




-
森林土壤的物理、化学及生物学性质综合表现为土壤肥力。土壤肥力的高低影响着森林植物的分布、生长以及群落的组成、结构和生产力,植物又通过凋落物、根系分泌物等反作用于土壤,改变着土壤的物理、化学及生物学性质,由此形成一个动态的相互作用机制,提升森林生态系统结构与功能[1-3]
毛竹Phyllostachys edulis属于禾本科Gramineae刚竹属Phyllostachys植物,原产于中国亚热带地区,以地下竹鞭进行无性繁殖,具有扩展蔓延速度快和竞争力强的特点。从20世纪90年代开始,毛竹因其具有良好的竹材、竹芛等多种用途而作为笋材两用的经济树种在中国亚热带地区大面积种植,为山区农民脱贫致富做出贡献[4]。随着中国经济的发展,毛竹林经营成本不断提高,而竹材价格反而降低,因此,很多竹农已经放弃了对毛竹林的经营管理,导致失去管理的毛竹林无序扩张蔓延。近年来,这种趋势不断加剧。在许多地方由于毛竹地下竹鞭扩张渗透到阔叶林等相邻森林中,迅速成林并侵占其他物种的生境,形成毛竹单优势群落,导致植物种类减少,群落层次结构简化,生物多样性显著降低,土壤物理性质、化学性质和土壤微生物发生显著变化,整个区域生态系统退化[5-9],威胁到生物多样性保护和区域生态安全。因此,控制毛竹林无序蔓延扩张,开展毛竹林皆伐后生态恢复效果研究,已受到全社会广泛关注。2015年,为控制和减少毛竹入侵对天目山国家级自然保护区生态破坏,经审批,对该区域部分入侵毛竹林进行皆伐。本研究在毛竹林皆伐区和未伐区设置样地进行比较研究,探讨毛竹林皆伐后土壤自然恢复效果,为天目山毛竹林入侵区域生态修复提供技术支持,为亚热带毛竹林入侵区生态修复提供借鉴。
-
研究区位于浙江省杭州市临安区天目山国家级自然保护区,地处中亚热带北缘,浙皖两省交界处,是南北植物汇流之区,也是中国最主要的野生植物类型自然保护区之一,面积为4 300 hm2,海拔300~1 556 m。研究区土壤类型随着海拔变化具有较大差异,600 m以下为红壤,600~850 m主要为黄红壤,850~1 200 m为黄壤,1 200 m以上为黄棕壤。植被分布也有着明显的垂直带谱特征:在海拔230~850 m分布常绿阔叶林,850~1 100 m分布常绿落叶阔叶混交林,1 100~1 350 m分布落叶阔叶林,1 400 m以上为落叶矮林。
在天目山国家级自然保护区,毛竹林作为一种特殊的森林类型,主要分布在海拔350~900 m。由于保护区不允许开展森林经营活动,毛竹林完全处于自然状态,立竹度高,并不断向周边蔓延扩张,面积从55 hm2蔓延扩张到105 hm2,对保护区原有植被蚕食的现象日益严重。为控制和减少毛竹入侵对天目山国家级自然保护区的生态破坏,促进生物多样性保护。2015年经上级批准,对部分入侵毛竹林进行皆伐作业,并于皆伐后连续3 a进行人工除笋,防止毛竹复占。毛竹林皆伐后经过5 a自然恢复,物种组成有了明显的变化,且物种种类明显增加,与未皆伐毛竹林相比植物多样性有显著增加[10-11]。
-
于2017年在毛竹林皆伐区域设置了清除(CR)和保留(UR)采伐剩余物样地各3个,在未伐区毛竹林内设置对照样地(ck) 3个。样地面积20 m×20 m,在每个样地四角和中间分别设置1个5 m×5 m样方。
在2020年11月样地复查的同时采集土壤样品,每个样地内采用5点取样法,均匀采取0~10、10~20 cm土层中3个点位土壤的混合样品,并过2 mm筛,去除样品中的植物碎屑及石块,经过研磨过筛处理后测定土壤化学性质;使用200 cm3环刀于样方中心采集0~10、10~20 cm土层土壤用于测定土壤物理性质;新鲜土壤样品保存在4 ℃环境中用于测定土壤胞外酶活性。
-
土壤理化性质参考《土壤农业化学分析方法》进行测定[12]。土壤容重、孔隙度、最大持水量、最小持水量、毛管持水量及含水量采用环刀法测定。土壤pH采用pH计测定(水土质量比为2.5∶1.0);土壤有机质采用高温外热重铬酸钾氧化-容量法测定;全氮用凯氏定氮法测定;全磷采用氢氧化钠熔融法-钼锑抗比色测定;全钾采用氢氧化钠熔融法测定;碱解氮采用碱解扩散法测定;有效磷采用盐酸-氟化铵浸提,钼锑抗比色法测定;速效钾采用火焰光度计测定。土壤胞外酶活性参考GERMAN等[13]使用鲜土进行测定,脲酶采用比色法测定,水解酶活性使用甲基伞形酮(4-MUB)作为底物标示,氧化酶活性使用L-二羟苯丙氨酸(L-DOPA)作为底物标示。利用微孔板荧光法、使用多功能酶标仪测定单位时间内特定底物被各种酶催化释放出的荧光产物的量来计算酶活性。
-
采用 Excel 2019 进行数据整理;用 SPSS 26.0 进行统计分析。对毛竹林样地和砍伐后自然恢复迹地土壤物理、化学性质及胞外酶活性差异做单因素方差分析(one-way ANOVA),显著性水平设置为0.05,并采用Tukey 方法进行显著性检验。统计结果用Origin 2018制图。
-
综合考虑土壤物理性质、化学性质及胞外酶活性,本研究选取了34 个候选指标,分别是:0~10、10~20 cm土层土壤容重、孔隙度、毛管持水量、田间持水量、饱和持水量、pH、有机质质量分数、全氮质量分数、全磷质量分数、全钾质量分数、碱解氮质量分数、有效磷质量分数、速效钾质量分数、过氧化物酶活性、脲酶活性、酸性磷酸酶和β-葡萄糖苷酶活性。在运用主成分分析时,首先,根据特征值不小于 1 和累积方差贡献率不小于 80%,提取主成分。然后,在每个主成分中,选出不小于最大载荷值 90%的指标,对其进行 Pearson 相关分析,将相关系数之和最大的指标作为评价指标;若相关系数之和最大的指标不止1个,则选取载荷值较大的指标;若不小于最大载荷值 90%的指标只有1个,则该指标直接作为评价指标。
-
根据主成分分析得到所选取指标的公因子方差,计算各指标的权重(Wi):
$ W_{i}={C_{i}}\bigg{/}{\displaystyle \sum_{i=1}^{n} C_{i}} $ 。其中:n是综合评价的指标个数,Ci是第 i 个指标的公因子方差。 -
基于模糊数学模型,计算土壤性质的综合得分(S) :
$ S=\displaystyle \sum_{i=1}^{n} W_{i} F_{i} $ 。其中:Fi 是指标 i的离差标准化后的值。S越大,表示毛竹林皆伐后自然恢复的土壤综合恢复效果越好,反之则越差。 -
由表1可见:在0~10 cm表层土壤中,ck处理的土壤容重分别比CR、UR处理高31%和 14%;UR处理土壤容重略高于CR处理, 但无显著差异;UR、CR处理的土壤总孔隙度、毛管持水量、田间持水量和饱和持水量均高于ck处理,但无显著差异。在10~20 cm土壤中,ck处理土壤容重分别比CR、UR处理高39%和37%;ck、CR和UR处理的总孔隙度无显著差异;UR处理的毛管持水量、田间持水量和饱和持水量均高于CR和ck处理(P<0.05)。
表 1 不同处理的土壤物理性质
Table 1. Soil physical properties of different treatments
土层/cm 处理 容重/(g·cm−3) 总孔隙度/% 毛管持水量/(g·kg−1) 田间持水量/(g·kg−1) 饱和持水量/(g·kg−1) 0~10 CR 0.67±0.02 b 58.1±2.3 a 470±17 a 434±18 a 654±24 a UR 0.77±0.06 b 52.2±1.9 a 519±40 a 464±35 a 670±51 a ck 0.88±0.04 a 52.2±1.3 a 443±20 a 406±18 a 605±29 a 10~20 CR 0.67±0.02 b 61.4±1.7 a 487±15 b 447±14 b 682±24 b UR 0.68±0.05 b 58.3±1.8 a 596±47 a 527±39 a 767±59 a ck 0.93±0.03 a 54.0±1.3 a 439±22 b 394±19 b 610±28 b 说明:数据为平均值±标准误。不同小写字母表示同一土层不同处理间差异显著(P<0.05) -
由表2可见:在0~10 cm土壤中,UR处理的pH显著低于 CR和ck处理(P<0.05),UR处理的有机碳、全氮、全磷和碱解氮质量分数均显著高于CR和ck处理,UR处理的有机碳质量分数比CR和ck处理高99%和117%,全氮高93%和123%,全磷高57%和87%,碱解氮高95%和107%。ck处理的速效钾质量分数显著低于UR和CR(P<0.05)。在10~20 cm土壤中,UR处理的pH略低于CR和ck处理,UR处理的有机碳、全氮、全磷和碱解氮质量分数均显著高于CR和ck处理(P<0.05);UR处理的全钾质量分数显著低于CR和ck处理,ck处理的速效钾质量分数显著低于UR和CR处理 (P<0.05)。
表 2 不同处理的土壤化学性质
Table 2. Soil chemical properties of different treatments
土层/cm 处理 pH 有机碳/
(g·kg−1)全氮/
(g·kg−1)全磷/
(g·kg−1)全钾/
(g·kg−1)碱解氮/
(mg·kg−1)有效磷/
(mg·kg−1)速效钾/
(mg·kg−1)0~10 CR 5.09±0.06 a 36.86±2.62 b 3.27±0.24 b 0.37±0.02 b 25.29±1.61 a 101±7 b 1.4±0.1 b 189±13 a UR 4.73±0.06 b 73.25±7.25 a 6.30±0.54 a 0.58±0.04 a 16.58±0.43 b 96±11 a 1.9±0.1 a 195±15 a ck 5.03±0.09 a 33.73±2.78 b 2.82±0.23 b 0.31±0.01 b 22.67±1.10 a 95±5 b 2.1±0.1 a 105±13 b 10~20 CR 5.26±0.08 a 31.11±2.43 b 2.71±0.21 b 0.34±0.02 b 26.31±1.78 a 95±9 b 1.1±0.1 b 169±11 a UR 4.98±0.09 a 53.68±6.03 a 4.62±0.47 a 0.55±0.05 a 17.00±0.43 b 159±9 a 1.6±0.2 a 162±12 a ck 5.03±0.10 a 29.07±1.59 b 2.38±0.14 b 0.29±0.02 b 22.21±0.99 a 82±4 b 1.7±0.1 a 97±12 b 说明:数据为平均值±标准误。不同小写字母表示同一土层不同处理间差异显著(P<0.05) -
由表3可知:整体来看,除土壤酸性磷酸酶外,毛竹林皆伐后UR、CR处理的土壤脲酶、β-葡萄糖苷酶和过氧化物酶活性高于ck处理;UR处理土壤的4种胞外酶活性均高于CR处理,幅度为46%~98%。在0~10 cm土壤中,UR处理的脲酶、酸性磷酸酶和β-葡萄糖苷酶活性均显著高于CR和ck处理(P<0.05),3种处理的过氧化物酶活性无显著差异;ck处理土壤的酸性磷酸酶活性比CR处理高47%。在10~20 cm土壤中,UR处理的脲酶、酸性磷酸酶和β-葡萄糖苷酶活性均显著高于CR和ck处理(P<0.05),3种处理的过氧化物酶活性无显著差异。
表 3 不同处理的土壤胞外酶活性
Table 3. Soil extracellar enzyme activity of different treatments
土层/cm 处理 脲酶/
(μmol·g−1·h−1)酸性磷酸酶/
(μmol·g−1·h−1)β-葡萄糖苷酶/
(μmol·g−1·h−1)过氧化物酶/
(μmol·g−1·h−1)土层/cm 处理 脲酶/
(μmol·g−1·h−1)酸性磷酸酶/
(μmol·g−1·h−1)β-葡萄糖苷酶/
(μmol·g−1·h−1)过氧化物酶/
(μmol·g−1·h−1)0~10 CR 424±40 b 476±52 b 41.1±4.9 b 768±75 a 10~20 CR 338±44 b 508±59 b 37.7±4.3 b 766±132 a UR 836±101 a 944±90 a 77.0±11.6 a 1 123±152 a UR 542±75 a 884±87 a 71.9±8.0 a 1 039±184 a ck 290±71 b 701±76 b 44.0±8.0 b 770±147 a ck 243±40 b 569±66 b 37.3±7.1 b 723±104 a 说明:数据为平均值±标准误。不同小写字母表示同一土层不同处理间差异显著(P<0.05) -
通过表层土壤34个指标的主成分分析,提取7个主成分,累积贡献率为83.82%;第1主成分特征值为14.100,贡献率41.5%,主要影响因子为0~10 cm 全氮(xTNⅠ)、10~20 cm全氮(xTNⅡ)、0~10 cm 有机质(xSOCⅠ)、10~20 cm有机质(xSOCⅡ)、0~10 cm碱解氮(xANⅠ)、10~20 cm碱解氮(xANⅡ)等6个指标;第2主成分特征值为4.646,贡献率为13.7%,主要影响因子为0~10 cm孔隙度(xPⅠ)、10~20 cm孔隙度(xPⅡ)和10~20 cm饱和持水量(xShcⅡ)等3个指标;第3主成分特征值为3.100,贡献率为9.1%,主要影响因子为0~10 cm速效钾(xAKⅠ)和10~20 cm速效钾(xAKⅡ);第4主成分特征值为2.500,贡献率为7.3%,主要影响因子为0~10 cm有效磷(xAPⅠ)、10~20 cm有效磷(xAPⅡ)和10~20 cm β-葡萄糖苷酶活性(xBDⅡ);第5主成分特征值为1.610,贡献率为4.735%,主要影响因子为0~10 cm孔隙度(xPⅠ)和10~20 cm过氧化物酶活性(xPERⅡ);第6主成分特征值为1.300,贡献率为3.8%,主要影响因子为10~20 cm β-葡萄糖苷酶活性(xBDⅡ);第7主成分特征值为1.300,贡献率为3.7%,主要影响因子为10~20 cm酸性磷酸酶活性(xACPⅡ)。各主成分影响因子经相关分析后最终选取7个评价指标,分别为xTNⅠ、xPⅡ、xAKⅠ、xAPⅠ、xPERⅡ、xBGⅡ、xACPⅡ。
根据选定指标的离差标准化值和指标权重(表4)构建土壤性质综合评价模型:
表 4 综合评价指标及其权重
Table 4. Comprehensive evaluation indices and weights
指标 xTNⅠ xPⅡ xAKⅠ xAPⅠ xPERⅡ xBGⅡ xACPⅡ 指标权重 0.20 0.15 0.18 0.17 0.12 0.17 0.18 公因子方差 0.951 0.772 0.880 0.830 0.598 0.857 0.888 S=0.20F(xTNⅠ)+0.15F(xPⅡ)+0.18F(xAKⅠ)+0.17F(xAPⅠ)+0.12F(xPERⅡ)+0.17F(xBGⅡ)+0.18F (xACPⅡ)。
从0~10、10~20 cm等2层土壤综合评分结果来看(图1),毛竹林皆伐后恢复迹地得分均高于毛竹林得分,且UR处理得分高于CR处理。CR、UR、ck处理综合得分分别为0.41、0.61、0.37,UR处理综合得分分别比CR和ck处理高49%和 65%。UR处理的0~10 cm全氮得分为0.11,高于CR处理(0.05)和ck处理(0.04);CR处理和UR处理的0~10 cm速效钾得分均为0.10,高于ck处理(0.04);UR处理的10~20 cm β-葡萄糖苷酶得分为0.08,高于CR和ck处理(0.04);UR处理的10~20 cm酸性磷酸酶得分为0.10,高于CR (0.05)和ck处理(0.06)。
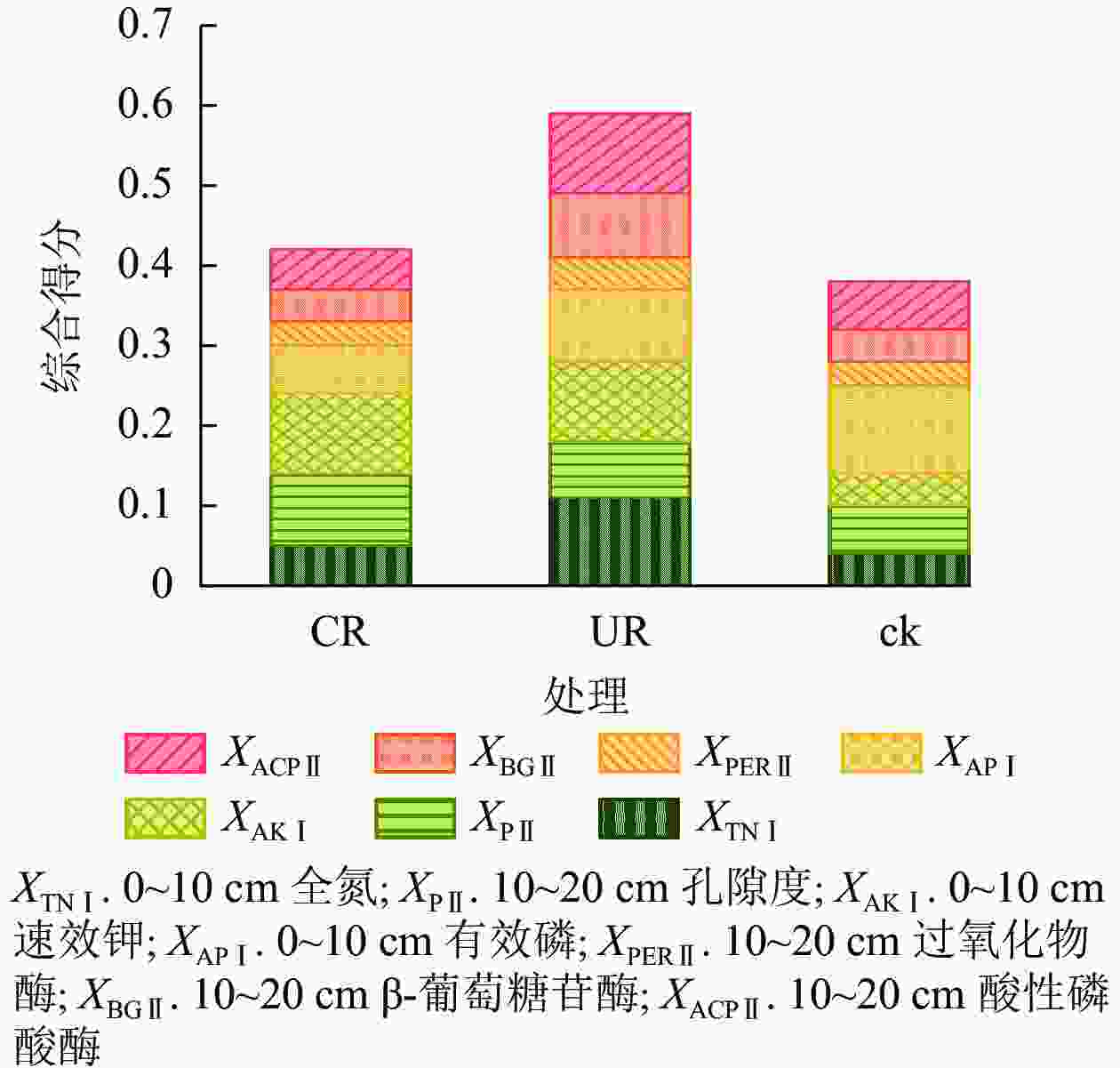
图 1 各样区的土壤性质综合得分
Figure 1. Synthesis scores of soil properties in the sample areas
-
自然条件下植物和土壤有着最直接的关联,具有密切的相互反馈作用,植物群落的变化会影响其生长环境中的土壤,最终改变土壤结构和土壤养分的有效性[14]。毛竹的入侵性极强,可通过其独特的竞争策略影响入侵林分其他物种的生长,改变入侵地的群落结构[11],导致植物物种多样性明显下降直至形成毛竹纯林,相应的土壤也会发生变化[15]。本研究结果表明:毛竹林皆伐后经5 a自然恢复,林地土壤与ck相比,土壤容重极显著降低,孔隙度和持水能力增加,土壤有机碳、全氮、全磷、碱解氮和速效钾质量分数提高,土壤胞外酶活性增强。主要原因是毛竹林皆伐后大量物种进入,植物多样性明显提高。这一结果与毛竹入侵相邻森林引起的植物多样性下降[16]、土壤退化是相反的,因此,通过毛竹林皆伐控制其快速扩张,可作为目前毛竹扩张蔓延区域的生态修复措施。
-
毛竹林采伐后会产生大量的采伐剩余物,处理方式不同会对迹地更新方式和生态修复产生影响。保留采伐剩余物覆盖在地表,腐烂分解后可增加土壤有机质和养分,减少土壤侵蚀,但会对植物种子萌发、幼苗生长和植被恢复产生一定的影响[17-18]。本研究表明:在0~10和10~20 cm土层土壤中,采伐剩余物保留样区的土壤有机碳、全氮、全磷、碱解氮及速效钾质量分数均显著高于采伐剩余物清理样区。这与池鑫晨等[15]、黄启堂等[19]、ZHAO等[20]、吴家森等[21]的研究结果一致,且采伐剩余物保留样区的脲酶、酸性磷酸酶、β-葡萄糖苷酶活性显著高于采伐剩余物清理样区。有研究表明:土壤酶活性与土壤有机质含量密切相关[22],土壤有机质含量高可以维持较高的土壤胞外酶活性[23],保留采伐剩余物后提高了土壤中的有机质。因此,建议在利用毛竹林皆伐手段进行区域生态修复时,应尽量保留采伐剩余物,但应让其分散均匀,不宜局部过厚堆积,减少对植被恢复影响。
-
毛竹林皆伐后自然恢复迹地土壤恢复良好,保留采伐剩余物的管理方式相较于清除采伐剩余物的管理方式可以更有效地提高土壤有机碳、全氮、全磷、碱解氮和速效钾质量分数,以及土壤胞外酶活性。在未来人工毛竹林弃管后实施生态修复时,保留采伐剩余物可以节省更多人力及资金成本,同时也可以达成更好的土壤恢复效果。
Effects of clear-cutting and harvest residue of Phyllostachys edulis forests on soil quality
-
摘要:
目的 毛竹Phyllostachys edulis林生态修复是当前中国亚热带地区面临的一个难题。了解毛竹林皆伐和剩余物保留后迹地土壤的自然恢复状况可为毛竹林生态修复提供指导。 方法 在毛竹林皆伐迹地设置了保留采伐剩余物(UR)、清理采伐剩余物(CR)和未采伐毛竹林地作为对照(ck)等3个处理。5 a后,通过土壤调查与测定,分析比较不同处理土壤指标变化,运用模糊判别和主成分分析,定量评价毛竹林皆伐后土壤自然恢复效果。 结果 ①CR、UR处理土壤容重分别比ck降低31%和14% (P<0.05),土壤总孔隙度、毛管持水量、田间持水量和饱和持水量均高于ck;UR处理土壤的持水力整体优于CR处理。②CR、UR处理土壤有机碳、全氮、全磷、碱解氮和速效钾质量分数均高于ck,各指标增加幅度为117%~123%;有效磷则表现为CR处理极显著(P<0.01)低于UR和ck;由于保留了毛竹林皆伐后采伐剩余物,UR处理土壤有机碳、全氮、全磷、碱解氮、有效磷显著高于CR处理33%~99% (P<0.05);③CR、UR处理土壤脲酶、β-葡萄糖苷酶和过氧化物酶活性高于ck;UR处理土壤3种胞外酶活性均高于CR处理46%~98%。④综合评价结果表明:土壤质量得到较好恢复,毛竹林皆伐后恢复迹地土壤综合得分从高到低依次为采伐剩余物保留样区、采伐剩余物清理样区、毛竹林样区。 结论 毛竹林皆伐后的土壤经过5 a自然恢复,与毛竹林林地土壤相比得到较快修复,毛竹林皆伐后保留采伐剩余物更有利于土壤修复。图1表4参23 Abstract:Objective Ecological restoration of Phyllostachys edulis forest is a challenge in subtropical regions of China. This study aims to understand the natural restoration status of the soil after clear-cutting and residue retention of Ph. edulis forest, so as to provide guidance for ecological restoration of forest. Method In the clear-cutting sites, 3 treatments were set up, i.e. cutting residue reserved (UR), cutting residue removed (CR), and uncut Ph. edulis forest as the control (ck). The changes of soil indexes under different treatments were analyzed and compared through soil survey and measurement 5 years later, and fuzzy mathematical discrimination and principal component analysis were used to quantitatively evaluate the natural restoration effect of Ph. edulis forest after clear-cutting. Result (1) The soil bulk density of CR and UR decreased by 31% and 14% respectively compared with ck (P<0.05). Soil total porosity, capillary water holding capacity, field water holding capacity and saturated water holding capacity were higher than those of ck. The water holding capacity of UR soil was better than that of CR. (2) The contents of soil organic carbon, total nitrogen, total phosphorus, alkali-hydrolyzed nitrogen and available potassium in CR and UR were higher than those in ck, and each index increased by 17%−123%. Available phosphorus showed that CR was significantly lower than UR and ck (P<0.01). Due to the retention of cutting residues of Ph. edulis forest after clear-cutting, the soil organic carbon, total nitrogen, total phosphorus, alkali-hydrolyzed nitrogen and available phosphorus in UR were significantly higher than those in CR treatment by 33%−99% (P<0.05). (3) The activities of urease, β-glucosidase and peroxidase in CR, UR soil were higher than those in ck. The activities of 3 extracellular enzymes in UR soil were 46%−98% higher than those in CR treatment. (4) The comprehensive evaluation results showed that the soil quality had been well restored, and the comprehensive scores ranging from high to low in the restored site soil of Ph. edulis forest after clear-cutting was sample area with cutting residue reserved, sample area with cutting residue removed, Ph. edulis forest sample area. Conclusion After 5 years of natural recovery, the soil of Ph. edulis forest after clear-cutting can be restored faster than that in Ph. edulis forest land, and the retention of the cutting residues after clear-cutting of Ph. edulis forest is more conducive to soil restoration. [Ch, 1 fig. 4 tab. 23 ref.] -
表 1 不同处理的土壤物理性质
Table 1. Soil physical properties of different treatments
土层/cm 处理 容重/(g·cm−3) 总孔隙度/% 毛管持水量/(g·kg−1) 田间持水量/(g·kg−1) 饱和持水量/(g·kg−1) 0~10 CR 0.67±0.02 b 58.1±2.3 a 470±17 a 434±18 a 654±24 a UR 0.77±0.06 b 52.2±1.9 a 519±40 a 464±35 a 670±51 a ck 0.88±0.04 a 52.2±1.3 a 443±20 a 406±18 a 605±29 a 10~20 CR 0.67±0.02 b 61.4±1.7 a 487±15 b 447±14 b 682±24 b UR 0.68±0.05 b 58.3±1.8 a 596±47 a 527±39 a 767±59 a ck 0.93±0.03 a 54.0±1.3 a 439±22 b 394±19 b 610±28 b 说明:数据为平均值±标准误。不同小写字母表示同一土层不同处理间差异显著(P<0.05)  下载: 导出CSV
下载: 导出CSV
表 2 不同处理的土壤化学性质
Table 2. Soil chemical properties of different treatments
土层/cm 处理 pH 有机碳/
(g·kg−1)全氮/
(g·kg−1)全磷/
(g·kg−1)全钾/
(g·kg−1)碱解氮/
(mg·kg−1)有效磷/
(mg·kg−1)速效钾/
(mg·kg−1)0~10 CR 5.09±0.06 a 36.86±2.62 b 3.27±0.24 b 0.37±0.02 b 25.29±1.61 a 101±7 b 1.4±0.1 b 189±13 a UR 4.73±0.06 b 73.25±7.25 a 6.30±0.54 a 0.58±0.04 a 16.58±0.43 b 96±11 a 1.9±0.1 a 195±15 a ck 5.03±0.09 a 33.73±2.78 b 2.82±0.23 b 0.31±0.01 b 22.67±1.10 a 95±5 b 2.1±0.1 a 105±13 b 10~20 CR 5.26±0.08 a 31.11±2.43 b 2.71±0.21 b 0.34±0.02 b 26.31±1.78 a 95±9 b 1.1±0.1 b 169±11 a UR 4.98±0.09 a 53.68±6.03 a 4.62±0.47 a 0.55±0.05 a 17.00±0.43 b 159±9 a 1.6±0.2 a 162±12 a ck 5.03±0.10 a 29.07±1.59 b 2.38±0.14 b 0.29±0.02 b 22.21±0.99 a 82±4 b 1.7±0.1 a 97±12 b 说明:数据为平均值±标准误。不同小写字母表示同一土层不同处理间差异显著(P<0.05)
下载: 导出CSV
表 3 不同处理的土壤胞外酶活性
Table 3. Soil extracellar enzyme activity of different treatments
土层/cm 处理 脲酶/
(μmol·g−1·h−1)酸性磷酸酶/
(μmol·g−1·h−1)β-葡萄糖苷酶/
(μmol·g−1·h−1)过氧化物酶/
(μmol·g−1·h−1)土层/cm 处理 脲酶/
(μmol·g−1·h−1)酸性磷酸酶/
(μmol·g−1·h−1)β-葡萄糖苷酶/
(μmol·g−1·h−1)过氧化物酶/
(μmol·g−1·h−1)0~10 CR 424±40 b 476±52 b 41.1±4.9 b 768±75 a 10~20 CR 338±44 b 508±59 b 37.7±4.3 b 766±132 a UR 836±101 a 944±90 a 77.0±11.6 a 1 123±152 a UR 542±75 a 884±87 a 71.9±8.0 a 1 039±184 a ck 290±71 b 701±76 b 44.0±8.0 b 770±147 a ck 243±40 b 569±66 b 37.3±7.1 b 723±104 a 说明:数据为平均值±标准误。不同小写字母表示同一土层不同处理间差异显著(P<0.05)
下载: 导出CSV
表 4 综合评价指标及其权重
Table 4. Comprehensive evaluation indices and weights
指标 xTNⅠ xPⅡ xAKⅠ xAPⅠ xPERⅡ xBGⅡ xACPⅡ 指标权重 0.20 0.15 0.18 0.17 0.12 0.17 0.18 公因子方差 0.951 0.772 0.880 0.830 0.598 0.857 0.888
下载: 导出CSV
-
[1] NAGATI M, ROY M, MANZI S, et al. Impact of local forest composition on soil fungal communities in a mixed boreal forest [J]. Plant Soil, 2018, 432(1/2): 345 − 357. [2] CRAWFORD K M, BAUER J T, COMITA S, et al. When and where plant-soil feedback may promote plant coexistence: a meta-analysis [J]. Ecol Lett, 2019, 22(8): 1274 − 1284. [3] QIAO Leilei, LI Yuanze, SONG Yahui, et al. Effects of vegetation restoration on the distribution of nutrients, glomalin-related soil protein, and enzyme activity in soil aggregates on the loess plateau, China[J/OL]. Forests, 2019, 10(9): 796[2021-12-10]. doi: 10.3390/10090796. [4] 蓝斌, 何东进. 闽北毛竹林生态系统能量分配规律的研究[J]. 应用生态学报, 2000, 11(2): 193 − 195. LAN Bin, HE Dongjin. Energy distribution of Phyllostachys pubescens ecosytem in north Fujian [J]. Chin J Appl Ecol, 2000, 11(2): 193 − 195. [5] OKUTOMI K, SHINODA S, FUKUDA H. Causal analysis of the invasion of broad-leaved forest by bamboo in Japan [J]. J Veg Sci, 1996, 7(5): 723 − 728. [6] ISAGI Y, TORII A. Range Expansion and its mechanisms in a naturalized bamboo species, Phyllostachys pubescens, in Japan [J]. J Sustainable For, 1997, 6(1/2): 127 − 141. [7] 丁丽霞, 王祖良, 周国模, 等. 天目山国家级自然保护区毛竹林扩张遥感监测[J]. 浙江林学院学报, 2006, 23(3): 297 − 300. DING Lixia, WANG Zuliang, ZHOU Guomo, et al. Monitoring Phyllostachys pubescens stands expansion in National Nature Reserve of Mount Tianmu by remote sensing [J]. J Zhejiang For Coll, 2006, 23(3): 297 − 300. [8] OSEI R, ANSONG M, ZERBE S. Comparison of socio-economic and ecological benefits of bamboo and trees: the perspectives of local communities in south-western Ghana [J]. Southern For J For Sci, 2019, 81(3): 255 − 260. [9] 白尚斌, 周国模, 王懿祥, 等. 天目山国家级自然保护区毛竹扩散过程的林分结构变化研究[J]. 西部林业科学, 2012, 41(1): 77 − 82. BAI Shangbin, ZHOU Guomo, WANG Yixiang, et al. Stand structure change of Phyllostachys pubescens forest expansion in Tianmushan National Nature Reserve [J]. J West China For Sci, 2012, 41(1): 77 − 82. [10] 王铮屹, 戴其林, 柏宬, 等. 天目山皆伐毛竹林自然更新群落类型与多样性分析[J]. 浙江农林大学学报, 2020, 37(4): 710 − 719. WANG Zhengyi, DAI Qilin, BAI Cheng, et al. Types and diversity of natural regeneration community after clear cutting of Phyllostachys edulis forests in Mount Tianmu, China [J]. J Zhejiang A&F Univ, 2020, 37(4): 710 − 719. [11] 陈旭, 刘宗悦, 徐钧杰, 等. 天目山毛竹林皆伐后群落的恢复特征[J]. 浙江农林大学学报, 2022, 39(4): 705 − 716. CHEN Xu, LIU Zongyue, XU Junjie, et al. Restoration characteristics of Phyllostachys edulis community after clear-cutting in Mount Tianmu [J]. J Zhejiang A&F Univ, 2022, 39(4): 705 − 716. [12] 鲁如坤. 土壤农业化学分析方法[M]. 北京: 中国农业科技出版社, 2000. LU Rukun. The Analysis Method of Soil Agricultural Chemistry[M]. Beijing: China Agricultural Science and Technology Press, 2000. [13] GERMAN D P, WEINTRAUB M N, GRANDY A S, et al. Optimization of hydrolytic and oxidative enzyme methods for ecosystem studies [J]. Soil Biol Biochem, 2011, 43(7): 1387 − 97. [14] FUJII K, SHIBATA M, KITAJIMA K, et al. Plant-soil interactions maintain biodiversity and functions of tropical forest ecosystems [J]. Curr Top Ecol, 2018, 33(1): 149 − 160. [15] 池鑫晨, 宋超, 朱向涛, 等. 毛竹入侵常绿阔叶林对土壤活性有机碳氮的动态影响[J]. 生态学杂志, 2020, 39(7): 2263 − 2272. CHI Xinchen, SONG Chao, ZHU Xiangtao, et al. Effects of moso bamboo invasion on soil active organic carbon and nitrogen in a evergreen broad-leaved forest in subtropical China [J]. Chin J Ecol, 2020, 39(7): 2263 − 2272. [16] 白尚斌, 周国模, 王懿祥, 等. 天目山保护区森林群落植物多样性对毛竹入侵的响应及动态变化[J]. 生物多样性, 2013, 21(3): 288 − 295. BAI Shangbin, ZHOU Guomo, WANG Yixiang, et al. Plant species diversity and dynamics in forests invaded by moso bamboo (Phyllostachys edulis) in Tianmu Mountain Nature Reserve [J]. Biodiversity Sci, 2013, 21(3): 288 − 295. [17] EISENBIES M H, VANCE E D, AUST W M, et al. Intensive utilization of harvest residues in southern pine plantations: quantities available and implications for nutrient budgets and sustainable site productivity [J]. Bioenergy Res, 2009, 2(3): 90 − 98. [18] 吴传敬, 郭剑芬, 许恩兰, 等. 采伐残余物不同处理方式对杉木幼林土壤有机碳组分和相关酶活性的影响[J]. 土壤学报, 2019, 56(6): 1504 − 1513. WU Chuanjin, GUO Jianfeng, XU Enlan, et al. Effects of logging residue on composition of soil carbon and activity of related enzymes in soil of a young Chinese fir plantation as affected by residue handling mode [J]. Acta Pedol Sin, 2019, 56(6): 1504 − 1513. [19] 黄启堂, 陈爱玲, 贺军. 不同毛竹林林地土壤理化性质特征比较[J]. 福建林学院报, 2006, 26(4): 299 − 302. HUANG Qitang, CHEN Ailing, HE Jun. Comparison of soil physical and chemical properties among various Phyllostachys pubescens plantation [J]. J Fujian Coll For, 2006, 26(4): 299 − 302. [20] ZHAO Yingzhi, LIANG Chenfei, SHAO Shuai, et al. Linkages of litter and soil C∶N∶P stoichiometry with soil microbial resource limitation and community structure in a subtropical broadleaf forest invaded by moso bamboo [J]. Plant Soil, 2021, 465(1/2): 473 − 490. [21] 吴家森, 姜培坤, 王祖良. 天目山国家级自然保护区毛竹扩张对林地土壤肥力的影响[J]. 江西农业大学学报, 2008, 30(4): 689 − 692. WU Jiasen, JIANG Peikun, WANG Zuliang. The effects of Phyllostachys pubescens expansionon onsoil fertility in National Nature Reserve of Mount Tianmu [J]. Acta Agric Univ Jiangxi, 2008, 30(4): 689 − 692. [22] GRANDY A S, STRICKLAND M S, LAUBER C L, et al. The influence of microbial communities, management, and soil texture on soil organic matter chemistry [J]. Geoderma, 2009, 150(3/4): 278 − 286. [23] CHODAK M, NIKLIŃSKA M. The effect of different tree species on the chemical and microbial properties of reclaimed mine soils [J]. Biol Fert Soil, 2010, 46(6): 555 − 566. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20220122
 点击查看大图
点击查看大图
计量
- 文章访问数: 1523
- HTML全文浏览量: 447
- PDF下载量: 97
- 被引次数: 0


