质元素的质量分数
Figure 2. Contents of mineral elements under different nitrogen (N) deposition
-
本研究毛竹笋中共检测出18种氨基酸,其中赖氨酸(Lys)、苯丙氨酸(Phe)、蛋氨酸(Met)、苏氨酸(Thr)、异亮氨酸(Ile)、亮氨酸(Leu)和缬氨酸(Val)为人体必需氨基酸。氮沉降显著提高毛竹笋中总氨基酸和必需氨基酸的质量分数,这与施肥[9, 26]和施氮肥[26]的研究效果相同。
环境因素会影响竹笋中总氨基酸质量分数,其中对氨基酸影响最大的因素为氮素供应水平[27]。氮素是植物体内氨同化和氨基酸合成的重要来源,是植物生理生化过程的重要营养物质[28]。土壤中的氮元素增加,为植物体合成氨基酸提供了丰富的氮源[9]。因此,氮沉降增加了毛竹笋中总氨基酸和必需氨基酸的质量分数,能够促进人体氨基酸平衡,减少因氨基酸失衡所造成的代谢紊乱、机体抵抗力下降等问题[29]。此外,氨基酸种类、含量和比例对竹笋中蛋白质种类和含量有一定影响,从而影响竹笋的品质[30]。其中,鲜味氨基酸(天冬氨酸和谷氨酸)和芳香类氨基酸(苯丙氨酸和酪氨酸)[31]质量分数在高氮处理下显著增加,说明过量氮沉降能提高毛竹笋的鲜味,使得竹笋的食用风味更好。苦味氨基酸(缬氨酸、异亮氨基酸、亮氨酸、酪氨酸、苯丙氨酸)[31]质量分数及比例在低氮下显著降低,而甜味氨基酸(甘氨酸、丙氨酸、脯氨酸、丝氨酸)[31]质量分数及比例在氮沉降下显著增加,表明适度的氮沉降有利于降低竹笋的苦涩味,增加甜味,从而提升其口感。
-
维生素C在增强体液免疫和细胞免疫方面具有显著功效,还能用来预防感冒、延缓衰老和防治冠心病等[29];粗纤维是影响竹笋食用口感的重要指标,在改善肠胃功能、降低血糖和胆固醇含量、防治肠癌和高脂血症等方面也具有重要作用[9];蛋白质在竹笋干物质组成中含量较高,同时也是生物体的主要组成部分和人体必需营养物质[32];可溶性总糖是评价竹笋品质的重要指标[33],也是人体能源物质的重要组成部分。
氮增加不仅有利于蔬菜、作物植株的生长发育,而且对提高果实产量、可溶性糖和蛋白质等营养成分、降低硝酸盐积累具有重要作用,从而改善植株的品质[24]。而土壤中氮素供应过高时,在硝化细菌的作用下会转化成硝酸根离子(
${\rm{NO}}_3^{-} $ )被蔬菜、作物等吸收。在植物体内积累过量硝酸盐,会对植物自身生长造成危害或品质下降,人们食用后也会对健康造成潜在的危害[34]。本研究发现:氮沉降促进了粗纤维和可溶性糖的积累,但降低了维生素C的质量分数。氮沉降下竹笋中粗纤维增加,与施肥促进粗纤维积累的研究结果相同[9]。膳食纤维作为“第七类营养素”,具有提高人体耐糖程度,降低胆固醇和刺激肠胃蠕动的作用[35-36]。氮沉降增加膳食纤维有助于改善竹笋品质。施肥促进竹笋中总糖的提高[37],其原因可能是竹子的光合作用因氮增加而增强,从而增加了运往笋体的糖分,使得笋体内的总糖提高[26]。氮沉降使竹笋中维生素C总体下降,但高氮下维生素C较低氮相比呈显著提高。其原因可能是,氮沉降使土壤中铵态氮(
${\rm{NH}}_4^{+} $ -N)、硝态氮(${\rm{NO}}_3^{-} $ -N)离子显著提高,同时也会导致土壤中离子失衡,使得硫酸根离子(${\rm{SO}}_4^{2-} $ )、氯离子(Cl−)、磷酸一氢根离子(${\rm{HPO}}_4^{2-} $ )积累,从而对竹笋维生素C的合成产生了一定的抑制作用,这与部分蔬菜的研究结果一致[38]。而离子浓度过高时,植物细胞离子通道会出现饱和现象,相关离子通道开放比率下降[39]。随着氮沉降水平的提高,土壤中${\rm{SO}}_4^{2-} $ 、Cl−、${\rm{HPO}}_4^{2-} $ 等离子浓度增加可能会使毛竹根系吸收这些离子的通道开放比率下降,从而降低其对竹笋中维生素C的抑制,故高氮下维生素C较低氮显著提高。 -
毛竹笋中这些元素含量之间存在明显差异,且对氮增加的响应不同,而人体中元素的缺乏和过量可导致多种疾病。氮沉降显著提高了硒、铁和钠质量分数,显著降低了铜质量分数;锰、钙和镁质量分数在低氮下增加,在高氮下降低。硒的缺乏会造成克山病和大骨节病,而氮沉降提高毛竹笋中硒质量分数,这使得人们通过食用同等质量毛竹笋摄入的硒更多,降低了患克山病和大骨节病的可能性;铁是造血元素,其含量的提高有助于食用者减少贫血、免疫力低、无力、头痛等问题[40];钠元素缺乏会使人出现恶心、食欲不振、心率加快、肌肉痉挛等反应[40],氮沉降对其含量的提高,有利于人体健康;成人每日可耐受铜的最高摄入量为8.0 mg·d−1[29]。本研究发现:氮沉降显著降低了毛竹笋中铜质量分数,因而可以降低铜过量所引起的黄疸肝炎、肝硬化、肠胃炎、癌症、Wilson病等疾病可能性[40]。氮沉降会影响土壤养分,同时影响土壤中钙离子、镁离子、钠离子和钾离子等盐基离子[41]。竹笋中不同矿质元素的变化可能与此相关,从而影响笋体内矿质元素含量,而元素间存在拮抗作用,所以不同矿质元素变化不同。
-
氮沉降显著增加了毛竹笋中总氨基酸、必需氨基酸、甜味氨基酸、粗纤维、硒、铁和钠的质量分数,显著降低了铜的质量分数,表明氮沉降提高毛竹笋的部分营养成分,可在一定程度上促进毛竹笋口感和营养品质的改善,有利于人体保健。
Effects of nitrogen application on nutritional components of Phyllostachys edulis shoot
-
摘要:
目的 探讨氮沉降对毛竹Phyllostachys edulis笋品质的影响,为竹笋品质改善和人体保健提供理论依据。 方法 通过人工喷氮模拟氮沉降方法,研究了3种不同水平的氮沉降[0 kg·hm−2·a−1(对照)、30 kg·hm−2·a−1(低氮,N30)和60 kg·hm−2·a−1(高氮,N60)]对毛竹笋不同氨基酸、营养成分(维生素C、粗纤维、粗蛋白质、可溶性糖)和矿质元素质量分数的影响。 结果 氮沉降对毛竹笋营养品质和矿质元素影响显著,氮沉降显著增加了总氨基酸、必需氨基酸和甜味氨基酸的质量分数(P<0.05),高氮沉降显著提高了鲜味、苦味和芳香类氨基酸的质量分数(P<0.05),而低氮沉降则相反;同时,氮沉降显著提高了毛竹笋中甜味氨基酸比例(P<0.05),显著降低了必需、鲜味、苦味和芳香类氨基酸比例(P<0.05);氮沉降显著降低了毛竹笋中维生素C质量分数(P<0.05),但却提高了粗纤维质量分数,对粗蛋白质和可溶性糖质量分数影响不显著;氮沉降显著提高了硒、铁和钠(P<0.05)质量分数,显著降低了铜(P<0.05)质量分数,低氮沉降显著增加了锰、钙和镁(P<0.05)质量分数,而高氮沉降则相反。 结论 适度的氮沉降对毛竹笋营养成分有一定的提升作用,有利于竹笋品质的改善和人体保健。图2表2参41 Abstract:Objective This paper aims to explore the effects of N deposition on Phyllostachys edulis shoots and provide theoretical basis for improving bamboo shoot quality and human health. Method A field experiment was conducted in a subtropical Ph. edulis plantation and the effects of three different levels of simulated N deposition (control: 0 kg·hm−2·a−1, N30: 30 kg·hm−2·a−1, and N60: 60 kg·hm−2·a−1) on the contents of amino acids, nutrients (vitamin C, crude fiber, crude protein, and soluble sugar) and mineral elements in Ph. edulis shoots were studied. Result N deposition significantly affected the nutrient quality and mineral elements of Ph. edulis shoots. Nitrogen deposition significantly increased the contents of total amino acids, essential amino acids, and sweet amino acids (P<0.05). High N deposition (N60) significantly increased the contents of umami amino acids, bitter amino acids, and aromatic amino acids (P<0.05), while low nitrogen deposition (N30) had the opposite effect. Nitrogen deposition significantly increased the proportion of sweet amino acids (P<0.05), but significantly decreased the proportion of essential amino acids, umami amino acids, bitter amino acids, and aromatic amino acids (P<0.05), and significantly reduced vitamin C content(P<0.05) but increased crude fiber content, and had no significant effect on the content of crude protein and soluble sugar. Nitrogen deposition significantly increased the contents of Se, Fe and Na but significantly decreased the content of Cu (P<0.05). Low N deposition (N30) significantly increased Mn, Ca and Mg contents, while the N60 treatment was the opposite(P<0.05). Conclusion Moderate N deposition has a certain promotion effect on nutritional components of Ph. edulis shoots, which is conducive to the improvement of bamboo shoot quality and human health. [Ch, 2 fig. 2 tab. 41 ref.] -
Key words:
- botany /
- nitrogen deposition /
- Phyllostachys edulis shoot /
- nutritional components /
- mineral elements /
- health food
-
图 1 不同氮沉降水平下毛竹笋维生素C、粗纤维、粗蛋白质和可溶性糖质量分数
Figure 1 Contents of vitamin C, crude fiber, crude protein and soluble sugar under different nitrogen (N) deposition
图 2 不同氮沉降水平下毛竹笋矿质元素的质量分数
Figure 2 Contents of mineral elements under different nitrogen (N) deposition
表 1 毛竹林分和表层土壤(0~20 cm)特征
Table 1. Original characteristics of the Ph. edulis plantation and surface soil(0−20 cm)
处理 密度/
(株·hm−2)胸径/
cm土壤密度/
(g·cm−3)土壤有机碳/
(mg·g−1)土壤全氮/
(mg·g−1)土壤全磷/
(mg·g−1)土壤pH 对照(ck) 3 362±309 10.16±0.13 0.97±0.07 23.73±0.24 1.11±0.04 0.52±0.01 4.46±0.01 低氮(N30) 3 408±382 10.31±0.32 0.98±0.04 22.56±1.17 1.17±0.09 0.57±0.05 4.43±0.04 高氮(N60) 3 208±187 10.76±0.59 1.00±0.05 23.15±1.74 1.21±0.12 0.54±0.03 4.48±0.08 说明:数值为均值±标准差  下载: 导出CSV
下载: 导出CSV
表 2 不同氮沉降水平下毛竹笋不同类型氨基酸质量分数及比例
Table 2. Contents and proportions of different types of amino acids under different nitrogen(N) deposition
氮处理 必需氨基酸 鲜味氨基酸 甜味氨基酸 苦味氨基酸 芳香类氨基酸 总氨基酸 质量分数/
(mg·g−1)比例/% 质量分数/
(mg·g−1)比例/% 质量分数/
(mg·g−1)比例/% 质量分数/
(mg·g−1)比例/% 质量分数/
(mg·g−1)比例/% 质量分数/
(mg·g−1)对照(ck) 4.04±0.16 Bb 22.57 0.93±0.03 Eb 5.20 2.00±0.10 Db 11.17 2.78±0.11 Cb 15.53 1.32±0.02 Eb 7.37 17.90±0.61 Ab 低氮(N30) 5.40±0.21 Ba 20.80 0.34±0.02 Ec 1.31 3.93±0.18 Ca 15.14 2.47±0.14 Dc 9.51 0.96±0.07 Ec 3.70 25.96±1.10 Aa 高氮(N60) 5.48±0.25 Ba 19.87 1.12±0.04 Da 4.06 4.13±0.14 Ca 14.97 3.22±0.35 Ca 11.68 1.60±0.08 Da 5.80 27.58±1.27 Aa 说明:同列不同小写字母表示不同氮沉降水平对各氨基酸影响差异显著(P<0.05),同行不同大写字母表示相同氮沉降水平对各氨基 酸影响差异显著(P<0.05)
下载: 导出CSV
-
[1] 洪伟, 林存炎, 吴承祯, 等. 毛竹笋品质的区域分异性分析[J]. 福建林学院学报, 2007, 27(4): 289 − 293. HONG Wei, LIN Cunyan, WU Chengzhen, et al. Analysis of the regions variation for nutrition quality of the Phyllostachys heterocycla var. pubescens bamboo shoots [J]. J Fujian Coll For, 2007, 27(4): 289 − 293. [2] 刘耀荣. 毛竹笋期的营养动态[J]. 林业科学研究, 1990, 3(4): 363 − 367. LIU Yaorong. Nutrient dynamics in the shoot stage of mao bamboo [J]. For Res, 1990, 3(4): 363 − 367. [3] NIRMALA C, BISHT M S, LAISHRAM M. Bioactive compounds in bamboo shoots: health benefits and prospects for developing functional foods [J]. Int J Food Sci Technol, 2014, 49(6): 1425 − 1431. [4] CHOUDHURY D, SAHU J K, SHARMA G D. Value addition to bamboo shoots: a review [J]. J Food Sci Technol, 2012, 49(4): 407 − 414. [5] 徐有明, 郝培应, 费本华. 竹笋形态发育构建过程中解剖结构及化学成分的动态变化[J]. 东北林业大学学报, 2008, 36(4): 8 − 11, 16. XU Youming, HAO Peiying, FEI Benhua. Dynamic change of anatomical structureand chemical constitutes of bamboo shoots for Phyllostachys pubescence at developmental stage [J]. J Northeast For Univ, 2008, 36(4): 8 − 11, 16. [6] 周文伟, 何奇江, 叶春球, 等. 不同季节毛竹笋营养成分比较分析[J]. 浙江林业科技, 2013, 33(4): 64 − 67. ZHOU Wenwei, HE Qijiang, YE Chunqiu, et al. Comparative analysis of nutrients in bamboo shoot at different seasons [J]. J Zhejiang For Sci Technol, 2013, 33(4): 64 − 67. [7] 王丹丹, 李琴, 贺亮, 等. 不同贮藏温度条件下鲜切毛竹笋品质的变化[J]. 食品与发酵科技, 2016, 52(6): 72 − 76, 90. WANG Dandan, LI Qin, HE Liang, et al. Effect of different storage temperatures on the quality of fresh-cut Phyllostachys heterocycla [J]. Food Ferment Sci Technol, 2016, 52(6): 72 − 76, 90. [8] 王海平, 张敏, 朱伦辉, 等. 不同包装材料对MAP毛竹笋品质影响的研究[J]. 四川食品与发酵, 2006, 42(6): 50 − 52. WANG Haiping, ZHANG Min, ZHU Lunhui, et al. Research on the effect of the quality of bamboo shoots with different packaging materials under MAP [J]. Sichuan Food Ferment, 2006, 42(6): 50 − 52. [9] 邱永华, 金爱武, 张四海, 等. 不同施肥方式对竹笋品质的影响[J]. 竹子学报, 2017, 36(1): 41 − 48. QIU Yonghua, JIN Aiwu, ZHANG Sihai, et al. Effect of different fertilization methods on quality of bamboo shoots [J]. J Bamboo Res, 2017, 36(1): 41 − 48. [10] 胡冬南, 陈立新, 李发凯, 等. 配方施肥对毛竹笋材的影响[J]. 江西农业大学学报, 2004, 26(2): 196 − 199. HU Dongnan, CHEN Lixin, LI Fakai, et al. Effect of optimun amount and proportion of fertilizer on bamboo shoots and timbers [J]. Acta Agric Univ Jiangxi, 2004, 26(2): 196 − 199. [11] 郭晓敏, 陈广生, 牛德奎, 等. 平衡施肥对毛竹笋产量的影响效应研究[J]. 江西农业大学学报(自然科学版), 2003, 25(1): 48 − 53. GUO Xiaomin, CHEN Guangsheng, NIU Dekui, et al. A study on the effects of balance fertilization on bamboo shoot yield [J]. Acta Agric Univ Jiangxi, 2003, 25(1): 48 − 53. [12] 程平, 彭九生, 王海霞. 毛竹增产剂及幼竹竹腔施肥对竹笋品质的影响研究[J]. 竹子学报, 2007, 26(3): 28 − 31. CHENG Ping, PENG Jiusheng, WANG Haixia. Effects of cavity-fertilizing on shoot qualities of moso bamboo [J]. J Bamboo Res, 2007, 26(3): 28 − 31. [13] 常运华, 刘学军, 李凯辉, 等. 大气氮沉降研究进展[J]. 干旱区研究, 2012, 29(6): 972 − 979. CHANG Yunhua, LIU Xuejun, LI Kaihui, et al. Research progress in atmospheric nitrogen deposition [J]. Arid Zone Res, 2012, 29(6): 972 − 979. [14] 莫江明, 薛璟花, 方运霆. 鼎湖山主要森林植物凋落物分解及其对N沉降的响应[J]. 生态学报, 2004, 24(7): 1413 − 1420. MO Jiangming, XUE Jinghua, FANG Yunting. Litter decomposition and its responses to simulated N deposition for the majorplants of Dinghushan forests in subtropical China [J]. Acta Ecol Sin, 2004, 24(7): 1413 − 1420. [15] JIA Yanlong, YU Guirui, HE Nianpeng, et al. Spatial and decadal variations in inorganic nitrogen wet deposition in China induced by human activity [J]. Sci Rep, 2014, 4: 3763. doi: 10.1038/srep03763. [16] ACKERMAN D, MILLET D B, CHEN Xin. Global estimates of inorganic nitrogen deposition across four decades [J]. Global Biogeochem Cycles, 2019, 33: 100 − 107. [17] SONG Xinzhang, CHEN Xiaofeng, ZHOU Guomo, et al. Observed high and persistent carbon uptake by Moso bamboo forests and its response to environmental drivers [J]. Agric For Meteorol, 2017, 247: 467 − 475. [18] 彭春菊, 李全, 顾鸿昊, 等. 模拟氮沉降及经营方式对毛竹林土壤酶活性的影响[J]. 应用生态学报, 2017, 28(2): 423 − 429. PENG Chunju, LI Quan, GU Honghao, et al. Effects of simulated nitrogen deposition and management type on soil enzyme activities in moso bamboo forest [J]. Chin J Appl Ecol, 2017, 28(2): 423 − 429. [19] 吕建华. 氮沉降对毛竹叶化学计量、养分重吸收及凋落叶养分归还的影响[D]. 杭州: 浙江农林大学, 2018. LÜ Jianhua. Effects of Nitrogen Deposition on Stoichiometry, Nutrient Resorption of Leaf and Nutrient Return of Leaf Litter in a Moso Bamboo Forest[D]. Hangzhou: Zhejiang A&F University, 2018. [20] 杜莹, 鲍永新, 裘子炎, 等. 模拟氮沉降对毛竹非结构性碳的影响[J]. 林业科学, 2017, 53(7): 10 − 17. DU Ying, BAO Yongxin, QIU Ziyan, et al. Effects of simulated nitrogen deposition on non-structural carbohydrates of moso bamboo [J]. Sci Silv Sin, 2017, 53(7): 10 − 17. [21] SONG Xinzhang, PENG Changhui, CIAIS P, et al. Nitrogen addition increased CO2 uptake more than non-CO2 greenhouse gases emissions in a moso bamboo forest [J]. Sci Adv, 2020, 6(12): eaaw5790. doi: 10.1126/sciadv.aaw5790. [22] 中华人民共和国国家卫生和计划生育委员会. 食品安全国家标准食品中抗坏血酸的测定: GB 5009.86−2016[S]. 北京: 中国标准出版社, 2016. [23] 宁正祥. 食品成分分析手册[M]. 北京: 中国轻工业出版社, 1998. [24] 李云云, 李强, 余宏军, 等. 花期短期低氮处理对黄瓜抗坏血酸及可溶性糖含量的影响[J]. 中国土壤和肥料, 2019, 56(5): 46 − 52. LI Yunyun, LI Qiang, YU Hongjun, et al. Effects of short-term low-nitrogen treatment on AsA and soluble sugar contents of cucumber during flowering stage [J]. Soil Fert Sci China, 2019, 56(5): 46 − 52. [25] 方炳华, 肖珊美. 湿法消化-火焰原子吸收光谱法测定植株中矿质元素[J]. 中国林副特产, 2005(5): 5 − 6. FANG Binghua, XIAO Shanmei. Measuring of the mineral elements in plants using the flame atom-imbibing spectrum method [J]. For By-Prod Spec China, 2005(5): 5 − 6. [26] 朱元洪, 孙羲, 洪顺山. 施肥和土壤养分对毛竹笋营养成分的影响[J]. 土壤学报, 1991, 28(1): 40 − 49. ZHU Yuanhong, SUN Xi, HONG Shunshan. The effect of fertilization and soil fertility on the nutritive composition of bamboo (Phyllostachys pubescense) shoot [J]. Acta Pedol Sin, 1991, 28(1): 40 − 49. [27] 刘振勇, 潘雁红, 钟哲科, 等. 不同肥料对覆盖早竹林竹笋产量和品质的影响[J]. 竹子研究汇刊, 2015, 34(4): 17 − 22. LIU Zhenyong, PAN Yanhong, ZHONG Zheke, et al. The effects of fertilizers on yield and quality of bamboo shoots of Phyllostachys praecox stands with mulch management [J]. J Bamboo Res, 2015, 34(4): 17 − 22. [28] 徐春英. 冬小麦氨基酸代谢与抗旱性关系的研究[D]. 北京: 中国农业科学院, 2008. XU Chunying. Studies on the Relationships Between Amino Acids Metabolism and Drought-Resistance of Winter Wheat[D]. Beijing: Chinese Academy of Agricultural Sciences, 2008. [29] 中国营养学会. 中国居民膳食营养素参考摄量标准[M]. 北京: 中国轻工业出版社, 2000. [30] 陈闻, 吴家森, 姜培坤, 等. 不同施肥对雷竹林土壤肥力及肥料利用率的影响[J]. 土壤学报, 2011, 48(5): 1022 − 1028. CHEN Wen, WU Jiasen, JIANG Peikun, et al. Effects of different fertilization on soil fertility quality, fertilizeruse efficiency, and bamboo shoot yields of Phyllostachys praecox stand [J]. Acta Pedol Sin, 2011, 48(5): 1022 − 1028. [31] 郭子武, 杨丽婷, 林华, 等. 沙县苦竹笋外观、营养和食味品质变异的海拔效应[J]. 生态学杂志, 2019, 38(1): 83 − 88. GUO Ziwu, YANG Liting, LIN Hua, et al. Effects of altitude on the variation of appearance, nutrition, and taste for bamboo shootsof Pleioblastus amarus in Shaxian, Fujian Province [J]. Chin J Ecol, 2019, 38(1): 83 − 88. [32] 王裕霞, 张光楚, 李兴伟. 优良丛生笋用竹及杂种竹竹笋品质评价的研究[J]. 竹子研究汇刊, 2005, 24(4): 39 − 44. WANG Yuxia, ZHANG Guangchu, LI Xingwei. An evaluation on shoot quality of sympodial bamboo species and their hybrids [J]. J Bamboo Res, 2005, 24(4): 39 − 44. [33] 申德省. 鲜切雷竹笋木质化和褐变的控制研究[D]. 杭州: 浙江工商大学, 2015. SHEN Dexing. Study on the Lignification and Browning of Fresh-Cut Bamboo Shoot[D]. Hangzhou: Zhejiang Gongshang University, 2015. [34] 周泽义, 胡长敏, 王敏健, 等. 中国蔬菜硝酸盐和亚硝酸盐污染因素及控制研究[J]. 环境科学进展, 1999, 7(5): 1 − 13. ZHOU Zeyi, HU Changmin, WANG Minjian, et al. The nitrate and nitrite contamination in vegetables and its control in China [J]. Chin J Environ Eng, 1999, 7(5): 1 − 13. [35] 谢碧霞, 李安平. 膳食纤维[M]. 北京: 科学出版社, 2006. [36] 吴良如, 高贵宾, 白瑞华, 等. 竹笋膳食纤维的研究与应用[J]. 竹子研究汇刊, 2010, 29(2): 1 − 5, 33. WU Liangru, GAO Guibin, BAI Ruihua, et al. Research and application of dietary fiber from bamboo shoots [J]. J Bamboo Res, 2010, 29(2): 1 − 5, 33. [37] 郭晓敏, 牛德奎, 张斌, 等. 集约经营毛竹林平衡施肥效应研究[J]. 西南林学院学报, 2005, 25(4): 84 − 88. GUO Xiaomin, NIU Dekui, ZHANG Bin, et al. Research on the effect of balanced fertilization in intensively bred Phyllostachys heterocycls cv. pubescens plantations [J]. J Southwest For Coll, 2005, 25(4): 84 − 88. [38] 丁文雅. 氮肥对蔬菜品质影响的研究进展[J]. 北京农业, 2015, 35(12): 117 − 118. DING Wenya. Research progress on effect of nitrogen fertilizer on vegetable quality [J]. Beijing Agric, 2015, 35(12): 117 − 118. [39] 何龙飞, 刘友良, 沈振国, 等. 植物离子通道特征、功能、调节与分子生物学[J]. 植物学通报, 1999, 16(5): 517 − 525. HE Longfei, LIU Youliang, SHEN Zhenguo, et al. Ion channel characteristics, functions, regulatory factors and molecular biology in plants [J]. Chin Bull Bot, 1999, 16(5): 517 − 525. [40] 江元汝. 化学与健康[M]. 北京: 科学出版社, 2009. [41] 刘佳婷, 廖迎春, 谢越, 等. 氮沉降对土壤养分影响研究综述[J]. 中国水土保持, 2019, 40(9): 63 − 65. LIU Jiating, LIAO Yingchun, XIE Yue, et al. Review on the effects of nitrogen deposition on soil nutrients [J]. Soil Water Conserv China, 2019, 40(9): 63 − 65. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20200256
 点击查看大图
点击查看大图
计量
- 文章访问数: 2534
- HTML全文浏览量: 741
- PDF下载量: 36
- 被引次数: 0







