







-
掌叶覆盆子Rubus chingii,又称华东覆盆子、葛公等,为蔷薇科Rosaceae悬钩子属Rubus植物,药食同源,广泛分布于浙江、安徽、江西等省份的山地丘陵中,为中国特有的红树莓[1−2]。未成熟果实入药,具有益肾、固精和缩尿等功效,药用历史悠久[3−4]。成分分析和药理研究表明:掌叶覆盆子富含多糖、酚酸和萜类等物质,具消炎、抗氧化、抗癌、抗糖尿病、降血脂等多重作用[5−8]。成熟果实酸甜可口,营养丰富,富含酚酸、氨基酸、维生素PP、锰、锌等,深受消费者喜爱[1, 9]。但是,覆盆子果实小、鲜果极不耐储藏和运输,限制了其产业的发展。阐明影响覆盆子果实膨大和成熟软化的关键基因,对于推动药果和鲜果产业发展具有重要的意义。
植物细胞壁由纤维素、半纤维素和果胶等多糖组成。多聚半乳糖醛酸酶(polygalacturonase,PG)是果胶水解酶的重要成员之一,含多个基因家族成员,与果胶甲酯酶(pectin methylesterase,PME)、β-半乳糖苷酶(β-galactosidase, β-GAL)和鼠李糖半乳糖醛酸酶(rhamnogalacturonase,RGase)等共同参与植物细胞壁果胶的水解,介导植物的种子萌发、侧根发育、器官脱落、逆境响应等发育和生理过程,在果实成熟与软化过程中发挥关键作用,已在拟南芥Arabidopsis thaliana、番茄Solanum lycopersicum、南瓜Cucumis sativus、西瓜Citrullus lanatus、玉米Zea mays和甘薯Ipomoea batatas等植物中被广泛关注[10−14]。
PG属于糖基水解酶28号家族(GH28),该蛋白含有至少1个结构域GH28 (Pfam00295)。研究发现:PG基因家族成员均有其特定的氨基酸结构,包括4个典型的保守功能结构域:Ⅰ(SPNTDG)、Ⅱ(GDDC)、Ⅲ (CGPGHGISIGSLG)和Ⅳ (RIK),其中,结构域Ⅲ的保守性较差,且在鼠李糖多聚半乳糖醛酸酶中缺失[15]。这4个保守结构域都具有特定的功能。结构域Ⅰ和Ⅱ中,NTD和DD中的天冬氨酸(D)残基是催化位点的组成部分,结构域Ⅲ中,组氨酸(H)残基起催化作用,而结构域Ⅳ可能与果胶底物的羧酸基团相互作用[16]。有学者在氨基酸水平上对PG基因家族进行分类,提出了2种分类系统。第一种由KIM等[17]提出,对来自拟南芥和水稻Oryza sativa的125个PGs进行系统发育分析,结果可分为3类(Class A~C),每个类别可再细分出不同亚类; MAHMOOD等[18]根据此方法对来自54种植物的
2786 个PGs做出分类。另一种由PARK等[15]提出,对来自水稻和一种丝状真菌Aspergillus oryzae的225个PGs进行系统发育和基因结构分析,结果分为6个分支(Clade A~F),同一分支的差异较小,内含子数量接近。之后,LIANG等[16]将拟南芥的QRT3及其在双子叶植物和单子叶中的14个同源蛋白归类为新分支(Clade G),从而将557个PGs分为7个分支,即Clade A~G。尽管PG基因在许多植物种中已被鉴定出来,但是在掌叶覆盆子中的成员与功能仍不清楚。本研究基于掌叶覆盆子基因组,对其PG基因家族进行了全基因组鉴定和分析,并结合转录组数据与实时荧光定量PCR (RT-qPCR),分析了果实不同发育阶段家族成员的表达模式,进而筛选出调控果实成熟软化的关键PG基因,以期为掌叶覆盆子PG基因功能的解析及分子育种和果实保鲜提供科学依据。
-
掌叶覆盆子L7株系的果实,按其果实发育过程分为8个阶段(图1):小绿(SG,花后7 d),中绿(MG,花后14 d),大绿Ⅰ(BG Ⅰ,花后21 d),大绿Ⅱ(BG Ⅱ,花后28 d),大绿Ⅲ(BG Ⅲ,花后35 d),青转黄(GY,花后42 d),黄转橙(YO,花后48 d)和红色(RE,花后54 d)[6]。分别采集各阶段果实液氮速冻后超低温保存。
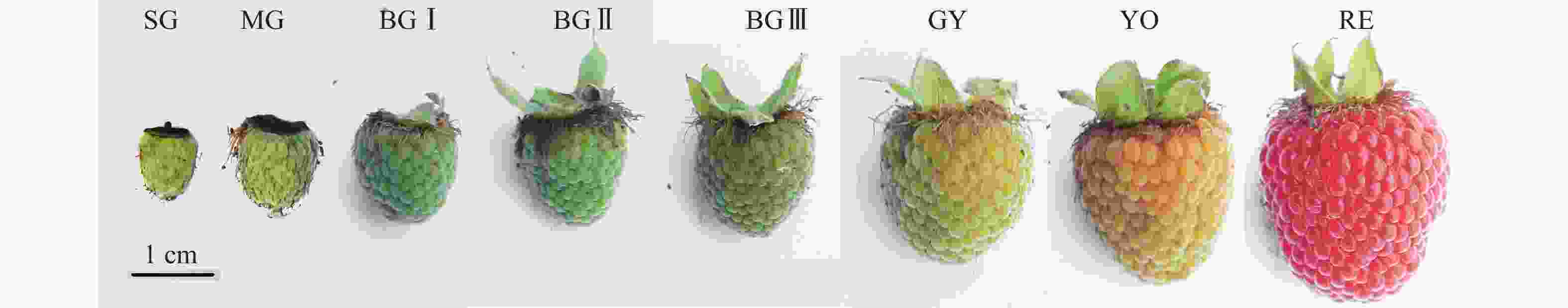
图 1 掌叶覆盆子不同发育阶段的果实
Figure 1. Fruits of R. chingii at different developmental stages
-
从蔷薇科植物数据库GDR网站(
https://www.rosaceae.org )、TAIR数据库和美国国家生物技术信息中心(NCBI) (https://www.ncbi.nlm.nih.gov )中下载掌叶覆盆子、拟南芥和番茄的全基因组序列与蛋白质序列[12, 19]。以糖基水解酶28号家族(PF00295)结构域的隐马尔可夫模型(HMM)作为规范结构域,使用HMMER 3.3.1 (http://hmmer.org/ download.html )进行HMM搜索序列同源物(E≤10−5),并利用Conserved Domain Database (https://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi )和每千个碱基/百万个定位片段(PFAM) 确认,以确保含有PG保守结构域。 -
将掌叶覆盆子、拟南芥和番茄PG家族成员氨基酸序列进行多序列比对,用MEGA 11软件构建进化树,采用邻近法(NJ),检验参数bootstrap设置为1 000。根据拟南芥分组的命名,结合染色体位置,对掌叶覆盆子PG基因进行重命名和分组。最后,通过Chiplot在线工具(https://www.chiplot.on line/)对构建的系统发育树进行注释和修饰。
-
根据掌叶覆盆子基因组注释数据,使用TBtools软件的基因定位可视化,绘制RchPGs基因在染色体上的位置,并绘制外显子-内含子的基因结构。使用MEME在线网站识别之前鉴定的RchPGs基因的保守基序,并在程序中将最大碱基数设置为10,预测结果通过TBtools展示[20]。掌叶覆盆子基因组内及其基因组与番茄基因组的共线性分析采用MCScanX,并用TBtools的advanced circos和dual synteny plot可视化。
-
选择典型的4个阶段果实(BG Ⅰ、GY、YO和RE),每组3个生物学重复,送至华大基因有限公司测序[6]。分析掌叶覆盆子PGs基因在果实发育阶段的表达模式。
-
使用全能型植物RNA提取试剂盒(CW25983)提取各阶段果实的总RNA,琼脂糖凝胶电泳检测3条条带,测定浓度,使用逆转录试剂盒(RR036A)逆转录成cDNA。RT-qPCR使用实时荧光定量PCR仪(CFX 96)。生物学重复4次。数据采用单因素方差分析法(LSD)。
-
通过HMM分析掌叶覆盆子的基因组数据,共鉴定出47个PGs基因(表1)。根据染色体位置分布,将47个RchPGs基因命名为RchPG1~RchPG47。氨基酸序列分析表明:RchPGs蛋白分子量为15.81~131.12 kDa,平均分子量为39.51 kDa。RchPGs蛋白等电点为5.00~9.49,说明RchPGs蛋白的酸碱性差异较大,其中47%为酸性,53%为碱性。亲疏水指数(GRAVY)为−0.585~0.056,其中91%的成员为亲水蛋白(GRAVY<0)。约34%的RchPGs不稳定,其余成员较稳定。亚细胞预测表明:RchPGs蛋白均位于细胞壁上。对掌叶覆盆子PG基因编码的氨基酸进行多序列对比,47个RchPGs中,有28个都具有4个典型的保守结构域Ⅰ~Ⅳ;11个RchPGs不包含结构域Ⅲ,除RchPG41外,均属于Clade E (图2)。此外,还有一部分RchPGs蛋白保守基序中个别氨基酸发生变异,如SPNTDG中的“S”变成“A”,CGPGHG中的第2个“G”变成“D”等。
表 1 掌叶覆盆子RchPGs家族成员特征
Table 1. Characteristics of RchPGs family members in R. chingii
基因编码 基因名称 等电点 分子量/kDa 亲疏水
指数不稳定
系数分支 基因编码 基因名称 等电点 分子量/kDa 亲疏水
指数不稳定
系数分支 LG01.1059 RchPG1 5.96 51.08 −0.073 44.79 E LG04.283 RchPG25 8.63 57.49 −0.356 45.27 A LG01.1356 RchPG2 6.83 51.99 −0.127 44.12 E LG04.396 RchPG26 5.26 49.55 −0.183 33.24 B LG01.1565 RchPG3 8.11 42.23 0.056 29.10 C LG04.993 RchPG27 9.02 52.04 −0.278 44.66 A LG01.1665 RchPG4 6.43 39.40 −0.063 24.24 C LG05.1140 RchPG28 8.48 44.96 −0.174 42.43 E LG01.2532 RchPG5 6.08 47.67 −0.267 38.63 D LG05.1261 RchPG29 5.19 52.20 −0.018 38.83 E LG01.2853 RchPG6 9.09 55.47 −0.193 43.86 E LG06.1099 RchPG30 8.90 50.20 −0.242 28.08 D LG01.3312 RchPG7 8.23 58.83 −0.239 46.37 F LG06.1103 RchPG31 6.96 43.60 −0.148 23.68 D LG01.3577 RchPG8 8.52 15.81 0.008 38.87 C LG06.1105 RchPG32 8.98 42.48 −0.210 27.47 D LG01.3578 RchPG9 9.49 41.00 0.035 39.66 C LG06.1675 RchPG33 7.81 43.81 −0.197 35.69 F LG01.3579 RchPG10 9.48 107.01 −0.032 41.27 C LG06.2235 RchPG34 8.81 49.23 −0.059 43.94 B LG02.1098 RchPG11 8.01 40.06 −0.059 30.89 F LG06.272 RchPG35 7.48 53.19 −0.275 36.87 A LG02.1344 RchPG12 5.39 49.49 −0.063 43.95 E LG06.3272 RchPG36 5.50 51.64 −0.186 36.12 B LG02.1359 RchPG13 5.76 50.91 −0.192 39.89 D LG06.3274 RchPG37 5.00 45.35 −0.167 34.66 F LG02.1479 RchPG14 6.24 131.12 −0.585 51.01 D LG06.3368 RchPG38 5.65 52.96 −0.054 44.42 E LG02.1829 RchPG15 9.35 49.19 −0.451 34.18 B LG06.4848 RchPG39 7.16 48.48 −0.108 48.34 A LG02.1835 RchPG16 8.94 49.80 −0.288 30.39 B LG07.1450 RchPG40 8.95 48.86 −0.283 34.24 B LG02.2252 RchPG17 7.48 49.52 −0.254 38.72 D LG07.1820 RchPG41 7.94 40.24 −0.292 32.80 B LG02.4536 RchPG18 6.38 44.22 −0.015 31.62 D LG07.1821 RchPG42 6.30 48.45 −0.227 25.48 B LG03.1665 RchPG19 8.94 44.71 −0.181 27.06 D LG07.1856 RchPG43 6.44 48.39 −0.256 29.74 B LG03.5002 RchPG20 5.95 52.65 −0.228 40.49 D LG07.1859 RchPG44 7.80 48.12 −0.263 23.46 B LG04.1300 RchPG21 6.39 40.22 −0.357 27.43 F LG07.2143 RchPG45 5.19 45.73 −0.275 37.35 F LG04.1461 RchPG22 6.14 74.72 −0.390 57.75 A LG07.325 RchPG46 7.54 51.44 −0.076 37.22 E LG04.1621 RchPG23 6.14 46.91 0.094 48.01 E LG07.3923 RchPG47 6.09 49.37 −0.170 34.22 E LG04.1640 RchPG24 8.74 46.54 −0.105 33.44 D 
图 2 RchPGs蛋白的保守结构域
Figure 2. Conserved domains of RchPGs proteins
-
将47个RchPGs蛋白和68个拟南芥AtPGs以及54个番茄SlPGs蛋白序列,使用邻接法构建系统进化树(图3)。这169个PGs蛋白被分为7个亚家族,分别是Clade A~G。其中Clade D包含11个RchPGs家族成员,Clade B和Clade E均包含10个RchPGs家族成员,Clade F包含6个RchPGs,Clade A和Clade C均包含5个RchPGs家族成员。在拟南芥和番茄中还有Clade G分支,分别包含At4G20050和SlPG71 (Solyc02g068400.2)。RchPG16属于Clade B分支,与RchPG36、RchPG43、RchPG42、RchPG41和RchPG44亲缘关系最近。

图 3 掌叶覆盆子与拟南芥、番茄PG家族成员的系统发育树
Figure 3. Phylogenetic tree of PG family members from R. chingii, A. thaliana, and S. lycopersicum
-
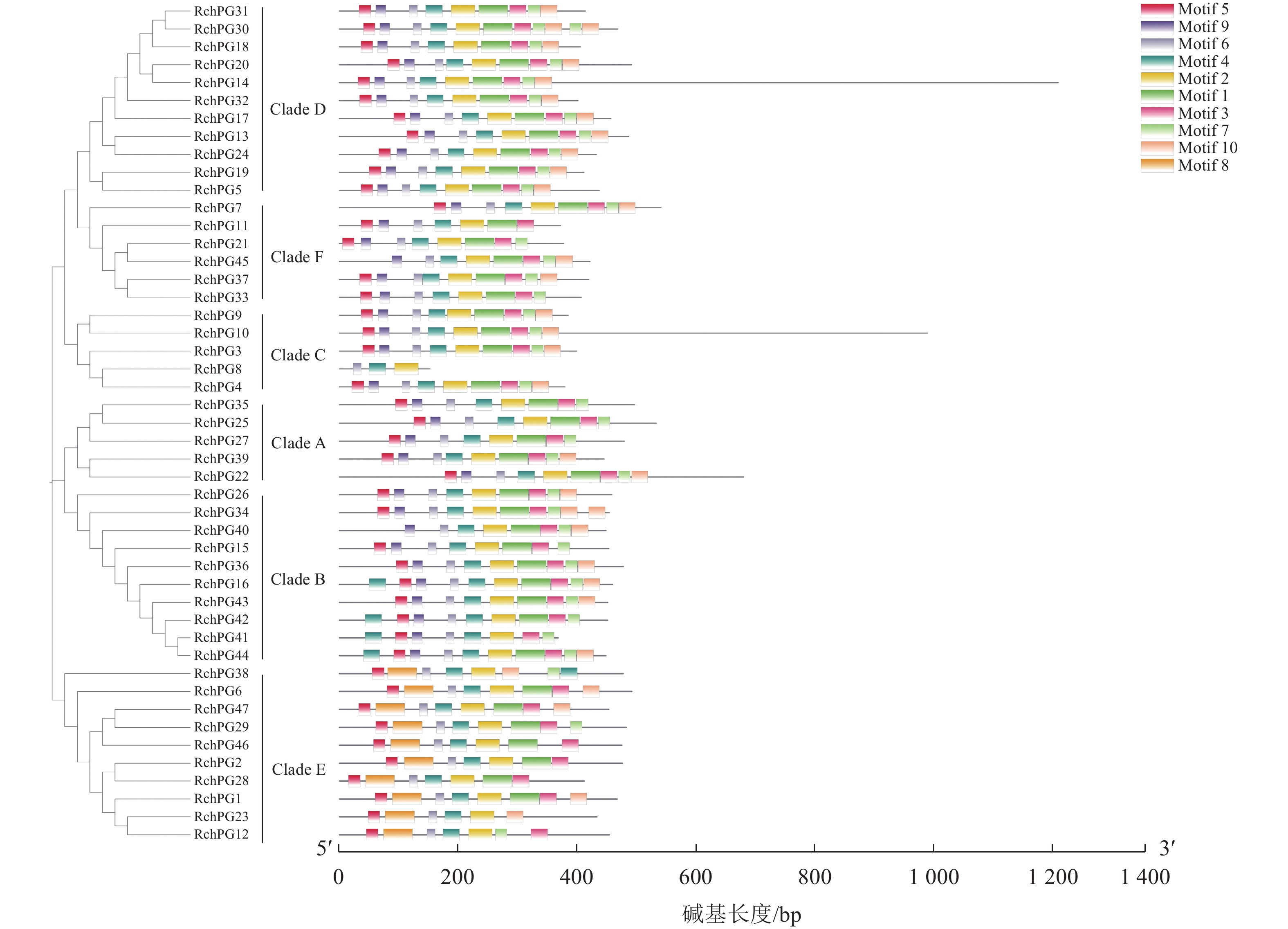
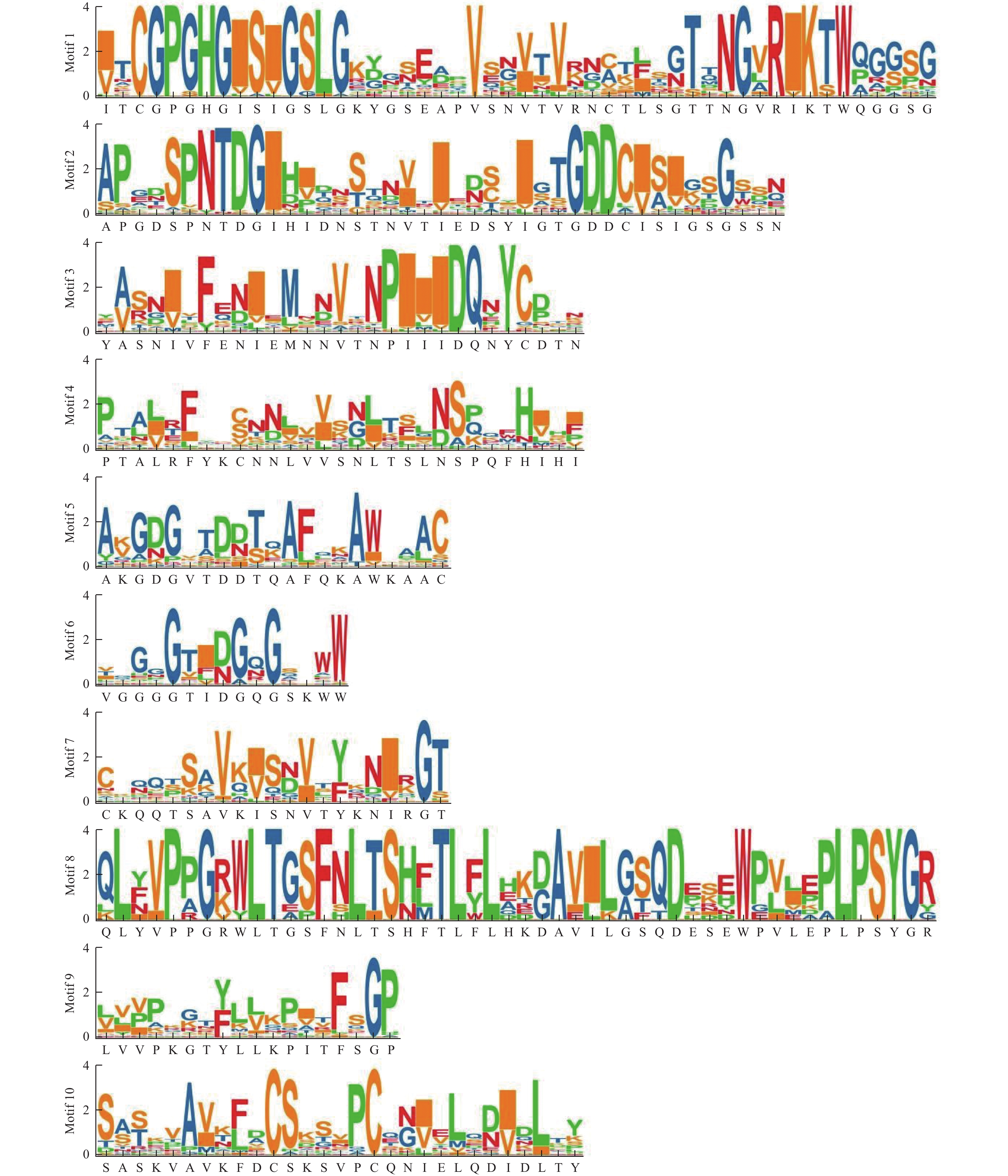
RchPGs基因结构显示:外显子为2~13个,内含子为1~12个(图4)。Clade E分支的RchPGs除了RchPG38含3个外显子外,其余均含有5~6个外显子;Clade B分支中大部分RchPGs含有7~9个外显子;Clade D分支大部分RchPGs含有4~7个外显子。不同分支的外显子和内含子结构不同,但同一分支外显子和内含子的位置和长度相对保守。使用MEME网站鉴定和分析47个RchPGs蛋白的保守基序,命名为Motif 1~10 (图5~6)。47条RchPGs蛋白均存在Motif 6、Motif 4和Motif 2;其中仅RchPG8、RchPG38、RchPG41、RchPG23和RchPG12不含Motif 1。Clade E成员均不含有Motif 9,且Motif 8只存在于Clade E中。Clade A中的RchPG39和RchPG22、Clade C中除RchPG8的其他成员和Clade D中的所有成员都含有除了Motif 8以外的9个基序。但是RchPG8仅含有Motif 6、Motif 4和Motif 2。此外,这些保守基序在PG蛋白序列中的组成和位置顺序在进化分支中大体相似,这表明了用于亚科分类的系统发育分析的可靠性。

图 4 掌叶覆盆子RchPGs基因家族结构分析以及系统发育树
Figure 4. Gene structures analyses of RchPGs families and phylogenetic tree of RchPGs in R. chingii

图 5 掌叶覆盆子RchPGs 蛋白家族保守基序预测
Figure 5. Motif prediction of RchPGs family members in R. chingii
-
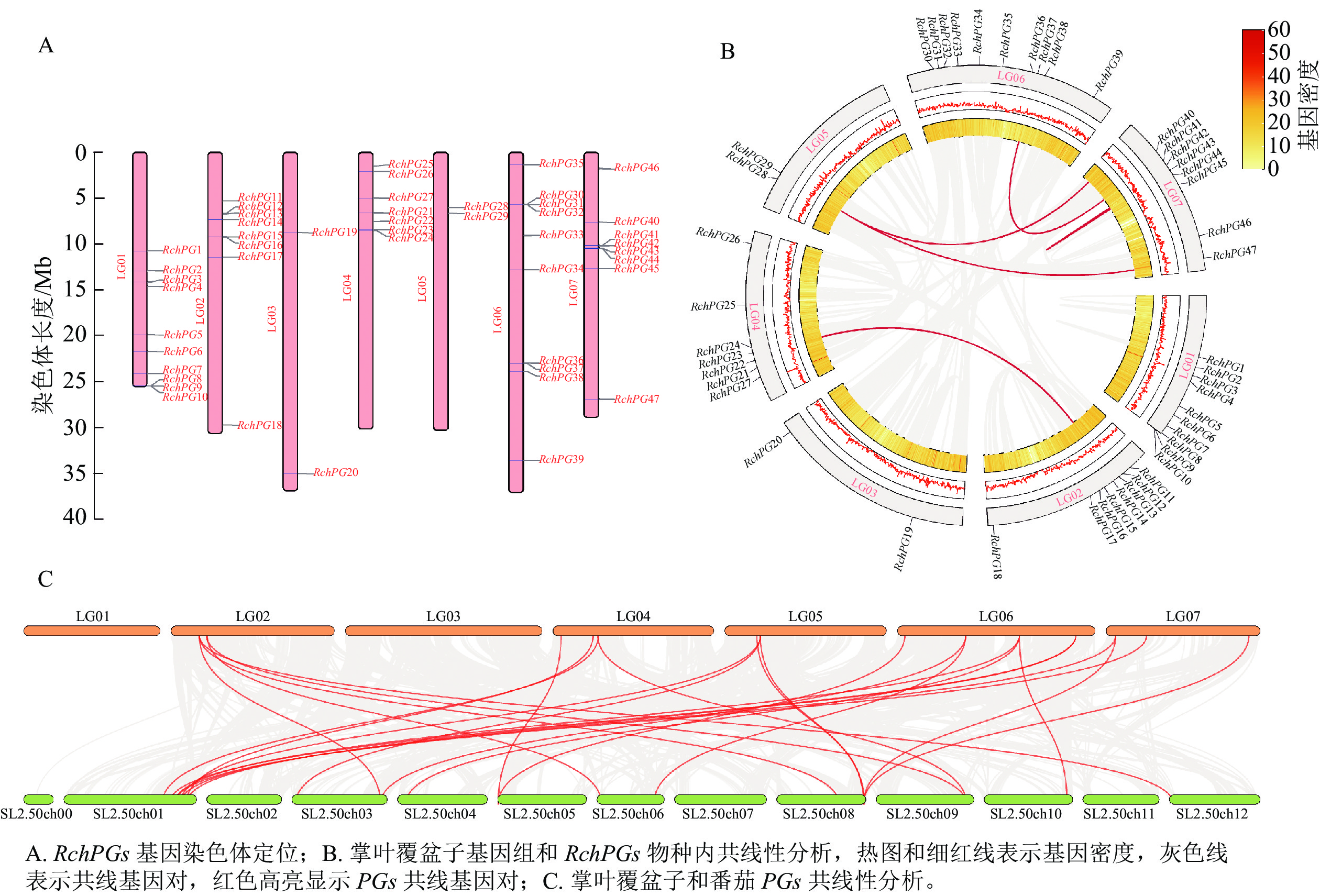
RchPGs基因在掌叶覆盆子的LG01~LG07染色体上均存在,并且分布不均匀(图7A)。LG01和LG06上RchPGs基因分布最多,均有10条;其次是LG02和LG07,分布有8条RchPGs基因;接着是LG04,含有7条RchPGs基因。LG03和LG05上分布最少,分别仅有2条RchPGs基因。在这些PGs基因中,发生了4次串联复制事件,分别是RchPG8~RchPG10、RchPG15和RchPG16、RchPG30~RchPG32以及RchPG36和RchPG37。共线性分析表明:掌叶覆盆子中存在5个PG编码基因对,包括RchPG12和RchPG23、RchPG13和RchPG24、RchPG29和RchPG47、RchPG36和RchPG40以及RchPG41和RchPG43 (图7B)。此外,番茄和掌叶覆盆子PGs基因间共有25个共线对:RchPG11分别和SlPG64、SlPG68、SlPG70存在共线性,RchPG12和RchPG23均和SlPG48-2存在共线性,RchPG28和SlPG49存在共线性,RchPG29分别和SlPG55-2、SlPG56-1、SlPG56-3存在共线性,RchPG36分别和SlPG16、SlPG15、SlPG14存在共线性等(图7C)。

图 6 掌叶覆盆子RchPGs 蛋白家族保守基序序列分析
Figure 6. Motif sequence analysis of RchPGs family members in R. chingii

图 7 RchPGs基因染色体定位和共线性分析
Figure 7. Chromosomal location and synteny analyses of RchPGs in R. chingii
-
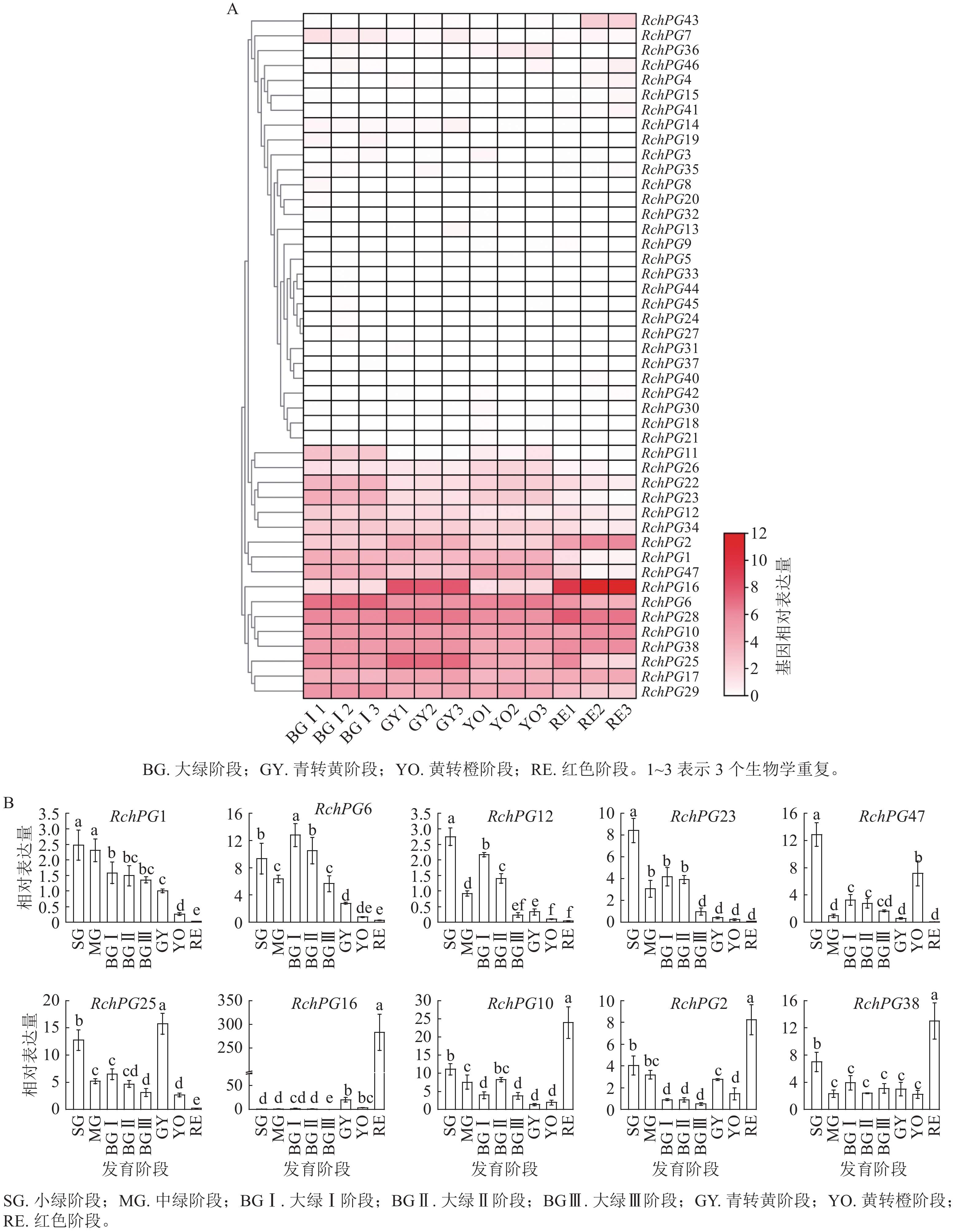
图8A结果表明:46条RchPGs基因家族成员均可在果实中检测到,其中RchPG1、RchPG2、RchPG6、RchPG10、RchPG16、RchPG17、RchPG22、RchPG23、RchPG25、RchPG28、RchPG38和RchPG47等在果实发育过程中表达量较高;而RchPG3~RchPG5、RchPG7~RchPG9、RchPG13~RchPG15、RchPG18~RchPG21、RchPG24和RchPG27等在果实中表达量很低,推测它们可能在根、茎、叶等其他器官发育过程中起作用。其中,RchPG6、RchPG1、RchPG12、RchPG23和RchPG29在果实发育早期起作用,介导果实膨大过程中的细胞壁水解,它们均属于Clade E分支。RchPG47 (Clade E)和RchPG25 (Clade A)在果实早期表达也较高,且RchPG25在GY阶段表达明显升高,而RchPG47在GY阶段表达量未明显升高,YO阶段表达量略微升高,此后又急剧下降。RchPG2、RchPG28、RchPG38、RchPG10和RchPG16等基因在果实发育后期表达量升高,介导果实成熟与软化。其中RchPG16基因表达量差异最大,BG Ⅰ阶段的表达量水平为1.78 (FPKM),GY阶段升高至236.54,YO阶段下降,最后在RE阶段急剧升高至
3859.35 ,是未成熟BG Ⅰ时期的2168.17 倍。
图 8 掌叶覆盆子RchPGs基因表达量转录组分析与RT-qPCR鉴定
Figure 8. Transcriptome analysis and RT-qPCR identification of RchPGs genes expression in R.chingii
选取8个发育阶段的果实,提取总RNA,逆转成cDNA后,RT-qPCR检测RchPG1、RchPG6、RchPG12、RchPG23、RchPG25、RchPG47、RchPG2、RchPG10、RchPG38和RchPG16等基因的相对表达量(以GY时期RchPG1的表达量为参照值1),结果表明各基因的表达量与转录组测序结果基本一致。RchPG16基因有两次表达量急剧增加,第1次是从BG Ⅲ到GY时期表达量增加了约285.71倍,第2次是从YO进入RE时期表达量增加了83.26倍,RE时期的表达量是BG Ⅰ时期的283.08倍(图8B)。
-
PG属于糖基水解酶28号家族(GH28),在1988年首次被发现。已在不少高等植物中鉴定了PG基因,其中在拟南芥、水稻、杨树Populus、黄瓜、西瓜、甘薯中分别鉴定到68、46、75、53、62和103个PGs基因[14]。本研究在覆盆子中鉴定到47个RchPGs基因,不均匀分布在7条染色体上,系统发育可分为6个分支(Clade A~F)。PG基因家族从最初分为3个进化枝(Clade A~C),扩展到6个(Clade A~F)和7个进化枝(Clade A~G)[15−17, 21]。研究表明同一分支具有相似的生物学功能,可以为基因功能分析提供参考[14]。
染色体定位结果表明:RchPGs基因随机不均匀分布在掌叶覆盆子的7条染色体上,且发生了4次串联复制事件。番茄中28个PGs基因也不均匀分布在12条染色体上,且有5个串联重复基因簇[12]。其中,在掌叶覆盆子染色体上存在2个三基因串联重复区域(RchPG8、RchPG9、RchPG10和RchPG30、RchPG31、RchPG32基因簇),分别位于系统发育树的Clade C和Clade D分支上。WANG等[22]在无花果Ficus carica中也发现了2个五基因的串联重复区域,也分别位于Clade C和Clade D分支上。这表明不同物种的PG基因分布模式相似,且存在基因扩张现象。
转录组分析结果表明:Clade E分支中,除RchPG46以外的PG基因在果实发育过程中表达量较高,但表达模式有差异。其中,RchPG6、RchPG29、RchPG1、RchPG12和RchPG23在果实发育早期起作用,参与果实膨大过程中的细胞壁水解;相反地,RchPG2、RchPG28、RchPG38在果实发育后期起作用,介导果实成熟与软化。且共线基因对RchPG29和RchPG47、RchPG12和RchPG23的表达模式相近。这些蛋白中均缺少完整的结构域Ⅲ,属于鼠李型PGs。拟南芥、番茄和甘薯的Clade E分支PGs蛋白成员也缺少结构域Ⅲ[12, 14, 17]。结构域Ⅰ和Ⅱ可能构成催化位点,结构域 Ⅲ可能参与催化反应,结构域Ⅳ可能与底物中羧酸分子的离子基团相互作用[14]。可见,在介导果实发育时PG的结构域Ⅲ可能是非必需的。另一方面,Clade C分支中的RchPG10和Clade B分支中的RchPG16在果实发育后期表达量剧增,为介导果实成熟软化的关键PG基因。它们均具有完整的4个典型结构域。KE等[12]研究发现:SlPG14、SlPG15和SlPG49基因在番茄果实发育后期有较高表达水平,并通过影响细胞壁裂解过程来调控番茄果实成熟软化[12]。共线性分析结果表明:SlPG49和RchPG28具有共线性。系统发育分析结果表明:RchPG16和SlPG14及SlPG15亲缘关系最近,这些结果进一步证实了RchPG28和RchPG16在掌叶覆盆子果实成熟软化过程中的主导作用。据推测,Clade A和Clade B的成员包含内切型PGs,Clade C和Clade D成员包含外切型PGs,Clade E成员包含鼠李型PGs,Clade F成员既不是外切型PGs也不是内切型PGs。由于不同类型的PG酶具有不同的底物和产物,每个分支所催化的果胶组分的差异可能是进化差异导致的[14]。掌叶覆盆子PG家族成员在发育过程中各司其职,协同促进果实的膨大与成熟软化。
掌叶覆盆子的育种目标之一是增大药果果型。果实的膨大受到细胞壁的限制。果实成熟过程中大小和硬度的变化包括纤维素、半纤维素和果胶等物质的溶解和解聚,以及它们之间的重塑。PG可降解果胶,催化果胶分子中α-1,4-聚半乳糖醛酸的裂解,使其降解为寡聚半乳糖醛酸和半乳糖醛酸,进一步被果胶裂解酶(PL)催化裂解[23]。而对于鲜果市场,口感佳是更重要的育种目标,若果实大硬度高,影响口感,因此需选育出花托软的品种。PG家族成员丰富,在果实发育过程中相互协调。樱桃Cerasus pseudocerasus在发育过程中,PG和PL在果实膨大期迅速升高,促进了原果胶的分解,可溶性果胶含量显著增加;而转色期PG活性显著降低[24]。梨Pyrus communis PcPG1和苹果Malus domestica MdPG1在果实成熟过程中表达量逐渐上升[25−26];番茄PG基因的表达量随乙烯的增加而增强;外源乙烯处理可通过增强PG基因的表达而促进番茄软化;在番茄中过表达红树莓Rubus idaeus RiPG2基因可加速果实软化[27−28]。草莓Fragaria × ananassa FaPG2的高表达是启动果实软化的因素之一[29]。曾燕如等[30]通过硼酸处理芒果Mangifera indica果实,发现PG基因表达量下降,延缓果实成熟。陈迪飞等[31]对莲雾Syzygium samarangense喷施外源脱落酸(ABA),发现果实中PG基因活性显著提高,促进果实的成熟与软化。在本研究中,不同覆盆子PG家族成员在果实发育阶段所起的作用不同,介导果实膨大和果实软化的成员不同。因此根据不同的育种目标,要针对性地选择合适的基因做分子标记或基因工程育种。
此外,RchPG3~RchPG5、RchPG7~RchPG9、RchPG13~RchPG15等基因在果实中表达量较低,推测它们可能在根、茎、叶等其他器官发育过程中起作用。相关研究表明:PG基因家族除作用于果实外,还在植物其他器官中呈现高表达水平。陈迪飞等[31]发现莲雾SsPG12、SsPG13和SsPG24在根中特异性表达,SsPG2和SsPG4在根、茎、叶中呈现高表达水平;霍如雪等[32]发现桃prunus persica 中PpaPG60在茎中呈现高表达量,PpaPG80在叶中呈现高表达量。关于掌叶覆盆子PG基因在根、茎、叶等其他器官发育过程的表达与调控有待后续深入研究。
-
本研究从掌叶覆盆子基因组中鉴定到47个RchPGs基因,可分为6个分支(Clade A~F)。RchPGs基因在掌叶覆盆子果实发育不同阶段差异表达,RchPG6和RchPG29等基因在果实发育早期高表达,而RchPG16和RchPG28等基因在果实成熟软化时强烈表达,说明RchPGs不同成员协同调控果实的膨大与软化。
Genome-wide identification of the polygalacturonase gene family in Rubus chingii and its roles in fruit development
-
摘要:
目的 对掌叶覆盆子Rubus chingii果胶水解酶中的多聚半乳糖醛酸酶(polygalacturonase,PG)基因家族进行鉴定与分析,找出果实发育过程中的关键编码基因PGs,可为实现药果果型大或红果花托软的育种目标提供理论参考。 方法 基于掌叶覆盆子基因组,利用生物信息学方法筛选和鉴定掌叶覆盆子PG基因家族成员,并利用转录组测序和实时荧光定量PCR (RT-qPCR)进行表达模式分析。 结果 共鉴定了47个覆盆子RchPGs基因,不均匀地分布在7条染色体上,根据系统发育可分为6个分支(Clade A~D)。其中28个RchPGs蛋白都具有完整的4个结构域Ⅰ (SPNTDG)、Ⅱ (GDDC)、Ⅲ (CGPGHGISIGSLG)和Ⅳ (RIK);11个RchPGs蛋白不包含结构域Ⅲ。转录组测序结果表明:RchPG6、RchPG29、RchPG1、RchPG23和RchPG12等在果实发育早期起作用,参与果实膨大过程中的细胞壁水解;而RchPG16、RchPG2、RchPG10、RchPG28和RchPG38等则在果实发育后期表达量升高,介导果实成熟与软化。尤其是RchPG16,红果期表达量急剧升高,每千个碱基/百万个定位片段(FPKM)是未成熟大绿(BGⅠ)时期的 2168.17 倍,说明RchPG16对果实成熟与软化具有至关重要的作用。结论 RchPG基因家族成员在掌叶覆盆子果实发育不同阶段差异表达,分别促进果实膨大和软化,其中RchPG16和RchPG28在果实成熟软化过程中可能起到关键作用。图8表1参32 Abstract:Objective The polygalacturonase (PG) gene family members of pectin hydrolase in Rubus chingii were identified and analyzed in this study, and the vital PG-coding genes during fruit development were screened, which could provide a theoretical reference for achieving the breeding goals to expand the fruit size or inprove fruit softening. Method Genome-wide identification and bioinformatics methods were used to identify the PG gene family members of R. chingii. Transcriptome sequencing and RT-qPCR were used to analyze expression patterns of PG genes during fruit developmental stages. Result A total of 47 RchPGs were predicted, which were unevenly distributed on 7 chromosomes. Phylogenetic relationships showed they could be divided into 6 clades (Clade A−D). Among these, 28 RchPGs proteins had integrated 4 domains, Ⅰ (SPNTDG), Ⅱ (GDDC), Ⅲ (CGPGHGISIGSLG), and Ⅳ (RIK), while 11 RchPGs missed domain Ⅲ. The result of the transcriptome sequencing showed that RchPG6, RchPG29, RchPG1, RchPG23 and RchPG12 involved in hydrolysis of cell wall for fruit expansion at earlier stages of fruit development. Oppositely, RchPG16, RchPG2, RchPG10 and RchPG38 were highly expressed at later stages and might be essential for fruit ripening and softening. In particular, the expression level (FPKM values from RNA-seq) of RchPG16 was dramatically increased at the red fruit stage, and the FPKM value was 2168.17 times higher than that at the immature big green (BGⅠ) stage, indicating that it played a significant role in fruit ripening and softening.Conclusion RchPG gene family in R. chingii were differentially expressed in different stages of fruit development to improve fruit expansion and softening. Among them, RchPG16 and RchPG28 genes may play a key role in the ripening and softening of R. chingii. [Ch, 8 fig. 1 tab. 32 ref.] -
Key words:
- polygalacturonase /
- Rubus chingii /
- fruit expanding /
- ripening and softening /
- bioinformatics
-
图 3 掌叶覆盆子与拟南芥、番茄PG家族成员的系统发育树
Figure 3 Phylogenetic tree of PG family members from R. chingii, A. thaliana, and S. lycopersicum
图 4 掌叶覆盆子RchPGs基因家族结构分析以及系统发育树
Figure 4 Gene structures analyses of RchPGs families and phylogenetic tree of RchPGs in R. chingii
图 5 掌叶覆盆子RchPGs 蛋白家族保守基序预测
Figure 5 Motif prediction of RchPGs family members in R. chingii
图 6 掌叶覆盆子RchPGs 蛋白家族保守基序序列分析
Figure 6 Motif sequence analysis of RchPGs family members in R. chingii
图 7 RchPGs基因染色体定位和共线性分析
Figure 7 Chromosomal location and synteny analyses of RchPGs in R. chingii
图 8 掌叶覆盆子RchPGs基因表达量转录组分析与RT-qPCR鉴定
Figure 8 Transcriptome analysis and RT-qPCR identification of RchPGs genes expression in R.chingii
表 1 掌叶覆盆子RchPGs家族成员特征
Table 1. Characteristics of RchPGs family members in R. chingii
基因编码 基因名称 等电点 分子量/kDa 亲疏水
指数不稳定
系数分支 基因编码 基因名称 等电点 分子量/kDa 亲疏水
指数不稳定
系数分支 LG01.1059 RchPG1 5.96 51.08 −0.073 44.79 E LG04.283 RchPG25 8.63 57.49 −0.356 45.27 A LG01.1356 RchPG2 6.83 51.99 −0.127 44.12 E LG04.396 RchPG26 5.26 49.55 −0.183 33.24 B LG01.1565 RchPG3 8.11 42.23 0.056 29.10 C LG04.993 RchPG27 9.02 52.04 −0.278 44.66 A LG01.1665 RchPG4 6.43 39.40 −0.063 24.24 C LG05.1140 RchPG28 8.48 44.96 −0.174 42.43 E LG01.2532 RchPG5 6.08 47.67 −0.267 38.63 D LG05.1261 RchPG29 5.19 52.20 −0.018 38.83 E LG01.2853 RchPG6 9.09 55.47 −0.193 43.86 E LG06.1099 RchPG30 8.90 50.20 −0.242 28.08 D LG01.3312 RchPG7 8.23 58.83 −0.239 46.37 F LG06.1103 RchPG31 6.96 43.60 −0.148 23.68 D LG01.3577 RchPG8 8.52 15.81 0.008 38.87 C LG06.1105 RchPG32 8.98 42.48 −0.210 27.47 D LG01.3578 RchPG9 9.49 41.00 0.035 39.66 C LG06.1675 RchPG33 7.81 43.81 −0.197 35.69 F LG01.3579 RchPG10 9.48 107.01 −0.032 41.27 C LG06.2235 RchPG34 8.81 49.23 −0.059 43.94 B LG02.1098 RchPG11 8.01 40.06 −0.059 30.89 F LG06.272 RchPG35 7.48 53.19 −0.275 36.87 A LG02.1344 RchPG12 5.39 49.49 −0.063 43.95 E LG06.3272 RchPG36 5.50 51.64 −0.186 36.12 B LG02.1359 RchPG13 5.76 50.91 −0.192 39.89 D LG06.3274 RchPG37 5.00 45.35 −0.167 34.66 F LG02.1479 RchPG14 6.24 131.12 −0.585 51.01 D LG06.3368 RchPG38 5.65 52.96 −0.054 44.42 E LG02.1829 RchPG15 9.35 49.19 −0.451 34.18 B LG06.4848 RchPG39 7.16 48.48 −0.108 48.34 A LG02.1835 RchPG16 8.94 49.80 −0.288 30.39 B LG07.1450 RchPG40 8.95 48.86 −0.283 34.24 B LG02.2252 RchPG17 7.48 49.52 −0.254 38.72 D LG07.1820 RchPG41 7.94 40.24 −0.292 32.80 B LG02.4536 RchPG18 6.38 44.22 −0.015 31.62 D LG07.1821 RchPG42 6.30 48.45 −0.227 25.48 B LG03.1665 RchPG19 8.94 44.71 −0.181 27.06 D LG07.1856 RchPG43 6.44 48.39 −0.256 29.74 B LG03.5002 RchPG20 5.95 52.65 −0.228 40.49 D LG07.1859 RchPG44 7.80 48.12 −0.263 23.46 B LG04.1300 RchPG21 6.39 40.22 −0.357 27.43 F LG07.2143 RchPG45 5.19 45.73 −0.275 37.35 F LG04.1461 RchPG22 6.14 74.72 −0.390 57.75 A LG07.325 RchPG46 7.54 51.44 −0.076 37.22 E LG04.1621 RchPG23 6.14 46.91 0.094 48.01 E LG07.3923 RchPG47 6.09 49.37 −0.170 34.22 E LG04.1640 RchPG24 8.74 46.54 −0.105 33.44 D  下载: 导出CSV
下载: 导出CSV
-
[1] CHEN Zhen, JIANG Jingyong, LI Xiaobai, et al. Bioactive compounds and fruit quality of Chinese raspberry, Rubus chingii Hu varied with genotype and phenological phase[J/OL]. Scientia Horticulturae, 2021, 281 : 109951[2024-05-26]. DOI: 10.1016/j.scienta.2021.109951. [2] MENG Qinglin, MANGHWAR H, HU Weiming. Study on Supergenus Rubus L.: edible, medicinal, and phylogenetic characterization[J/OL]. Plants, 2022, 11 (9): 1211[2024-05-26]. DOI: 10.3390/plants11091211. [3] SHENG Jiayun, WANG Siqi, LIU Kaohua, et al. Rubus chingii Hu: an overview of botany, traditional uses, phytochemistry, and pharmacology[J]. Chinese Journal of Natural Medicines, 2020, 18 (6): 401−416. [4] 国家药典委员会. 中华人民共和国药典[M]. 北京: 中国医药科技出版社, 2020: 399. Chinese Pharmacopoeia Commission. Pharmacopoeia of the People’s Republic of China[M]. Beijing: China Medical Science Press, 2020: 399. [5] YU Guohua, LUO Zhiqiang, WANG Wubin, et al. Rubus chingii Hu: a review of the phytochemistry and pharmacology [J/OL]. Frontiers in Pharmacology, 2019, 10 : 799[2024-05-26]. DOI: 10.3389/fphar.2019.00799. [6] CHEN Zhen, JIANG Jingyong, SHU Liangzuo, et al. Combined transcriptomic and metabolic analyses reveal potential mechanism for fruit development and quality control of Chinese raspberry (Rubus chingii Hu)[J]. Plant Cell Reports, 2021, 40: 1923−1946. [7] HUA Yujiao, DAI Bingyi, LUO Yiyuan, et al. Integrated analysis of multiple metabolome and transcriptome revealed the accumulation of flavonoids and associated molecular regulation mechanisms in Rubus chingii Hu at different developmental stages[J/OL]. Plant Physiology and Biochemistry, 2023, 204 : 108085[2024-05-26]. DOI: 10.1016/j.plaphy.2023.108085. [8] WU Mengjun, LYU Yuxin, XU Hangying, et al. Raspberry polysaccharides attenuate hepatic inflammation and oxidative stress in diet-induced obese mice by enhancing butyrate-mediated intestinal barrier function[J/OL]. International Journal of Biological Macromolecules, 2024, 262 : 130007[2024-05-26]. DOI: 10.1016/j.ijbiomac.2024.130007. [9] ADAMCZUK N, OŚKO J, GREMBECKA M, et al. Evaluation of the content of micro- and macroelements in raspberries depending on the species, cultivar variety, and geographical environment[J/OL]. Nutrients, 2023, 15 (17): 3782[2024-05-26]. DOI: 10.3390/nu15173782. [10] GONZÁLEZ-CARRANZA Z H, ELLIOTT K A, ROBERTS J A. Expression of polygalacturonases and evidence to support their role during cell separation processes in Arabidopsis thaliana[J]. Journal of Experimental Botany, 2007, 58(13): 3719−3730. [11] YU Youjian, LIANG Ying, LÜ Meiling, et al. Genome-wide identification and characterization of polygalacturonase genes in Cucumis sativus and Citrullus lanatus [J]. [J]. Plant Physiology and Biochemistry, 2014, 74: 263−275. [12] KE Xubo, WANG Huasen, LI Yang, et al. Genome-wide identification and analysis of polygalacturonase genes in Solanum lycopersicum[J/OL]. International Journal of Molecular Sciences, 2018, 19 (8): 2290[2024-05-26]. DOI: 10.3390/ijms19082290. [13] LU Lu, HOU Quancan, WANG Linlin, et al. Genome-wide identification and characterization of polygalacturonase gene family in maize (Zea mays L. )[J/OL]. International Journal of Molecular Sciences, 2021, 22(19): 10722[2024-05-26]. DOI: 10.3390/ijms221910722. [14] HE Peiwen, ZHANG Jingzhen, LÜ Zunfu, et al. Genome-wide identification and expression analysis of the polygalacturonase gene family in sweetpotato[J/OL]. BMC Plant Biology, 2023, 23 : 300[2024-05-26]. DOI: 10.1186/s12870-023-04272-1. [15] PARK K C, KWON S J, KIM P H, et al. Gene structure dynamics and divergence of the polygalacturonase gene family of plants and fungus[J]. Genome, 2008, 51(1): 30−40. [16] LIANG Ying, YU Youjian, CUI Jinlong, et al. A comparative analysis of the evolution, expression, and cis-regulatory element of polygalacturonase genes in grasses and dicots[J]. Functional & Integrative Genomics, 2016, 16(6): 641−656. [17] KIM J, SHIU S H, THOMA S, et al. Patterns of expansion and expression divergence in the plant polygalacturonase gene family[J/OL]. Genome Biology, 2006, 7 (9): R87[2024-05-26]. DOI: 10.1186/gb-2006-7-9-r87. [18] MAHMOOD U, FAN Yonghai, WEI Siyu, et al. Comprehensive analysis of polygalacturonase genes offers new insights into their origin and functional evolution in land plants[J]. Genomics, 2021, 113(1): 1096−1108. [19] WANG Longji, LEI Ting, HAN Guomin, et al. The chromosome-scale reference genome of Rubus chingii Hu provides insight into the biosynthetic pathway of hydrolyzable tannins[J]. The Plant Journal, 2021, 107: 1466−1477. [20] CHEN Chengjie, CHEN Hao, ZHANG Yi, et al. TBtools: an integrative toolkit developed for interactive analyses of big biological data[J]. Molecular Plant, 2020, 13: 1194−1202. [21] QIAN Ming, ZHANG Yike, YAN Xiangyan, et al. Identification and expression analysis of polygalacturonase family members during peach fruit softening[J/OL]. International Journal of Molecular Sciences, 2016, 17 (11): 1933[2024-05-26]. DOI: 10.3390/ijms17111933. [22] WANG Yuan, FAN Zhiyi, ZHAI Yanlei, et al. Polygalacturonase gene family analysis identifies FcPG12 as a key player in fig (Ficus carica L. ) fruit softening[J/OL]. BMC Plant Biology, 2023, 23 (1): 320[2024-05-26]. DOI: 10.1186/s12870-023-04315-7. [23] MARKOVIC O, JANECEK S. Pectin degrading glycoside hydrolases of family 28: sequence-structural features, specificities and evolution[J]. Protein Engineering, 2001, 14(9): 615−631. [24] 齐勇, 韩秀梅, 宋莎, 等. “落别”樱桃果实不同发育期糖酸组分积累及软化相关成分研究[J]. 食品工业科技, 2021, 42(21): 344−352. QI Yong, HAN Xiumei, SONG Sha, et al. Study on accumulation of sugar and acid components and softening related components of “Luobie” cherry fruit in the different development stages[J]. Science and Technology of Food Industry, 2021, 42(21): 344−352. [25] SEKINE D, MUNEMURA I, GAO Mei, et al. Cloning of cDNAs encoding cell-wall hydrolases from pear (Pyrus communis) fruit and their involvement in fruit softening and development of melting texture[J]. Physiologia Plantarum, 2006, 126(2): 163−174. [26] ATKINSON R G, SCHRÖDER R, HALLETT I C, et al. Overexpression of polygalacturonase in transgenic apple trees leads to a range of novel phenotypes involving changes in cell adhesion[J]. Plant Physiology, 2002, 129(1): 122−133. [27] SITRIT Y, BENNETT A B. Regulation of tomato fruit polygalacturonase mRNA accumulation by ethylene: a re-examination[J]. Plant Physiology, 1998, 116(3): 1145−1150. [28] LI Tiemei, GUO Xiao, CHEN Yuxiao, et al. Overexpression of the Rubus idaeus polygalacturonases gene RiPG2 accelerates fruit softening in Solanum lycopersicum[J/OL]. Agronomy, 2024, 14 (1): 160[2024-05-26]. DOI: 10.3390/agronomy14010160. [29] REN Yibo, LI Baijun, JIA Haoran, et al. Comparative analysis of fruit firmness and genes associated with cell wall metabolisms in three cultivated strawberries during ripening and postharvest[J/OL]. Food Quality and Safety, 2023, 7 : fyad020[2024-05-26]. DOI: 10.1093/fqsafe/fyad020. [30] 曾燕如, SRIVASTAVA G C, PANDEY M, 等. 硼酸处理对Sensatiou芒果成熟的影响[J]. 浙江林学院学报, 1996, 13(3): 263−269. ZENG Yanru, SRIVASTAVA G C, PANDEY M, et al. Effect of borie acid treatment on the ripening of mango (Mangifera indica) cv. Sensation[J]. Journal of Zhejiang Forestry College, 1996, 13(3): 263−269. [31] 陈迪飞, 魏秀清, 许玲, 等. 莲雾PG基因家族全基因组鉴定及表达分析[J]. 果树学报, 2022, 39(4): 548−563. CHEN Difei, WEI Xiuqing, XU Ling, et al. Genome-wide identification and expression analysis of PG gene family in wax apple [Syzygium samarangense (Bl. ) Merr. et Perry][J]. Journal of Fruit Science, 2022, 39(4): 548−563. [32] 霍如雪, 刘振宁, 杨青, 等. 桃PG基因家族的鉴定与分析[J]. 江苏农业科学, 2016, 44(6): 33−40. HUO Ruxue, LIU Zhenning, YANG Qing, et al. Identification and analysis of PG gene family in peach[J]. Jiangsu Agricultural Sciences, 2016, 44(6): 33−40. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20240427
 点击查看大图
点击查看大图

计量
- 文章访问数: 2029
- HTML全文浏览量: 641
- PDF下载量: 57
- 被引次数: 0
