




-
枳Poncirus trifoliata是当前柑橘Citrus种植中应用最广泛的砧木,且已有研究表明:枳在抗旱能力方面明显优于其他柑橘,具有广泛的适应性和对环境压力的高度耐受性[1−2]。高渗透压诱导的钙离子(Ca2+)增加通道(hyperosmolality-induced [Ca2+] increase channel,OSCA)是一类具有Ca2+渗透特性的阳离子通道蛋白,广泛存在于植物中的跨膜蛋白家族,其主要功能是调节细胞内外的Ca2+平衡,从而帮助植物响应环境胁迫,尤其在干旱胁迫下起到重要作用。目前OSCA基因家族在枳耐旱响应中的作用尚不清楚。YUAN等[3]通过基于钙成像的遗传筛选策略,在高渗透应激下分离出拟南芥Arabidopsis thaliana突变体OSCA1.1,并发现OSCA1是一个未知的高渗透性门控钙通道,可能在渗透应激中发挥作用。OSCA1.1属于拟南芥OSCA基因家族的一员,且在其他植物和真核生物中有同源基因[3−6]。该家族的蛋白质含有钙依赖性通道结构域(DUF221),在真核生物中保守[7]。在干旱条件下,OSCA蛋白作为渗透传感器激活,调节细胞内Ca2+浓度,启动抗旱反应,包括调控气孔开闭和脱落酸(ABA)信号途径[6−8]。已有研究表明:多个物种的OSCA基因在干旱胁迫下表达上调,如玉米Zea mays ZmOSCA2.4提高拟南芥的耐旱性[6],番茄Solanum lycopersicum SlOSCA6和SlOSCA8在聚乙二醇(PEG)处理下表达显著上调[9],水稻Oryza sativa OsOSCA3.1在干旱模拟条件下也表现出表达上调[4]。
本研究分析了枳OSCA基因家族的生物信息学特征,包含其序列特征、理化性质以及启动子区域的顺式作用元件,同时利用转录组和实时荧光定量PCR (RT-qPCR)分析OSCA基因家族在不同组织、不同干旱胁迫程度下的表达特征,为后期解析OSCA调控柑橘砧木枳的耐旱机制和利用该基因开展柑橘砧木抗性育种提供科学依据。
-
以枳老籽播种生长3个月的实生苗为试材[10],育苗期为2024年2月中旬至5月中旬,培养于湖南农业大学国家柑橘改良中心长沙分中心温室大棚同一栽培槽内。选取长势一致的3月龄枳苗置于温度为25 ℃,相对湿度为75%,光照强度为4 000 lx,光周期为16 h光照/8 h黑暗条件的人工气候室内,用去离子水反复漂洗根系后,移栽固定于2 L水培盘漂,采用Hoagland营养液预培养1 d。在含质量分数为20%PEG 6000的Hoagland营养液中模拟干旱胁迫[5, 9],分别于处理0 min (T1, ck)、5 min (T2)、20 min (T3)、1 h (T4)、6 h (T5)和24 h (T6)后,采集侧根及植株形态学上端第2、3枚完全展开叶片,并用去离子水反复冲洗根部,每个采样点进行3次生物学重复,每次重复5株样本,样品经液氮速冻后,保存在−80 ℃冰箱中。
-
从拟南芥基因组数据库TAIR-Home Page 获取拟南芥OSCA基因家族的氨基酸序列,作为参考序列。从华中农业大学甜橙基因组数据库(Citrus Pan-genome to Breeding Database, CPBD,
)中获取甜橙Citrus sinensis、枳全基因组氨基酸序列。使用TB tools软件Two Sequence Files程序对枳、甜橙、拟南芥蛋白序列进行比对,设定比对期望值(E)<10−5,以确保比对结果的统计学显著性,并设定同源度>60%,以筛选进化上较为保守的同源基因[11−12],最终获得甜橙和枳的OSCA家族候选基因。运用LINUX虚拟机系统,采用HMM Search功能获得的结果,并结合Blast分析,对甜橙和枳OSCA基因家族的候选基因进行比对和筛选,随后,利用每千个碱基的转录每百万映射读取的片段 (PFAM) ( http://pfam.xfam.org/ )对这些候选基因进行预测分析,最终筛选出含完整DUF221结构域的基因,并确定它为甜橙和枳OSCA的基因家族成员。为进一步分析枳OSCA蛋白的功能,利用WoLF PSORT (https://wolfpsort.Hgc.jp/ )进行枳OSCA蛋白的亚细胞定位预测。同时,利用ExPASy (https://web.expasy.org/protparam/ )对枳OSCA基因家族成员的蛋白序列进行分析以及氨基酸数量、分子量、等电点等理化性质进行分析。 -
从CPBD获取枳OSCA基因核苷酸序列及其编码序列。使用MEME工具(
https://meme-suite.org/meme/ )对枳OSCA基因编码的蛋白质进行保守基序分析,设置基序(Motif)长度范围为6~50个氨基酸,查找Motif数量设定为20,以研究其基序顺序和排列顺序。使用TBtools软件中的Gene Structure View(Advanced)功能对分析结果进行基因结构的可视化。 -
为深入分析枳OSCA基因家族的进化关系,选择拟南芥和甜橙OSCA基因家族作为对照,构建系统发育树。从TAIR-Home Page获取拟南芥OSCA家族蛋白序列,从CPBD获取枳和甜橙OSCA蛋白序列,将枳、甜橙OSCA蛋白序列导入MEGA 5.0,采用Clustal W进行多重序列比对,采用最大似然法(Maximum likelihood)构建系统进化树(Bootstrap值设定为1 000)[13],最后,利用ITOL在线网站(
https://itol.embl.de/ )对所得进化树进行编辑与可视化处理。为进一步分析PtOSCA基因启动子区的顺式作用元件,首先使用TBtools提取转录起始位点上游2 000 bp的序列,在PlantCARE (
http://bioinformatics.psb.ugent.be/webtools/plantcare/html/ )网站对枳OSCA基因的启动子区域进行顺式作用元件的预测分析[14]。 -
从CPBD下载枳的13个OSCA基因成员在果肉、叶、胚珠、根等组织中的FPKM值,并基于log2 FPKM值生成基因表达热图[15]。此外,基于前期转录组数据获得枳OSCA基因在轻度干旱、中度干旱、重度干旱胁迫下枳根组织的表达数据,以FPKM值为基础,结合|log2FC|≥1.5且P<0.05为筛选条件[5, 16],其中,FC为差异倍数,筛选出在不同干旱胁迫下显著差异表达的PtOSCAs基因。基因表达热图的绘制通过TB tools软件中的HeatMap功能完成,以展示PtOSCAs基因在不同组织及不同干旱胁迫条件下的表达特征。基于在不同组织中的表达特异性可以为预测潜在调控的功能提供线索[17]。
-
采用植物RNA提取试剂盒(SteadyPure,艾科瑞生物)提取枳侧根和叶总RNA。利用反转录试剂盒(Evo M-MLV,艾科瑞)进行逆转录反应。根据PtOSCA基因的CDS序列设计相应引物,委托擎科生物科技有限公司合成(表1)。RT-qPCR在CFX96 real-time PCR仪(Bio-Rad)中进行,采用的试剂盒为SsoAdvancedTM SYBR® Green Supermix。PCR程序为:95 ℃ 3 min,95 ℃ 3 s,60 ℃退火30 s,40个循环;72 ℃延伸5 min,4 ℃保存;熔解曲线65~95 ℃,每0.05 s温度增量0.5 ℃。
表 1 实时荧光定量PCR引物
Table 1. Primers for quantitative real-time PCR analysis
基因名称 正向引物(5′→3′) 反向引物(5′→3′) PtOSCA1 TGGAACGAGCAAGGGAACCA TATGTTTGCTTGGCAACGGAG PtOSCA2 GCACACCGGTGCATTTGTAA ACAGCAGAATCCAATCCCCC PtOSCA3 CAGAGGGAACAAGACCGACC GCAGAAGCTGCAGTTACCCT PtOSCA4 AGACTGAAAAGGACAGGGCG GGGTGTCACTGTGGCATACA PtOSCA5 CCAAACCCACCGAGTCAAGT AGCAGAGCATGTGAAGGTCC PtOSCA6 GGAGCTGCAATTGCTTTCCATT TAAGGCCTTGCACAACCAGC PtOSCA7 CTTCCGTGGCCACATTTCCA ACTTTCTCGGACACTGCGTT PtOSCA8 TGGATCAACAAGATGAGCTGGA CCAATCTCTGCCGTCTCTGC PtOSCA9 TTTGGAACGGAGGACGTCAG AATGCCAGCCCATCCATCAA PtOSCA10 TTCTTCAGTTGTTCCTATCATTCG CCACCAGACCTTTGAACAAGC PtOSCA11 CCGCTCAGTTCCTCCTCATC ACTCCATGCATTCCTTTTTCAAT PtOSCA12 ACAGTCGCATTCGGTTTCAC CCCTGATGATCCTTATGTCCAGA PtOSCA13 AAGGCTTGACTCGACTTCACC TTGCCCCATAATGACCAGCA PtActin CCGACCGTATGAGCAAGGAAA TTCCTGTGGACAATGGATGGA -
基因相对表达量的计算采用2-ΔΔCt方法,基因相对表达水平通过Excel 2021进行分析,数据的显著性差异采用SPSS 29.0统计处理,绘图使用Prism 10.0。
-
利用枳的全基因组序列和OSCA基因家族的保守结构DUF221,通过生物信息学分析鉴定得到枳OSCA基因家族的13个基因,并根据在染色体上的定位顺序,将这些基因依次命名为PtOSCA1~PtOSCA13 (表2)。其中,PtOSCA12和PtOSCA13未被注释到具体染色体,其余基因不均等地分布在枳18条染色体中的7条染色体上:2号染色体上4个,4号染色体和5号染色体上各2个,3号、6号和7号染色体上各1个。通过对这些基因的长度、氨基酸序列长度、等电点和分子量分析发现:已鉴定的PtOSCA基因长度为2 103~4 497 bp,PtOSCA蛋白的分子量为46 921.00~93 051.95 Da,平均值为81.34 kDa。PtOSCA的氨基酸数量为409~807个,平均值为716个。等电点为6.75~9.52,平均值为8.77。根据亚细胞定位预测结果,PtOSCA4和PtOSCA5定位于细胞膜、细胞壁和叶绿体,PtOSCA2和PtOSCA9定位于细胞膜和叶绿体,其余9个成员则定位于细胞膜。
表 2 枳OSCA基因家族信息
Table 2. Information on OSCA gene family in P. trifoliata
基因名称 基因 ID 基因全长/bp 氨基酸数量/个 等电点 分子量/Da 染色体 亚细胞定位 PtOSCA1 Pt2g013910.1 2 280 759 9.20 86 802.76 2 细胞膜 PtOSCA2 Pt2g013930.1 2 280 759 8.96 86 861.76 2 细胞膜,叶绿体 PtOSCA3 Pt2g023140.1 2 181 726 9.17 81 688.79 2 细胞膜 PtOSCA4 Pt2g028480.4 2 319 772 9.30 88 145.30 2 细胞膜,细胞壁,叶绿体 PtOSCA5 Pt3g032590.1 3 006 758 9.08 86 482.77 3 细胞膜,细胞壁,叶绿体 PtOSCA6 Pt4g000270.1 2 127 708 8.74 80 821.35 4 细胞膜 PtOSCA7 Pt4g001040.1 2 103 700 8.85 81 011.45 4 细胞膜 PtOSCA8 Pt5g011440.2 2 229 742 8.50 83 798.96 5 细胞膜 PtOSCA9 Pt5g018480.1 2 424 807 9.52 93 051.95 5 细胞膜,叶绿体 PtOSCA10 Pt6g003200.3 4 494 682 8.86 77 495.58 6 细胞膜 PtOSCA11 Pt7g021120.1 2 424 786 6.75 89 331.88 7 细胞膜 PtOSCA12 PtUn012370.2 2 139 712 8.38 80 420.82 未知 细胞膜 PtOSCA13 PtUn037020.1 3 889 409 9.23 46 921.00 未知 细胞膜 -
为探讨枳OSCA基因家族的功能特性及其系统进化关系,基于15个AtOSCAs、12个CsOSCAs和13个PtOSCAs基因编码的蛋白序列,构建了系统进化树(图1)。OSCA蛋白可分为四大类: Ⅰ类包含17个OSCAs成员,其中包含5个PtOSCAs成员(PtOSCA1、PtOSCA2、PtOSCA4、PtOSCA5、PtOSCA9)、8个AtOSCAs成员(AtOSCA1.1~AtOSCA1.8)和4个CsOSCAs成员(CsOSCA2、CsOSCA4、CsOSCA6、CsOSCA9);Ⅱ类包含17个OSCAs成员,其中6个PtOSCAs成员(包括PtOSCA6、PtOSCA7、PtOSCA10、PtOSCA11、PtOSCA13)、5个AtOSCAs成员(主要包括AtOSCA2.1~AtOSCA 2.5)和6个CsOSCAs成员(包括CsOSCA5、CsOSCA7、CsOSCA8、CsOSCA10~CsOSCA12);Ⅲ类包含3个OSCAs成员,即PtOSCA3、AtOSCA3.1和CsOSCA3;Ⅳ类包含3个OSCAs成员,即PtOSCA11、AtOSCA4.1和CsOSCA1。系统发育分析结果表明:枳OSCAs基因与拟南芥和甜橙OSCAs基因家族一同分为4个分支,说明枳OSCAs基因家族经历了与拟南芥和甜橙OSCAs基因家族相似的进化历史。此外,CsOSCAs基因与PtOSCAs基因之间的进化枝长度较短,表明来自枳与甜橙的OSCA蛋白具有更高的相似性,枳和甜橙之间的系统发育关系更加密切,这与它们同属于柑橘亚科Aurantioideae有关。
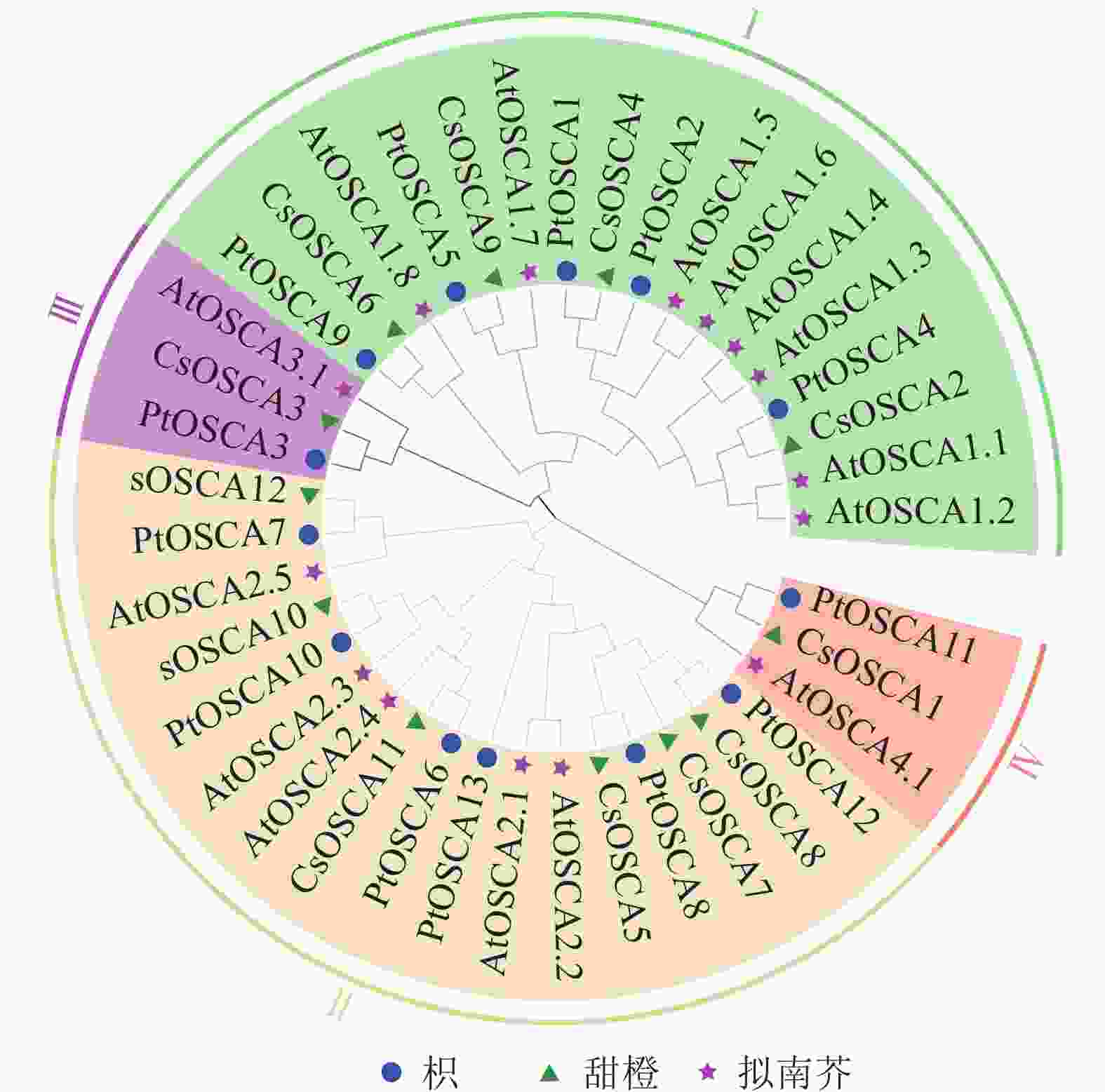
图 1 枳、甜橙、拟南芥的OSCA基因编码产物系统进化树
Figure 1. Phylogenetic tree of the coding products of the OSCA gene in P. trifoliata,C. sinensis and A.thaliana
-
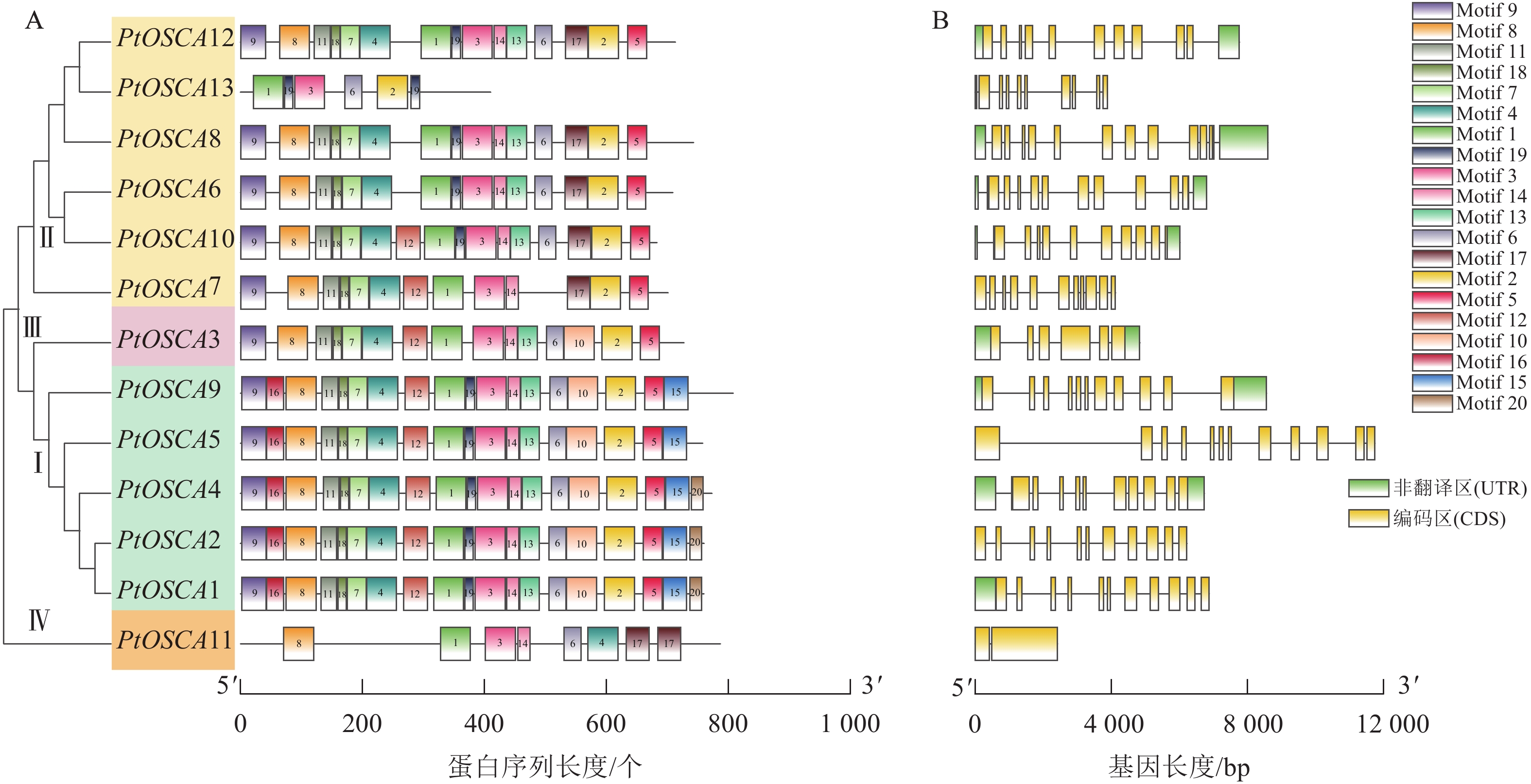
保守结构域、外显子-内含子结构及功能域比较分析表明:13个PtOSCAs基因共分为4组(图2),该基因家族的蛋白序列具有高度保守性,所有成员均包含保守的Motif 1和Motif 3基序,且这些基序排列顺序相同。Ⅰ类中的Motif基序最多,其中PtOSCA1、PtOSCA2、PtOSCA4共享相同的Motif基序,涵盖了Motif 1~Motif 20。相比之下,Ⅱ类中的PtOSCA13和Ⅳ类中的PtOSCA11的Motif数量最少。基于PtOSCAs基因的编码序列(coding sequence, CDS)和相应的基因组DNA序列的可视化分析结果显示:该基因家族大多数成员都富含内含子,且其内含子分布模式相对保守,多数基因具有相似数量的内含子。其中Ⅰ、Ⅱ、Ⅲ和Ⅳ类基因的内含子数量分别为10~11、9~12、5和1个。
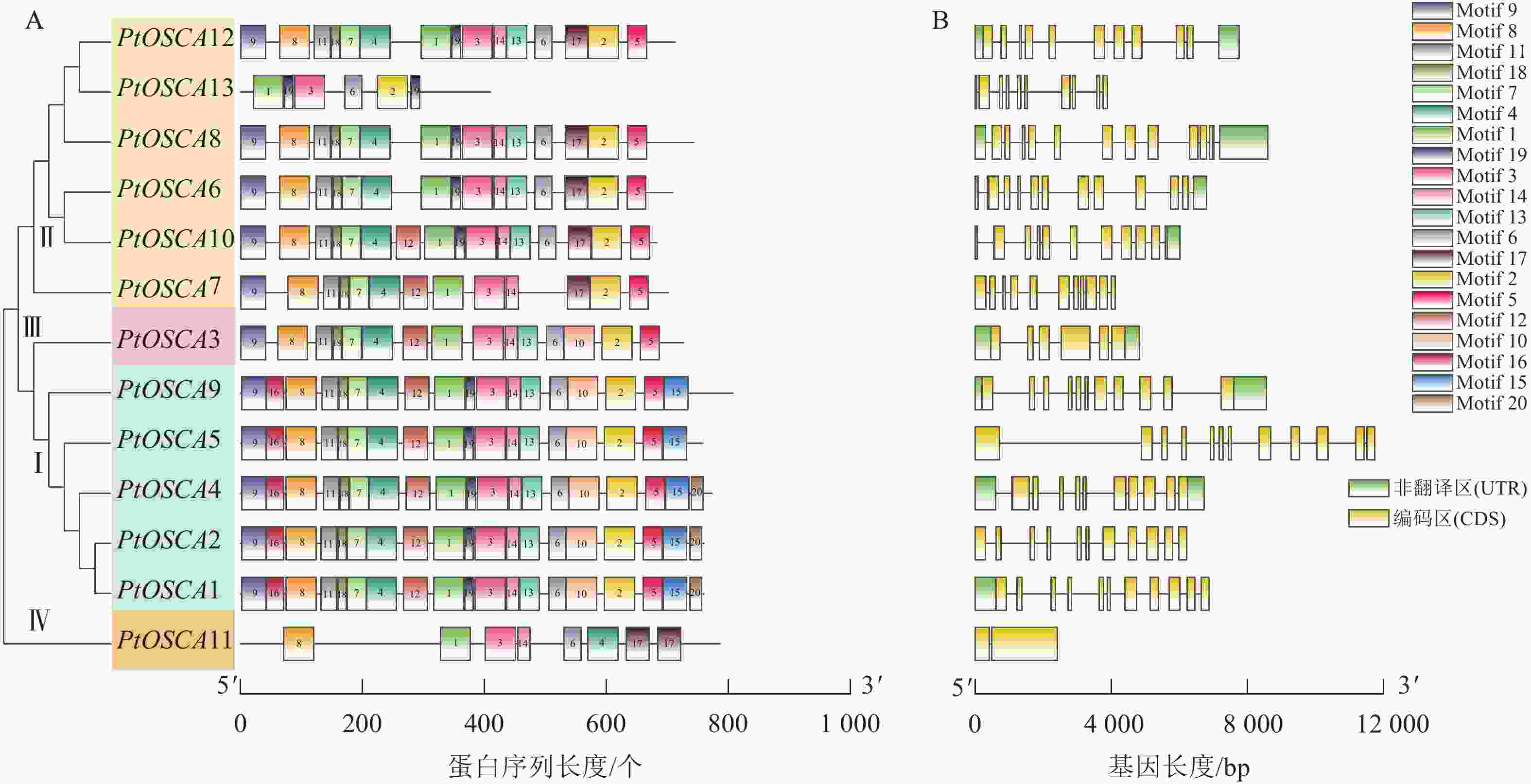
图 2 枳OSCA家族成员蛋白基序(A)和基因结构(B)分析
Figure 2. Analysis of conserved domains (A) and gene structure (B) of OSCA family members in P. trifoliata
-
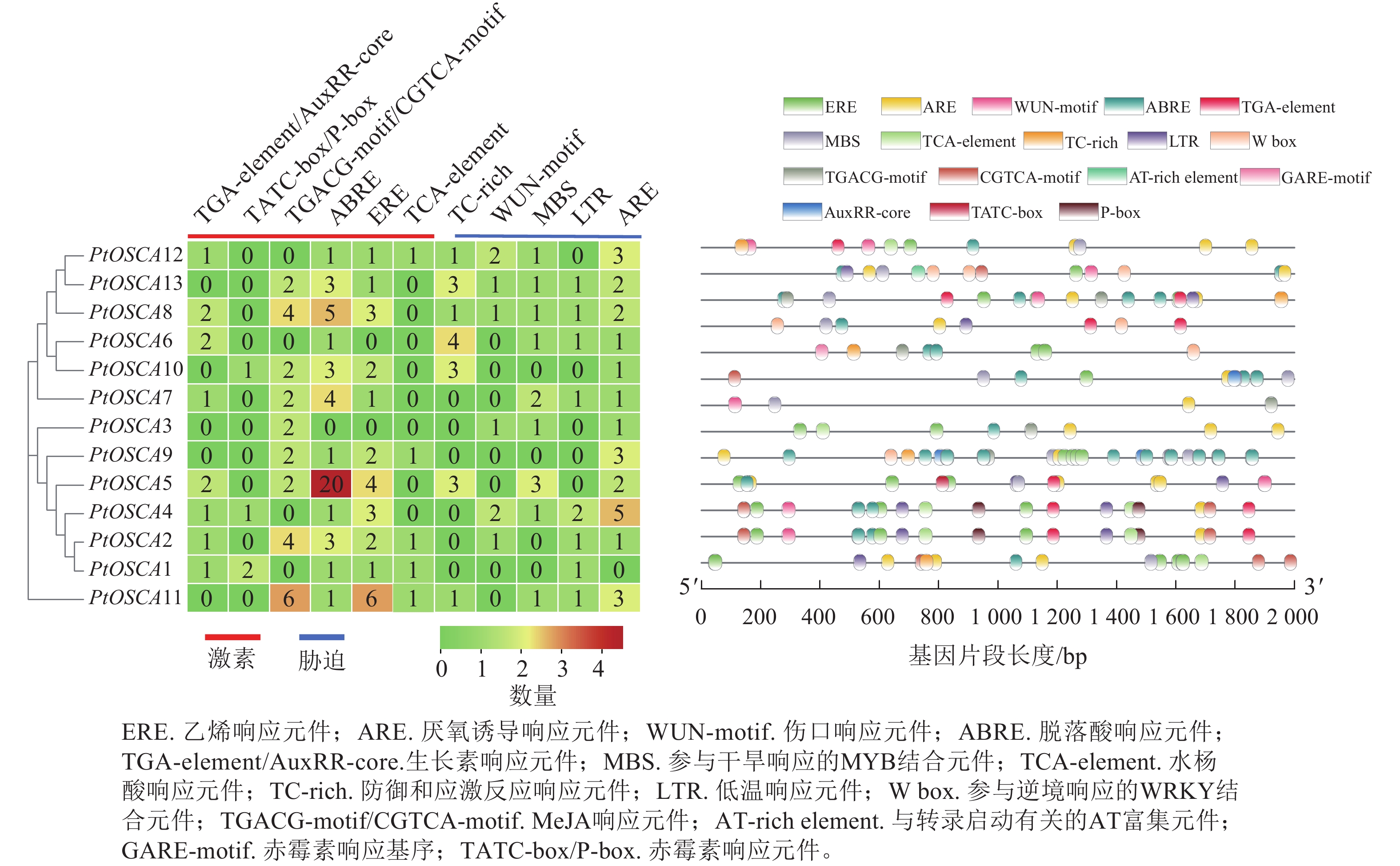
对13个PtOSCAs基因上游2 000 bp的序列启动子区域中,参与激素和非生物胁迫反应的顺式作用元件进行预测(图3),识别出6个胁迫相关的顺式作用元件,即干旱诱导性的MYB结合位点(MBS)、水杨酸响应元件(TCA-element)、厌氧诱导响应元件(ARE)、防御和应激反应响应顺式元件(TC-rich)、伤口响应元件(WUN-motif)和低温响应元件(LTR);此外,还包括5种激素相关顺式作用元件:生长素响应元件(TGA-element)、脱落酸响应元件(ABRE)、赤霉素响应元件(TATC-box/P-box)、茉莉酸甲酯(MeJA)响应元件(TGACG-motif/CGTCA-motif)、乙烯响应元件(ERE)等。在12个PtOSCAs基因启动子区域中,均可发现脱落酸响应的ABRE顺式作用元件以及厌氧诱导相关的ARE元件,8个PtOSCAs基因的启动子区域包含LTR元件,5个PtOSCAs基因启动子包含TC-rich元件,这些基因可能在植物对胁迫的应答和应激反应发挥作用。进一步分析发现:9个PtOSCAs基因启动子区域含有MBS元件,表明这些基因可能在干旱响应中发挥作用。由此,推测PtOSCAs基因在非生物胁迫响应中具有关键作用。
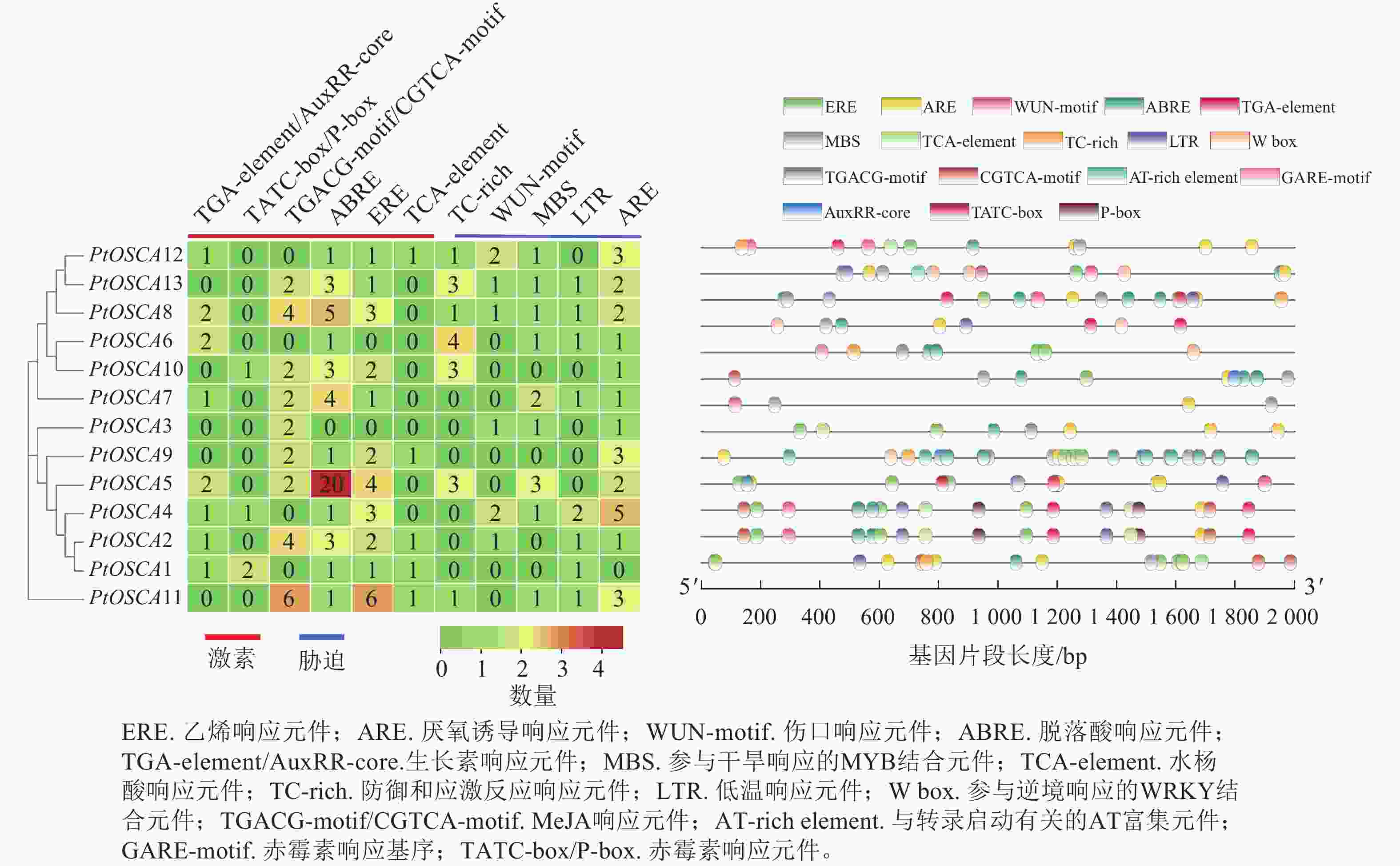
图 3 枳OSCA基因顺式作用元件
Figure 3. Cis-elements of OSCA genes in P. trifoliata
-
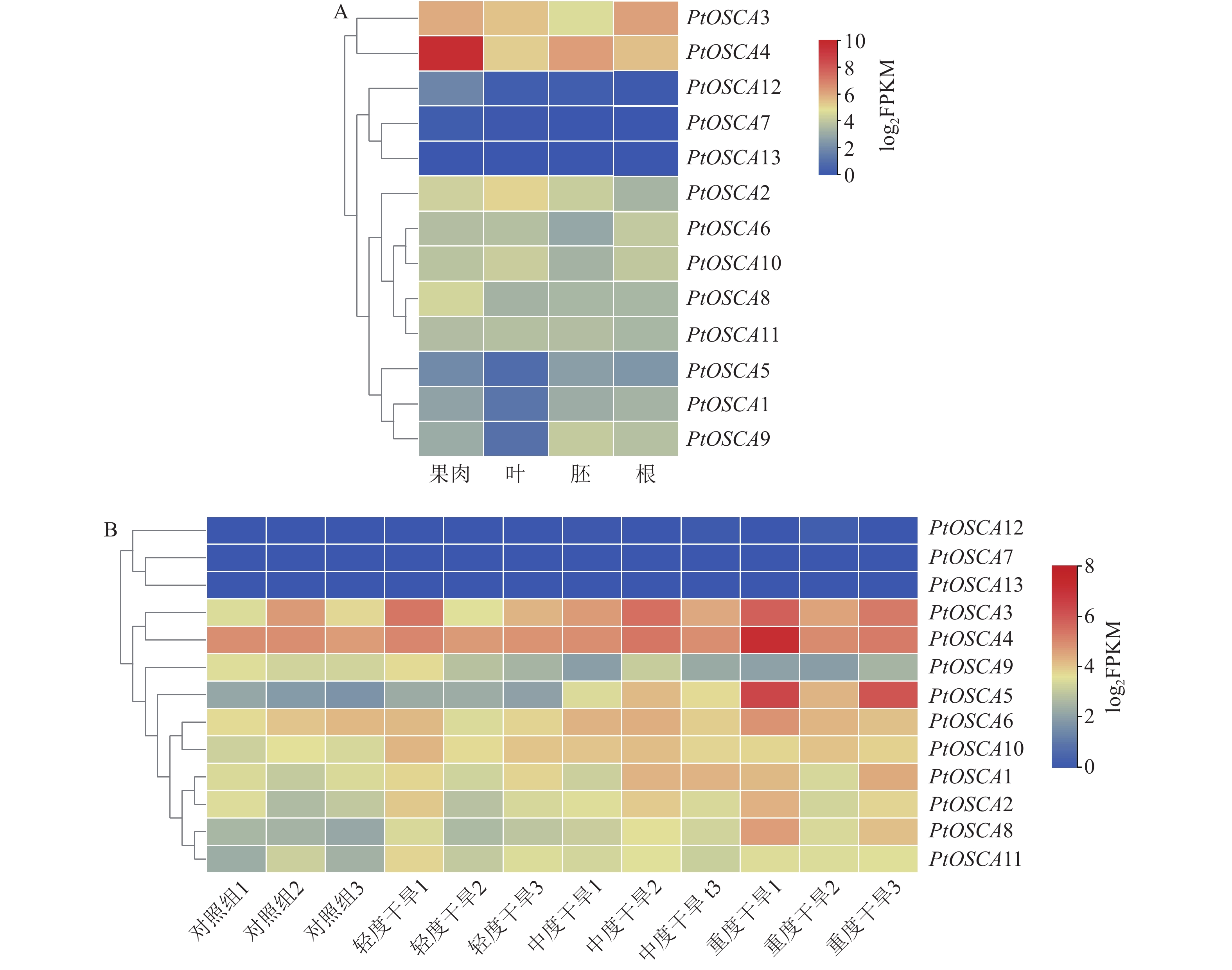
对枳果肉、叶、胚和根组织中13个PtOSCAs成员的相对表达水平进行分析,绘制枳不同组织中PtOSCA基因表达模式热图(图4A)。在各组织中均未检测到PtOSCA13表达量,其余12个基因在不同组织中表现出不同的表达模式。其中,PtOSCA3和PtOSCA4在各组织中表达水平均较高,表明其可能在枳生长发育阶段具有重要作用;PtOSCA12和PtOSCA7在各组织中表达水平较低;PtOSCA7在胚和根中无表达;PtOSCA11在各组织中的表达基本一致,不存在组织特异性表达。上述结果表明:PtOSCA基因在枳进化过程中发生了功能进化。
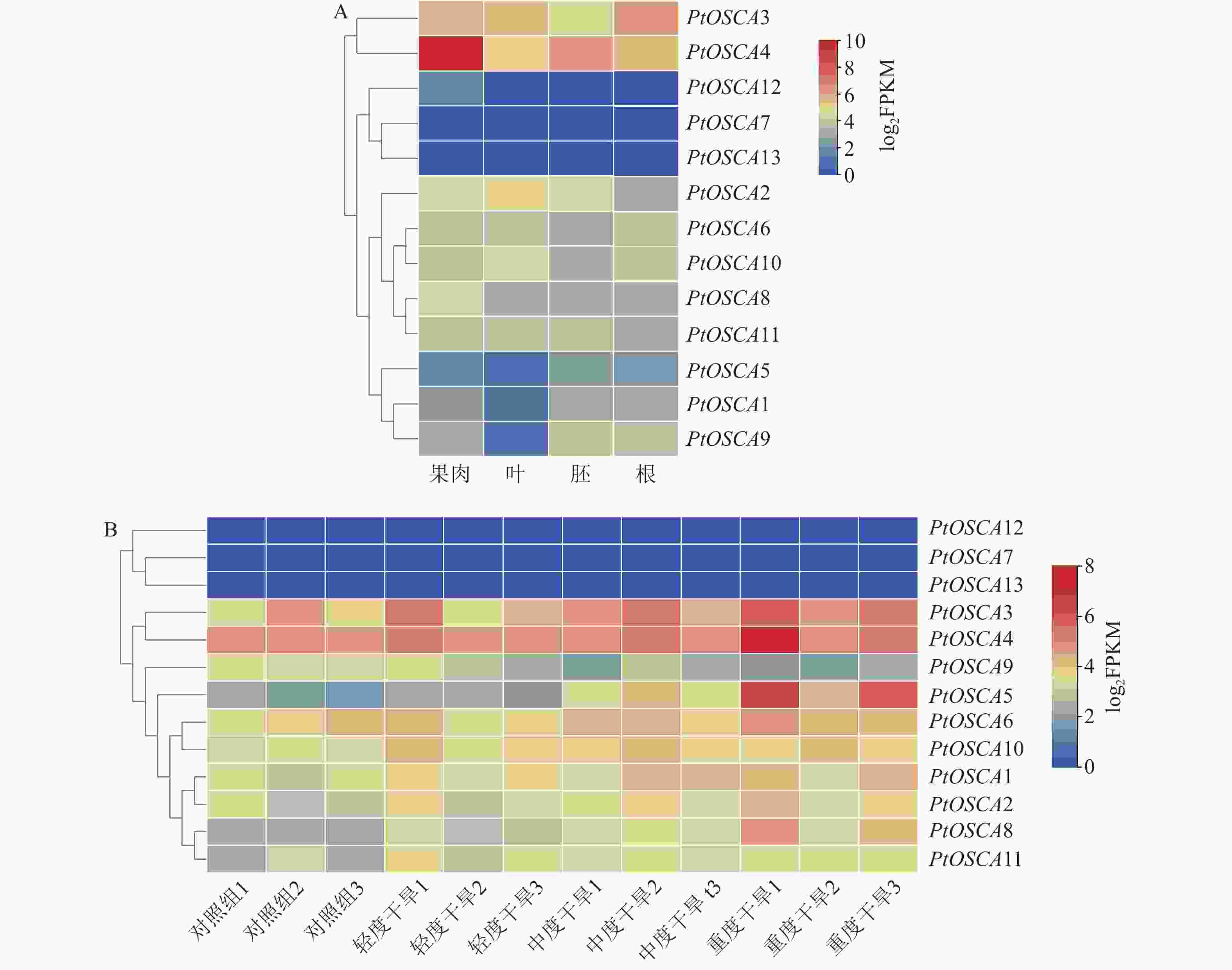
图 4 PtOSCA基因在不同组织中(A)及响应干旱胁迫(B)的表达模式
Figure 4. Expression patterns of PtOSCA in different tissues (A) and response to drought stress (B)
分析枳根组织在不同干旱胁迫程度下的转录组(图4B)。结果显示: PtOSCA13、PtOSCA7未检测到表达,PtOSCA12表达水平极低,其他PtOSCAs基因在轻度干旱、中度干旱和重度干旱胁迫下呈现显著的差异表达,其表达模式分为三类。第一类包括PtOSCA1~PtOSCA6、PtOSCA8,它们的表达量随着干旱程度的加剧而上升,且在各级胁迫组中的表达量均高于对照组(正常水分供应);第二类PtOSCA9则表现相反的趋势,随着干旱程度的增加,表达量下降,且在胁迫组中的表达量低于对照组;第三类基因PtOSCA10和PtOSCA11的表达量随着干旱程度的加剧保持平稳趋势,且在各胁迫组中的表达量均高于对照组。
-
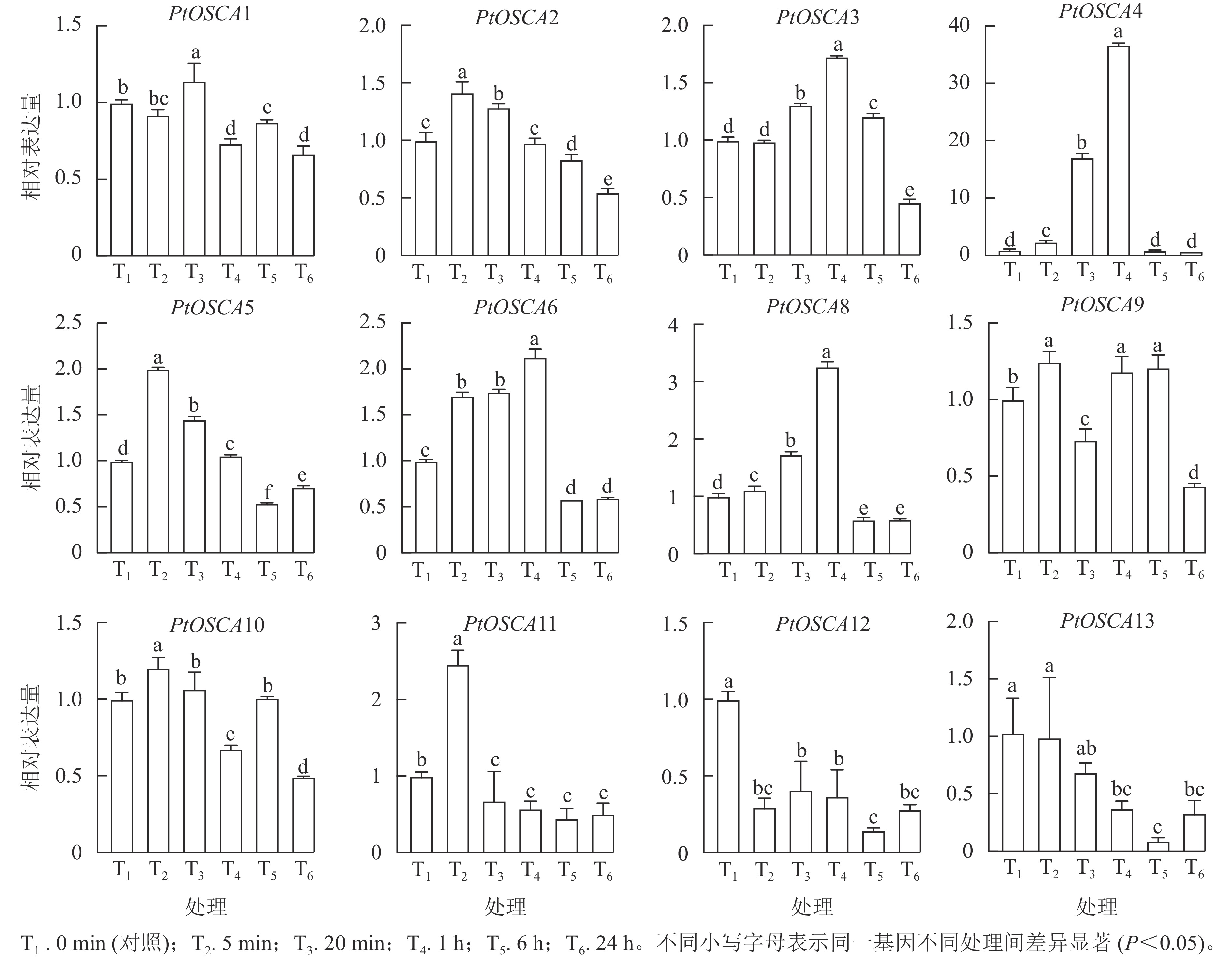
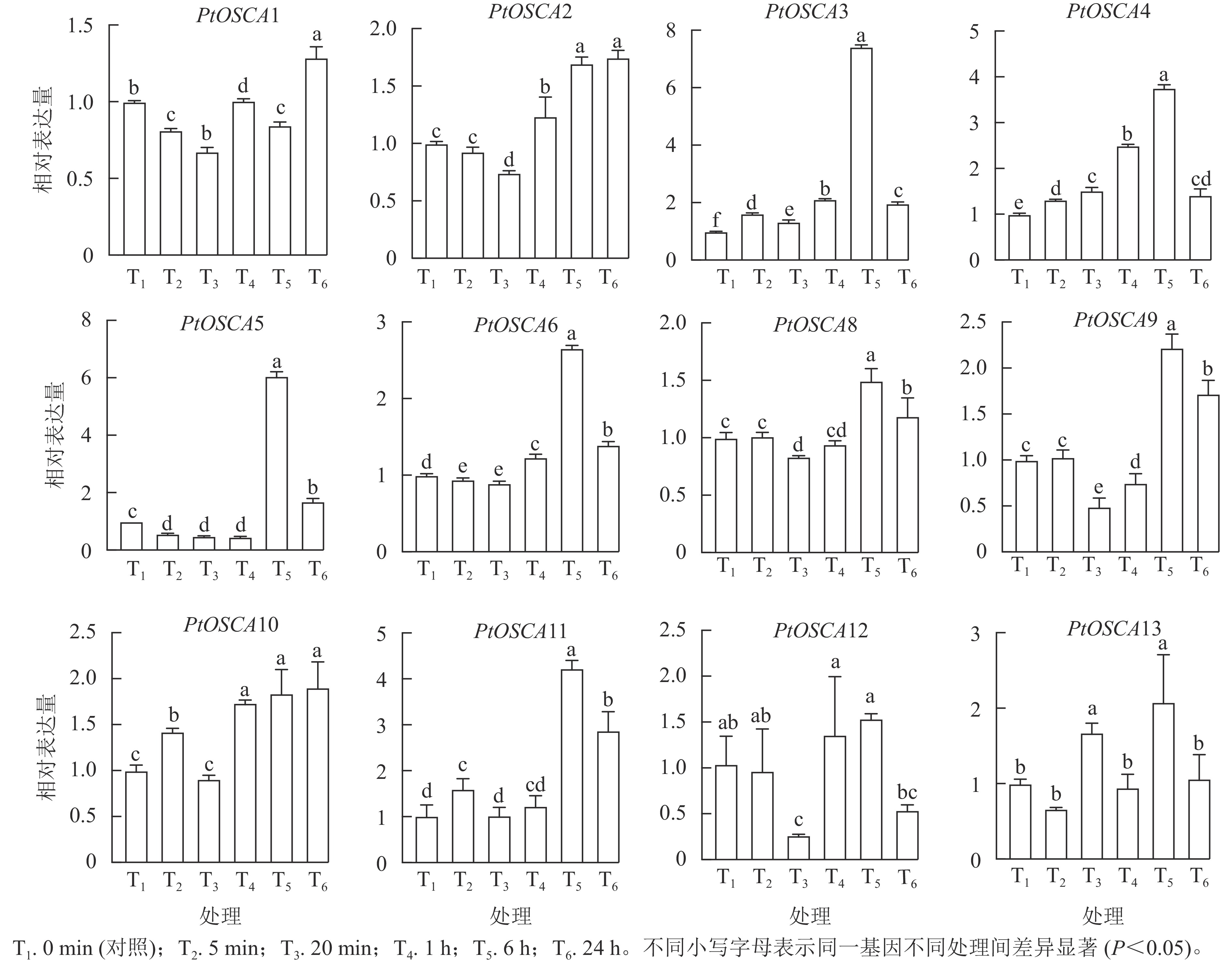
由图5和图6表明:PtOSCA7基因在根和叶组织中未检测到,其余PtOSCAs基因在枳的根和叶中均有表达,且大多数PtOSCAs基因能够响应干旱胁迫。在枳根组织中,与对照(0 min)相比,PtOSCA5和PtOSCA11在处理5 min时相对表达量显著上升,其相对表达量分别是对照的2.00和2.46倍;PEG处理1 h,PtOSCA4、PtOSCA6和PtOSCA8相对表达量显著上升,其中PtOSCA4增幅最明显,达对照的36.74倍;PtOSCA6和PtOSCA8相对表达量分别为对照的2.13和3.26倍;此外,PtOSCA1在处理20 min时达到最高相对表达量,是对照的1.14倍;而PtOSCA12和PtOSCA13随处理时间的延长基因相对表达量逐渐降低,在6 h时达到最低点。在叶片中,PtOSCA3~PtOSCA6、PtOSCA9、PtOSCA11和PtOSCA13在处理6 h时相对表达量显著上升,达到对照的7.43、3.76、6.06、2.66、2.22、4.23和2.08倍;PtOSCA12基因的相对表达量在24 h内呈现先下降后上升再下降的趋势,20 min时达到最低点,最高值出现在6 h时。综上,根部的大部分PtOSCAs基因在PEG处理1 h相对表达量达峰值,而在叶中则在PEG处理1 h后达到最高表达水平。与对照相比,干旱胁迫处理下PtOSCA4~PtOSCA6、PtOSCA11在根和叶中的表达显著上调,结果与转录组数据基本一致。
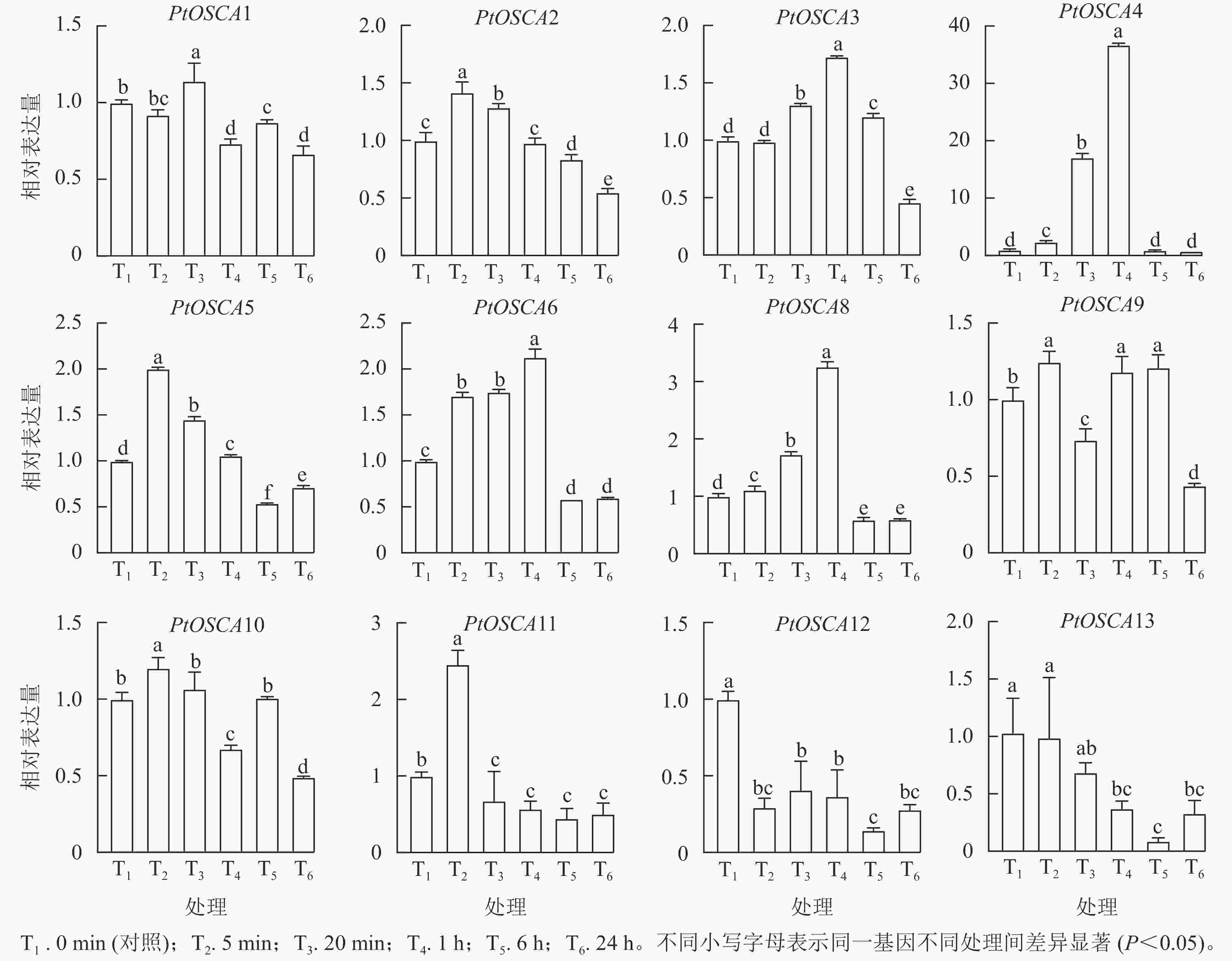
图 5 PEG处理下枳PtOSCAs在根中的表达模式
Figure 5. Expression patterns of PtOSCAs in roots under PEG treatment
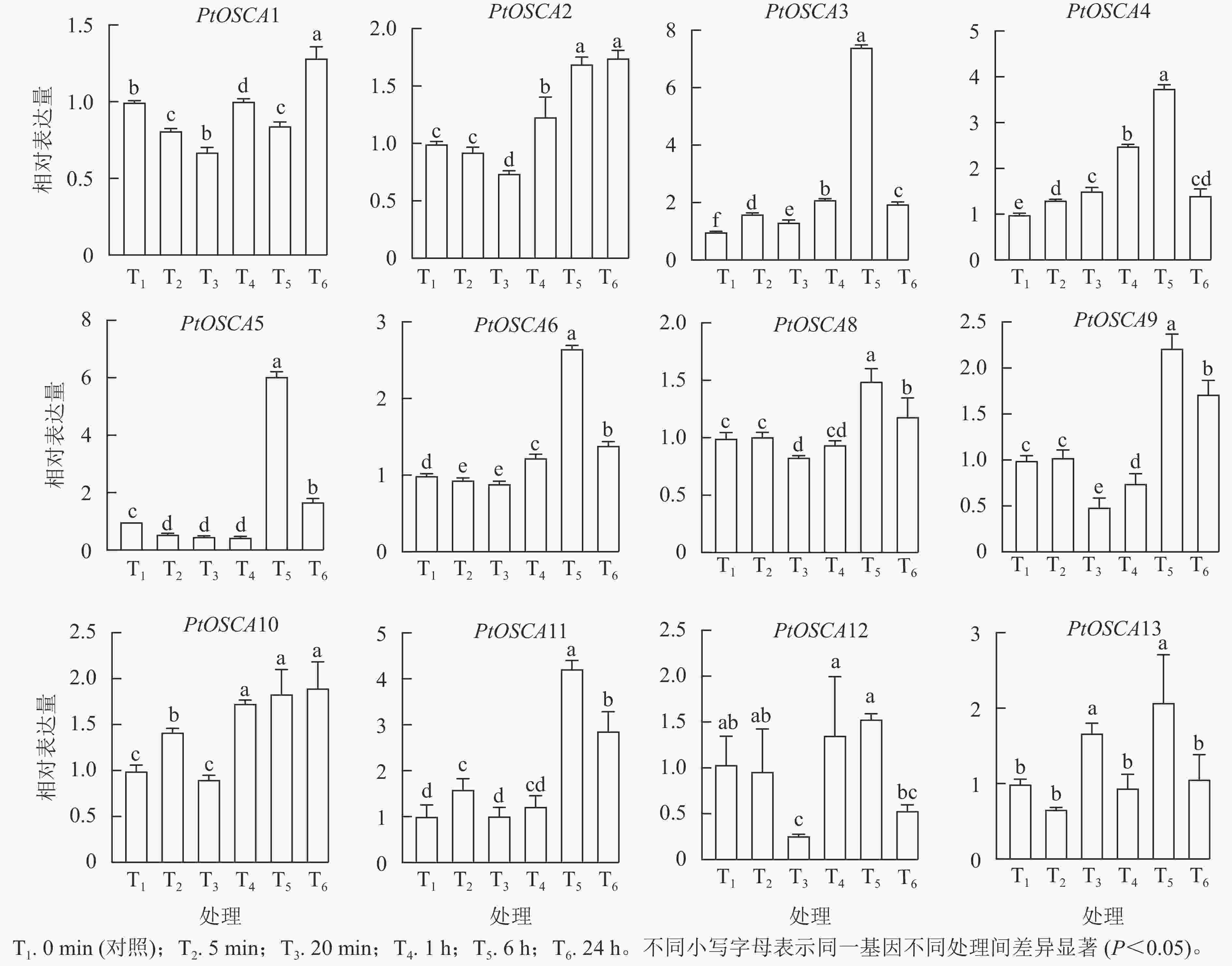
图 6 PEG处理下枳PtOSCAs在叶中的表达模式
Figure 6. Expression patterns of PtOSCAs in leaves under PEG treatment
-
植物在生长阶段持续面临高温干旱、低温、盐害等多种环境胁迫[18−19]。由于无法逃避这些有害环境,植物逐步形成了一套高效的信号传导系统和耐逆机制以应对环境变化。其中,OSCA基因在植物Ca2+信号转导途径中起着至关重要的作用[20]。本研究对枳OSCA基因进行了全面分析,共鉴定出13个PtOSCA基因家族成员。研究发现:PtOSCA蛋白的序列和理化特性差异很大,与其他植物的OSCA基因存在差别[3−4, 6, 21−22]。系统进化树表明:PtOSCA蛋白成员可分为4组,这一结果与拟南芥[3]、水稻[4]、玉米[6]、番茄[21]和猕猴桃Actinidia chinensis[22]等的研究结果一致。每个分支都包含来自拟南芥、枳和甜橙的OSCA成员,这表明OSCA家族的起源和多样化早于拟南芥、枳和甜橙的分化。OSCA家族的第Ⅲ和Ⅳ组各含有1个基因,它们在进化过程中未被淘汰,可能在植物进化中具有重要作用。此外,从进化树可以看出:枳与甜橙之间表现出较高的垂直同源基因相似性,这可能与它们均属于柑橘亚属,亲缘关系密切相关。OSCA基因家族在不同植物种类中的成员数量和分布差异可能是由基因组复制和功能分化导致的,这表明枳中鉴定到的13个PtOSCA基因可能是通过基因家族扩展和多样化来适应不同生态环境。
PtOSCAs基因保守结构域和基因结构分析显示:枳OSCA蛋白家族的所有分支成员都含有保守基序Motif 1和Motif 3,且这些保守基序在不同分支中的分布位置高度一致。在基因结构方面,枳OSCA家族的第Ⅰ、Ⅱ类基因内含子数量类似,而分别属于Ⅲ类和Ⅳ类的PtOSCA3和PtOSCA11内含子数量相对较少。已有研究表明:不同功能的基因往往经历不同程度的内含子获得和丢失,并且物种间共享内含子的位点随着进化距离的增加而减少,因此内含子的保守性可作为系统发育研究的重要标记,进化保守的基因更倾向于保留内含子[23]。此外,内含子较少或缺失的基因转录时间较短,能够在一定程度上提高基因的表达效率,从而实现快速响应[24]。特别是植物胁迫响应相关基因,如ShOSCA3基因在低温、干旱、ABA和灰霉菌 Botrytis cinerea胁迫下均表现出显著的表达变化,基因结构分析表明该基因不含内含子[21],这提示内含子数量可能与植物的环境适应性密切相关。本研究中PtOSCA11仅含有1个内含子,在PEG处理5 min时,其根部相对表达量较0 min时上调至2.46倍;而具有12个内含子的PtOSCA8在PEG处理6 h后,其根相对表达量较0 min时上调至3.26倍。尽管PtOSCAs基因的蛋白质结构和CDS序列内含子分布具有较高的保守性,但不同基因在胁迫响应中的表达模式仍存在显著差异,这种表达差异可能反映了PtOSCAs基因家族在进化过程中经历了功能分化。
对13个PtOSCAs基因的启动子组件进行分析,结果揭示了与主要激素(TGA-element、ABRE、TGACG-motif/CGTCA-motif、TATC-box/P-box和ERE)相关以及重要胁迫响应(ARE、MBS、LTR、TCA-element、 WUN-motil和TC-rich)相关的不同类型的核心组件。如除PtOSCA3外,所有基因都含有ABRE元件,这表明PtOSCA3在ABA响应基因表达中发挥至关重要的作用[25]。而且属于同一进化枝的PtOSCAs基因的启动子包含不同类型和数量的反应元件,因此,同一进化枝的不同基因可能表现出功能多样性,也可能具有不同的作用机制。在进化枝Ⅰ中,仅有PtOSCA4和PtOSCA5含有与干旱反应相关的MBS元件。在PEG处理下,PtOSCA4和PtOSCA5的相对表达量显著高于进化枝Ⅰ中的其他PtOSCAs,该结果与上述基因在干旱胁迫处理下的相对表达量和转录组数据的结果一致,这表明PtOSCA4和PtOSCA5可能在干旱胁迫响应中起到更重要的作用。枳PtOSCA基因家族启动子元件的分析显示:PtOSCA1、PtOSCA2、PtOSCA9、PtOSCA10基因的启动子区域未检测到干旱胁迫响应元件,不过在干旱胁迫下仍有一定的响应。在棉花中,GhOSCA1.1、GhOSCA1.9、GhOSCA1.14和GhOSCA2.12没有盐碱胁迫响应元件,但它们在盐碱胁迫条件下表达量显著上调[26]。这些结果说明:基因对胁迫的响应机制可能不完全依赖于启动子区域中的特定顺式作用元件。当植物受到特定胁迫时,其他胁迫响应元件可能参与调控,体现出不同顺式作用元件潜在的协同作用,进而增强对胁迫的应答能力。
前人研究表明:水稻多个OsOSCAs基因对盐胁迫、干旱胁迫及脱落酸处理作出响应,其中至少8个基因在单一渗透胁迫条件下显著上调,如OsOSCA1.1和OsOSCA3.1在PEG处理下的相对表达量可增加至40倍[4]。玉米中ZmOSCA2.3和ZmOSCA2.4可响应PEG处理、盐胁迫和脱落酸处理,其相对表达量增加了200倍以上且与脯氨酸含量一致,将ZmOSCA2.4在拟南芥中过表达可显著提高拟南芥的抗旱能力[6]。本研究通过分析PtOSCA的动态渗透胁迫响应表达模式,发现枳OSCA基因家族响应干旱胁迫,推测这可能与逆境调节相关。利用RT-qPCR检测了PtOSCA基因家族在PEG胁迫处理下相对表达量,结果显示:几乎所有枳OSCAs基因均对干旱胁迫有所响应。其中PtOSCA4~PtOSCA6、PtOSCA11在干旱胁迫处理下,根和叶组织中的表达上调非常明显,这一结果与转录组数据基本一致。根中大部分PtOSCAs基因在PEG处理1 h内达到最高表达量,而叶中则在1 h后达到最高。植物依靠植物顶端产生的蒸腾压力和根部产生的根压来吸收水分,从而促使水分从根部向上运动,PtOSCAs基因的不同表达模式符合水分由地下部运往地上部的过程时间差异。与其他PtOSCA家族成员相比,PtOSCA4基因对干旱胁迫的响应最为显著,在根组织中1 h时的表达量是0 min时的36.74倍,因此在响应干旱胁迫过程中,PtOSCA4可能发挥了关键作用,由此推测PtOSCA4在干旱胁迫的响应通路中可能涉及一定的相互作用。已有研究表明:AtOSCA1.1 (At4g04340)是植株响应渗透胁迫的关键基因,其编码的蛋白定位于根和叶细胞膜,充当离子通道,干旱胁迫下促进大量Ca2+进入细胞[27];AtOSCA1.2 (At4g
22120 )与AtOSCA1.1有较高同源性,在调节Ca2+的渗透过程中起到重要作用,该蛋白在高渗透压条件下能提升细胞内Ca2+的含量[8]。在本研究中,PtOSCA4与AtOSCA1.1和AtOSCA1.2具有较高的序列相似性,均属于进化树分支Ⅰ,且在干旱胁迫下均能表现出强烈的响应。因此,PtOSCA4可能与AtOSCA1.1、AtOSCA1.2在功能上存在一定的相似性,推测PtOSCA4可能对渗透压较敏感且在抗旱中具有重要作用。 -
本研究基于全基因组数据对枳OSCA基因家族进行了全面分析,共鉴定出13个枳OSCAs基因,并通过系统进化分析将它们划分为4个分支。枳OSCA基因家族的不同成员在各组织中对干旱胁迫的响应存在差异;枳OSCA基因在进化过程中具有一定的保守性,与拟南芥和甜橙有较近的亲缘关系;PtOSCA4可能在响应干旱胁迫机制中发挥重要作用。本研究为深入解析枳OSCA基因的生物学功能及在干旱响应中的分子机制提供了重要的理论依据。
Genome-wide identification and drought stress response analysis of the OSCA gene family in Poncirus trifoliata
-
摘要:
目的 揭示OSCA基因家族在枳Poncirus trifoliata中的结构特征及在干旱胁迫下的表达特征,为PtOSCA基因功能分析与枳抗旱性研究提供理论依据。 方法 利用生物信息学方法对全基因组范围内的枳OSCA基因家族成员进行鉴定和综合分析,通过实时荧光定量PCR (RT-qPCR)分析PtOSCA在聚乙二醇(PEG)处理下的动态表达特征。 结果 在枳基因组中共鉴定出13个PtOSCAs基因,并根据其与拟南芥Arabidopsis thaliana OSCA基因家族成员的同源性命名为PtOSCA1~PtOSCA13。染色体定位分析表明:PtOSCAs不均等地分布在枳18条染色体中的7条染色体上。系统发育分析显示:PtOSCAs可分为4组,分别包含5、6、1、1个家族成员。蛋白质理化性质和保守结构域分析表明:PtOSCAs之间的蛋白长度和分子质量存在差异,但蛋白基序较为保守且均含有Motif 1和Motif 3。顺式作用元件分析显示:PtOSCAs基因的启动子区域存在大量响应干旱、脱落酸、厌氧诱导、低温胁迫等顺式作用元件。基因表达分析表明:PtOSCAs基因表达具有组织特异性和干旱胁迫响应特征,干旱胁迫下,PtOSCA基因家族各成员在叶和根中的表达量均有不同程度变化,叶中11个PtOSCAs基因和根中10个PtOSCAs基因受干旱胁迫诱导上调表达,其中PtOSCA4在叶和根中的表达量分别上调了3.76和36.74倍。 结论 PtOSCA基因在进化过程中表现出一定的保守性,并与拟南芥和甜橙Citrus sinensis OSCAs基因呈现较近的亲缘关系。PtOSCA基因家族内部存在功能差异,其中,PtOSCA4可能在干旱胁迫响应机制中发挥着关键作用。图6表2参27 Abstract:Objective This study aims to reveal the structural characteristics of the OSCA gene family in Poncirus trifoliata and its expression profile under drought stress, providing a reference for PtOSCA gene functional analysis and drought resistance research. Method Bioinformatic methods were used to identify and comprehensively analyze PtOSCA gene family members across the whole genome. RT-qPCR was used to analyze the dynamic expression profiles of PtOSCA genes under PEG treatment. Result Thirteen PtOSCAs were identified in the P. trifoliata genome and named PtOSCA1−PtOSCA13 according to their homology with Arabidopsis thaliana OSCA genes. Chromosomal localization analysis indicated that these genes were unevenly distributed across 7 out of the 18 chromosomes in P. trifoliata. Phylogenetic analysis showed that PtOSCAs were divided into four groups, with 5, 6, 1, and 1 family members respectively. Analysis of protein physicochemical properties and conserved domains revealed that there were differences in protein length and molecular weight among PtOSCAs, while the motifs were relatively conserved, with all PtOSCAs proteins containing Motif 1 and Motif 3. Cis-acting element analysis indicated that the promoter regions of PtOSCAs had numerous cis-elements responsive to drought, abscisic acid, anaerobic induction, and low-temperature stress. Gene expression analysis demonstrated that PtOSCAs had tissue-specific expression and responses to drought stress. Under drought stress, the expression levels of PtOSCA family members changed to varying degrees in both leaves and roots. In leaves, 11 PtOSCAs were induced by drought stress,and in roots, 10 PtOSCAs were upregulated, with PtOSCA4 upregulated 3.76-fold in leaves and 36.74-fold in roots after drought stress induction. Conclusion The PtOSCA gene family shows evolutionary conservation and shares a close phylogenetic relationship with the OSCA genes of A.thaliana and Citrus sinensis. Functional differentiation exists within the PtOSCA gene family, and PtOSCA4 may play a key role in drought stress response of P. trifoliata. The results of this study provide an important basis for research on the molecular functions of PtOSCAs under drought stress in P. trifoliata. [Ch, 6 fig. 2 tab. 27 ref.] -
Key words:
- Poncirus trifoliata /
- Citrus /
- OSCA /
- drought stress /
- expression analysis
-
图 1 枳、甜橙、拟南芥的OSCA基因编码产物系统进化树
Figure 1 Phylogenetic tree of the coding products of the OSCA gene in P. trifoliata,C. sinensis and A.thaliana
图 2 枳OSCA家族成员蛋白基序(A)和基因结构(B)分析
Figure 2 Analysis of conserved domains (A) and gene structure (B) of OSCA family members in P. trifoliata
图 4 PtOSCA基因在不同组织中(A)及响应干旱胁迫(B)的表达模式
Figure 4 Expression patterns of PtOSCA in different tissues (A) and response to drought stress (B)
图 5 PEG处理下枳PtOSCAs在根中的表达模式
Figure 5 Expression patterns of PtOSCAs in roots under PEG treatment
图 6 PEG处理下枳PtOSCAs在叶中的表达模式
Figure 6 Expression patterns of PtOSCAs in leaves under PEG treatment
表 1 实时荧光定量PCR引物
Table 1. Primers for quantitative real-time PCR analysis
基因名称 正向引物(5′→3′) 反向引物(5′→3′) PtOSCA1 TGGAACGAGCAAGGGAACCA TATGTTTGCTTGGCAACGGAG PtOSCA2 GCACACCGGTGCATTTGTAA ACAGCAGAATCCAATCCCCC PtOSCA3 CAGAGGGAACAAGACCGACC GCAGAAGCTGCAGTTACCCT PtOSCA4 AGACTGAAAAGGACAGGGCG GGGTGTCACTGTGGCATACA PtOSCA5 CCAAACCCACCGAGTCAAGT AGCAGAGCATGTGAAGGTCC PtOSCA6 GGAGCTGCAATTGCTTTCCATT TAAGGCCTTGCACAACCAGC PtOSCA7 CTTCCGTGGCCACATTTCCA ACTTTCTCGGACACTGCGTT PtOSCA8 TGGATCAACAAGATGAGCTGGA CCAATCTCTGCCGTCTCTGC PtOSCA9 TTTGGAACGGAGGACGTCAG AATGCCAGCCCATCCATCAA PtOSCA10 TTCTTCAGTTGTTCCTATCATTCG CCACCAGACCTTTGAACAAGC PtOSCA11 CCGCTCAGTTCCTCCTCATC ACTCCATGCATTCCTTTTTCAAT PtOSCA12 ACAGTCGCATTCGGTTTCAC CCCTGATGATCCTTATGTCCAGA PtOSCA13 AAGGCTTGACTCGACTTCACC TTGCCCCATAATGACCAGCA PtActin CCGACCGTATGAGCAAGGAAA TTCCTGTGGACAATGGATGGA  下载: 导出CSV
下载: 导出CSV
表 2 枳OSCA基因家族信息
Table 2. Information on OSCA gene family in P. trifoliata
基因名称 基因 ID 基因全长/bp 氨基酸数量/个 等电点 分子量/Da 染色体 亚细胞定位 PtOSCA1 Pt2g013910.1 2 280 759 9.20 86 802.76 2 细胞膜 PtOSCA2 Pt2g013930.1 2 280 759 8.96 86 861.76 2 细胞膜,叶绿体 PtOSCA3 Pt2g023140.1 2 181 726 9.17 81 688.79 2 细胞膜 PtOSCA4 Pt2g028480.4 2 319 772 9.30 88 145.30 2 细胞膜,细胞壁,叶绿体 PtOSCA5 Pt3g032590.1 3 006 758 9.08 86 482.77 3 细胞膜,细胞壁,叶绿体 PtOSCA6 Pt4g000270.1 2 127 708 8.74 80 821.35 4 细胞膜 PtOSCA7 Pt4g001040.1 2 103 700 8.85 81 011.45 4 细胞膜 PtOSCA8 Pt5g011440.2 2 229 742 8.50 83 798.96 5 细胞膜 PtOSCA9 Pt5g018480.1 2 424 807 9.52 93 051.95 5 细胞膜,叶绿体 PtOSCA10 Pt6g003200.3 4 494 682 8.86 77 495.58 6 细胞膜 PtOSCA11 Pt7g021120.1 2 424 786 6.75 89 331.88 7 细胞膜 PtOSCA12 PtUn012370.2 2 139 712 8.38 80 420.82 未知 细胞膜 PtOSCA13 PtUn037020.1 3 889 409 9.23 46 921.00 未知 细胞膜
下载: 导出CSV
-
[1] 钟灶发, 张利娟, 高思思, 等. 干旱胁迫下4种柑橘砧木叶片细胞学特征及抗旱性比较[J]. 园艺学报, 2021, 48(8): 1579−1588. ZHONG Zaofa, ZHANG Lijuan, GAO Sisi, et al. Leaf cytological characteristics and resistance comparison of four citrus rootstocks under drought stress [J]. Acta Horticulturae Sinica, 2021, 48(8): 1579−1588. [2] 刘晓纳, 徐媛媛, 朱世平, 等. 不同柑橘砧木的耐旱性评价[J]. 果树学报, 2016, 33(10): 1230−1240. LIU Xiaona, XU Yuanyuan, ZHU Shiping, et al. Evaluation of drought tolerance in different citrus rootstocks [J]. Journal of Fruit Science, 2016, 33(10): 1230−1240. [3] YUAN Fang, YANG Huimin, XUE Yan, et al. OSCA1 mediates osmotic-stress-evoked Ca2+ increases vital for osmosensing in Arabidopsis [J]. Nature, 2014, 514(7522): 367−371. [4] LI Yunshuang, YUAN Fang, WEN Zhaohong, et al. Genome-wide survey and expression analysis of the OSCA gene family in rice[J/OL]. BMC Plant Biology, 2015, 15(1): 261[2024-12-26]. DOI: 10.1186/s12870-015-0653-8. [5] YIN Lili, ZHANG Meiling, WU Ruigang, et al. Genome-wide analysis of OSCA gene family members in Vigna radiata and their involvement in the osmotic response[J/OL]. BMC Plant Biology, 2021, 21(1): 408[2024-12-26]. DOI: 10.1186/S12870-021-03184-2. [6] CAO Liru, ZHANG Pengyu, LU Xiaomin, et al. Systematic analysis of the maize OSCA genes revealing ZmOSCA family members involved in osmotic stress and ZmOSCA2.4 confers enhanced drought tolerance in transgenic Arabidopsis[J/OL]. International Journal of Molecular Sciences, 2020, 21(1): 351[2024-12-26]. DOI: 10.3390/ijms21010351. [7] LIU Xin, WANG Jiawei, SUN Linfeng. Structure of the hyperosmolality-gated calcium-permeable channel OSCA1.2[J/OL]. Nature Communications, 2018, 9: 5060[2024-12-26]. DOI: 10.1038/s41467-018-07564-5. [8] HOU Congcong, TIAN Wang, KLEIST T, et al. DUF221 proteins are a family of osmosensitive calcium-permeable cation channels conserved across eukaryotes [J]. Cell Research, 2014, 24(5): 632−635. [9] 王傲雪, 张可为, 张瑶, 等. 番茄OSCA基因家族鉴定及不同胁迫条件下表达分析[J]. 东北农业大学学报, 2019, 50(1): 19−28. WANG Aoxue, ZHANG Kewei, ZHANG Yao, et al. Identification of tomato OSCA gene family and expression analysis under different stress conditions [J]. Journal of Northeast Agricultural University, 2019, 50(1): 19−28. [10] WEI Tonglu, WANG Yue, XIE Zongzhou, et al. Enhanced ROS scavenging and sugar accumulation contribute to drought tolerance of naturally occurring autotetraploids in Poncirus trifoliata [J]. Plant Biotechnology Journal, 2019, 17(7): 1394−1407. [11] 黄奕孜, 钱旺, 邱姗, 等. 光皮桦AP2/ERF基因家族鉴定与表达分析[J]. 浙江农林大学学报, 2022, 39(6): 1183−1193. HUANG Yizi, QIAN Wang, QIU Shan, et al. Identification and expression analysis of AP2/ERF gene family of Betula luminifera [J]. Journal of Zhejiang A&F University, 2022, 39(6): 1183−1193. [12] 张瑜, 吴才君, 苏文桢, 等. 西瓜OSCA基因家族全基因组鉴定及胁迫响应分析[J]. 南方农业学报, 2021, 52(12): 3330−3339. ZHANG Yu, WU Caijun, SU Wenzhen, et al. Genome-wide identification and stress response analysis of OSCA gene family in watermelon [J]. Journal of Southern Agriculture, 2021, 52(12): 3330−3339. [13] TIAN Feng, YANG Dechang, MENG Yuqi, et al. PlantRegMap: charting functional regulatory maps in plants [J]. Nucleic Acids Research, 2020, 48(D1): D1104−D1113. [14] LESCOT M. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences[J]. Nucleic Acids Research, 30(1): 325−327. [15] 崔红丽, 孙明哲, 贾博为, 等. 蒺藜苜蓿OSCA基因家族鉴定及低温逆境表达分析[J]. 草业学报, 2024, 33(9): 111−125. CUI Hongli, SUN Mingzhe, JIA Bowei, et al. Genome-wide analysis and expression of the OSCA family genes from Medicago truncatula in response to low temperature stresses [J]. Acta Prataculturae Sinica, 2024, 33(9): 111−125. [16] SHE Kuijun, PAN Wenqiu, YAN Ying, et al. Genome-wide identification, evolution and expressional analysis of OSCA gene family in barley (Hordeum vulgare L. )[J/OL]. International Journal of Molecular Sciences, 2022, 23(21): 13027[2024-12-26]. DOI: 10.3390/ijms232113027. [17] 陈梦瑶, 胡怡然, 郑志富, 等. 大豆IGT基因家族的全基因组鉴定及组织表达分析[J]. 浙江农林大学学报, 2025, 42(1): 64−73. CHEN Mengyao, HU Yiran, ZHENG Zhifu, et al. Genome-wide identification and tissue expression analysis of IGT gene family in soybean [J]. Journal of Zhejiang A&F University, 2025, 42(1): 64−73. [18] GUO Meng, LIU Jinhong, LU Jinping, et al. Genome-wide analysis of the CaHsp20 gene family in pepper: comprehensive sequence and expression profile analysis under heat stress [J/OL]. Frontiers in Plant Science, 2015, 6: 806[2024-12-26]. DOI: 10.3389/fpls.2015.00806. [19] CHEN Xuexue, DING Yanglin, YANG Yongqing, et al. Protein kinases in plant responses to drought, salt, and cold stress [J]. Journal of Integrative Plant Biology, 2021, 63(1): 53−78. [20] SU Ruiping, WANG Yuning, CUI Ping, et al. Isolation of OSCAs in wheat and over-expression of TaOSCA14D increased salt stress tolerance [J/OL]. Journal of Plant Physiology, 2025, 306: 154449[2024-12-26]. DOI: 10.1016/j.jplph.2025.154449. [21] MIAO Shuang, LI Fengshuo, HAN Yang, et al. Identification of OSCA gene family in Solanum habrochaites and its function analysis under stress[J/OL]. BMC Genomics, 2022, 23(1): 547[2024-12-26]. DOI: 10.1186/s12864-022-08675-6. [22] 徐子怡, 罗晨宇, 占坤, 等. 猕猴桃OSCA基因家族鉴定及其在非生物胁迫下的表达分析[J]. 果树学报, 2024, 41(3): 436−447. XU Ziyi, LUO Chenyu, ZHAN Kun, et al. Genome-wide identification of OSCA gene family members and their expression analysis under different abiotic stresses in kiwifruit [J]. Journal of Fruit Sciences, 2024, 41(3): 436−447. [23] 曹军. 真核生物内含子研究进展[J]. 生物过程, 2011, 1: 9−12. CAO Jun. Research progress in eukaryotic intron [J]. Bioprocess, 2011, 1: 9−12. [24] ZHAO Peng, WANG Dongdong, WANG Ruoqiu, et al. Genome-wide analysis of the potato Hsp20 gene family: identification, genomic organization and expression profiles in response to heat stress[J/OL]. BMC Genomics, 2018, 19(1): 61[2024-12-26]. DOI: 10.1186/s12864-018-4443-1. [25] JI Lexiang, WANG Jia, YE Meixia, et al. Identification and characterization of the Populus AREB/ABF subfamily [J]. Journal of Integrative Plant Biology, 2013, 55(2): 177−186. [26] ABBASI H, JAMIL M, HAQ A, et al. Salt stress manifestation on plants, mechanism of salt tolerance and potassium role in alleviating it: a review [J]. Zemdirbyste-Agriculture, 2016, 103(2): 229−238. [27] ZHANG Heng, ZHAO Yang, ZHU Jiankang. Thriving under stress: how plants balance growth and the stress response [J]. Developmental Cell, 2020, 55(5): 529−543. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20250103
 点击查看大图
点击查看大图
计量
- 文章访问数: 2127
- HTML全文浏览量: 644
- PDF下载量: 90
- 被引次数: 0
